
Abstract
Forests can prevent and/or mitigate hydrogeomorphic hazards in mountainous landscapes. Their effect is particularly relevant in the case of shallow landslides phenomena, where plants decrease the water content of the soil and increase its mechanical strength. Although such an effect is well known, its quantification is a relatively new challenge. The present work estimates the effect of some forest species on hillslope stability in terms of additional root cohesion by means of a model based on the classical Wu and Waldron approach (Wu in Alaska Geotech Rpt No 5 Dpt Civ Eng Ohio State Univ Columbus, USA, 1976; Waldron in Soil Sci Soc Am J 41:843–849, 1977). The model is able to account for root distribution with depth and non-simultaneous root breaking. Samples of European beech (Fagus sylvatica L.), Norway spruce (Picea abies (L.) Karst.), European larch (Larix decidua Mill.), sweet chestnut (Castanea sativa Mill.) and European hop-hornbeam (Ostrya carpinifolia Scop.), were taken from different locations of Lombardy (Northern Italy) to estimate root tensile strength, the Root Area Ratio and the root cohesion distribution in the soil. The results show that, in spite of its dramatic variability within the same species at the same location and among different locations, root cohesion can be coherently interpreted using the proposed method. The values herein obtained are significant for slope stabilisation, are consistent with the results of direct shear tests and back-analysis data, and can be used for the estimation of the stability of forested hillslopes in the Alps.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forests can play a significant role in preventing and/or mitigating hydrogeomorphic hazards, such as floods, shallow landslides, debris flows, debris avalanches, rockfalls, and snow avalanches (see Sidle and Ochiai 2006). The beneficial effects of forests in mountainous landscape are well-known. Rules demonstrating recognition of the protective function of forests by limiting forest-clearing activities can be found in documents and regulations of the Republic of Venice from as early as the 13th and 14th centuries. Since the end of the 19th century, preserving the protective effect of forests has been key strategy of the governments of European Alpine countries to defend mountain territory from disasters. Such policies represent a systematic combination of an engineering approach with a biological approach, originating a new discipline referred to as torrent control and hillslope stability (called sistemazioni idraulico-forestali, in Italy, Wildbach und Lawinenverbauung, in Austria and Germany, and Restauration de Terrains en Montagne, in France). The same principle underlies those techniques now referred to soil bioengineering.
The protective effect of forests was not studied from a scientific perspective until the second half of the last century, based on the consequences of forest-clearing operations (see Sidle and Ochiai 2006). Some decades later, such studies were confirmed by more specific experiments concerning the shear resistance of rooted soils, in the field (Endo and Tsuruta 1969; Wu et al. 1979; Wu et al. 1988) and the laboratory (e.g. Waldron 1977; Waldron and Dakessian 1981).
Vegetation affects the stability of slopes, influencing both hydrological processes (which affect the water content in the soil and then the pore pressure) and the mechanical structure of the soil (which affects its strength). The magnitude of such effects depends on root system development, which, in turn, is a function of genetic properties of the plant species and of environmental characteristics (soil texture and structure, aeration, moisture, temperature, competition with other plants, etc.). The environmental characteristics, in particular, induce a great spatial variability of root patterns, introducing a dramatic heterogeneity in soil reinforcement across different depths, planes and locations.
Limiting our attention to the mechanical effects of the root system, two main actions are recognised. The first of these involves small flexible roots that mobilise their tensile strength by soil-root friction, increasing the compound matrix (soil-fibre) strength. The second involves large roots intersecting the shear surface, which, acting as individual anchors that eventually slip through the soil matrix without breaking, mobilise a soil-root friction force instead of the entire tensile strength (Waldron 1977). Both effects can be quantified via modelling (see Gray and Laiser 1982; Morgan and Rickson 1995; Greenwood 2006) if appropriate parameters are provided. Only the fibre reinforcement mechanism, however, is generally considered. The latter is expressed in terms of additional root cohesion, which can be easily incorporated into slope stability models (Schmidt et al. 2001; Roering et al. 2003; Greenwood 2006; Sidle and Ochiai 2006).
After the pioneering works of Schiechtl (1958) and Endo and Tsuruta (1969), researchers have begun to systematically investigate this area, and the number of studies on soil reinforcement by roots has increased ever since (Schmidt et al. 2001; Roering et al. 2003; Sakalas and Sidle 2004; Bischetti et al. 2005; Norris 2005; van Beek et al. 2005; Tosi 2007; De Baets et al. 2008; Normaniza et al. 2008; Norris et al. 2008a; Sun et al. 2008; limiting to works dealing with hillslope stability and the last few years). Most of the work in this field has been performed in North America, Asia and Oceania (Nilaweera and Nutalaya 1999; Schmidt et al. 2001; Roering et al. 2003; Normaniza et al. 2008) and, although several researches have been recently carried out in Europe (Bischetti et al. 2005; Norris 2005; van Beek et al. 2005; Tosi 2007; De Baets et al. 2008), there is still a lack of knowledge for the European Alpine and Prealpine species.
In order to fill this gap and provide quantitative information regarding natural hazard prevention and mitigation, we investigated the role of five Alpine and Prealpine forest species in stabilizing hillslopes, in terms of additional root cohesion. A method based on a general interpretation of the scheme of Wu and Waldron (Wu 1976; Waldron 1977) was developed to specifically account for the distribution of root cohesion within the soil and non-simultaneous root breaking. Tensile strength and root density data, which are the necessary inputs for this method, were collected for more than one profile at some locations of the Alps and Prealps of the Lombardy Region and the results are analysed to estimate root cohesion with depth.
Root cohesion evaluation
It is widely recognised that fibre reinforcement of rooted soil depends on the strengths of roots and their density and distribution in the soil (Wu 1976; Waldron 1977 and Ziemer 1981 among the first). The evaluation of such reinforcement, in terms of root cohesion, can be obtained by means of direct shear tests (in situ or in laboratory) and by means of back analysis of collapsed forested hillslopes (see Wu 1995). Such methods, however, can only provide data for the upper soil layer, in the first case, and averaged values for the entire soil profile, in the second case. Moreover, due to site-specific development of root systems, which leads to a dramatic space variability of root density and size, the results are valid only for the specific (or highly similar) conditions that occur in the location where the investigations are carried out.
A more general way of estimating root reinforcement is to model root behaviour along the soil profile during shearing. The most common scheme adopted to estimate root cohesion in this way is the Wu (1976) and Waldron (1977) model (W&W Model), despite its simplicity (Sidle and Ochiai 2006). New and more complex models have recently been proposed (e.g. Ekanayake and Phillips 1999; Frydman and Operstein 2001; Pollen and Simon 2005), but the W&W model still represents the benchmark.
The W&W model assumes that roots are cylindrical and elastic, and that they are extending perpendicular to the shear surface. When the rooted soil is sheared, the embedded roots bend and mobilise their tensile strength by means of root-soil friction. The mobilised tensile force is then resolved into a tangential component and a normal component. The tangential component opposes the shear force and the normal component increases the confining pressure on the shear surface and the soil resistance, assuming the Mohr–Coulomb equation as shear criterion. The fibre reinforcement in terms of root cohesion (c r) can then be written as:
where φ is the soil friction angle, θ is the angle of root deformation from the vertical, t r is the mean root tensile strength mobilised per unit area of soil.
tr can be estimated as \(T_{\text{r}} \;\left( {{{A_{\text{r}} } \mathord{\left/ {\vphantom {{A_{\text{r}} } A}} \right. \kern-\nulldelimiterspace} A}} \right)\), where Tr is the mean tensile strength of the roots and (Ar/A) is the ratio between the cross sectional area of the roots crossing a plane within the soil and the plane area (the so called Root Area Ratio, RAR).
It can be shown that for 40° < θ < 90° and 25° < φ < 40°, which are generally considered reliable values for most real cases (Wu et al. 1979), the term in the brackets of Eq. 1 has a limited range between 1.0 and 1.3, and, thus, an average value can be set. Few studies have explored this point, though a recent paper by Docker and Hubble (2008) reports, for riparian vegetation of New Zealand, values less than 1 (around 0.75). Based on this finding, and due to the multiplicative form of the W&W model, the common use of the standard average value of 1.15 (Waldron 1977) or 1.2 (Wu et al. 1979), might lead to a significant overestimation of root cohesion.
In any case, Eq.1 can be written in a general form as follows:
where k′ is the factor accounting for the decomposition of root tensile strength according to the bending angle of roots with respect to the shear plane.
Tr is affected by species and differences in diameter. The tensile strength-diameter relationship is generally accepted to follow a power law form (Burroughs and Thomas 1977; Abe and Iwamoto 1986; Gray and Sotir 1996; Nilaweera and Nutalaya 1999; Bischetti et al. 2005; Genet et al. 2005):
where a and b are species-dependent parameters (Bischetti et al. 2005), although some researchers have observed intra-species differences (Hathaway and Penny 1975; Genet et al. 2005 and 2006). Genet et al. (2005, 2006), in particular, showed that tensile resistance is dependent on cellulose content, which can vary with the local growth conditions.
To account for the variability of root diameter, Eq. 2 must be rewritten as follows:
where T r is the tensile strength and a r is the RAR, both specified per diameter class i, and N is the number of classes considered.
The original W&W model assumes that all the roots crossing the shear surface break at the same time. In real cases, it is expected that roots will break at different times, based on their size, growth direction and tortuosity. Such behaviour, typical of composite materials constituted by fibres embedded in a matrix, can be easily observed on qualitative basis. For example, when pulling out a plant, several consecutive snaps can be heard, which are related to the progressive breaking of roots. This phenomenon has been demonstrated by means of pullout experiments on branched roots (Riestenberg 1994; Norris 2005; Docker and Hubble 2008) and direct shear tests (Docker and Hubble 2008).
As a consequence, the application of the original W&W model tends to overestimate root reinforcement, as observed by several authors (Waldron and Dakessian 1981; Operstein and Frydman 2000; Pollen and Simon 2005; Docker and Hubble 2008) and the values obtained should be viewed as the maximum potential reinforcement.
To account for non-simultaneous root breaking, an adjustment factor can be introduced and Eq. 1 can be rewritten as follows:
where k″ is a factor accounting for the non-simultaneous breaking of roots.
Few references are available that discuss the reduction factor k″; Hammond et al. (1992) proposed, for forest vegetation, a reduction factor of 0.56, whereas Waldron and Dakessian (1981), Operstein and Frydman (2000), Pollen and Simon (2005) and Docker and Hubble (2008) observed lower values for herbaceous plants and very young trees. Pollen and Simon (2005) carried out experiments using the Fiber Bundle Model (FBM) approach on riparian tree vegetation and obtained reduction factors between 0.60 and 0.82. The FBM approach is a well-known scheme introduced several decades ago to evaluate the strength of fibrous materials (Daniels 1945). In the last few years, this approach has been widely adopted to study the statics and dynamics of failures in materials under stress (Hemmer et al. 2007; Kun et al. 2007 and Raischel et al. 2008). As such, the FBM approach represents a promising perspective for evaluating the reduction factor k″; however, it needs to be tested further because some of the underlying hypotheses have not been fully verified for rooted soil (e.g., all roots have the same elasticity).
Several studies have been conducted regarding the distribution of roots in forested soil, although most of these focus on forest ecology and are not necessarily applicable to evaluation of root cohesion. In those studies where root density and RAR were determined, it is shown that such quantities decrease with depth and distance from the stem. (e.g. Abernethy and Rutherfurd 2001; Danjon et al. 2008).
Material and methods
Study sites and species
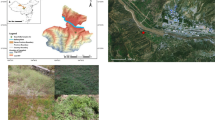
The data presented here were collected in six locations of the Lombardy Alps and Prealps (Fig. 1 and Table 1). Morterone (M) is located in the Taleggio valley (a right-hand flank tributary of Val Brembana - Bergamo); we excavated four trenches in a European beech (Fagus sylvatica L.) forest at 1,100 m a.s.l. In this location, the soil consists of silt and clayey sand and fine gravel and the average annual precipitation is about 1,800 mm. Alpe Gigiai (AG) is located on the north-western side of Como Lake (Sirco Valley); we excavated three trenches in a coppiced beech forest at about 1,400 m a.s.l., three trenches in a Norway spruce (Picea abies (L.) Karst.) forest at about 1,500–1,600 m a.s.l., and four trenches in a European larch (Larix decidua Mill.) forest at about 1,600 m a.s.l.. In this location, the soil is a gravel-sand mixture with silty matrix and the average annual precipitation is about 1,750 mm. Monte Pora (MP) is located in Northern Val Seriana (Bergamo); we excavated ten trenches in a Norway spruce forest at 1,400–1,500 m a.s.l., two trenches in a European larch forest at 1,500 m a.s.l. and one trench in a beech forest at 1,400 m a.s.l.. In this location, the soil is clayey and the average annual precipitation is about 1,500 mm. Alpe Bess (AB) and Alpe Giumello (AGm) are located on the north-eastern side of Como Lake (Higher Valsassina); we excavated five trenches in sweet chestnut (Castanea sativa Mill.) forests both at 1,000 m a.s.l.. In this location, the soil is a gravel-sand mixture and the average annual precipitation is about 1,550 mm. Pasturo (P) is located on the south-eastern side of Como Lake (Valsassina); we excavated four trenches in a European hop-hornbeam (Ostrya carpinifolia Scop.) forest at 750 m a.s.l. In this location, the soil is gravel-sand mixture and the average annual precipitation is about 1,600 mm.

Location of experimental sites
Site and forest characteristics are summarised in Table 1.
Tensile strength tests
Live roots used for tensile strength tests were collected by digging pits or trenches, taking care to avoid any root damage or stress. Samples were then put in separate bags, sealed and transported to the laboratory for tensile strength testing. In most of cases, tensile tests were carried out on fresh roots within 1 week after sampling; in other cases, we preserved the roots for a few weeks using a 15% alcohol solution (Meyer and Gottsche 1971), which has no influence on the measured parameters (Bischetti et al. 2003). Tests were carried out on roots with typical tortuousness, ranging in size from threads to diameters between 0.12 mm to about 7 mm.
Testing was performed with a device designed and built by the Institute of Agricultural Hydraulics, consisting of a strain apparatus controlled by an electrical motor. Roots were attached to the specifically developed clamping devices that avoid root damage at the clamping points, and tensile force was exerted by a system of gears at a rate of 10 mm/min. Tensile strength was recorded by a load cell (F.S. = 500 N, accuracy = 0.1% F.S.) connected to an acquisition system. Only specimens that broke near the middle were evaluated (since ruptures near the clamps may have been induced by root structure damage instead of tension). Tensile strength at rupture (Pa) was calculated by dividing the peak load (N) by the cross-sectional area of the root (m2), estimated as the average of root diameters measured with bark before traction. A more detailed discussion can be found in Bischetti et al. (2003).
Statistical analysis of the data was performed after log transformation of the strength and diameter values. A Kolmogorov–Smirnov test (ks-test) was used to test the normality of the data at a 1% level of significance before proceeding with analyses of variance (Yazici and Yolacan 2007; Genet et al. 2005).
To evaluate the differences between tensile strength properties of each species, analysis of covariance (ANCOVA) was used, taking into account the diameter as covariate factor. ANCOVA was also applied to evaluate differences between sites of origin of each sample for each species and also between species.
RAR evaluation
RAR data were obtained through the root-wall technique (Burke and Raynal 1994; Schmid and Kazda 2001 & 2002; Vinceti et al. 1998; Xu et al. 1997) by applying image analysis (Vogt and Persson 1991). The more common core-break sampling method (Babu et al. 2001; Burke and Raynal 1994; Büttner and Leuschner 1994; Hendriks and Bianchi 1995; Schmid and Kazda 2002; Xu et al. 1997) was discarded because RAR estimation from root biomass, root number or root length, implies hypotheses about the 3D distribution of roots inside the sample (Lopez-Zamora et al. 2002).
At sampling sites, cut-slopes of forest roads under construction in undisturbed stands were chosen to facilitate the excavation of trenches and expose undisturbed profiles of rooted soil down to the bedrock, as in some cases, trenches in undisturbed slopes proved difficult to excavate (Fig. 2). For each profile, a frame of known size was applied and several images were taken; images were then rectified to correct geometrical deformation and roots were manually digitised.

Example of site and rooted profile (sweet chestnut at AB)
RAR values were obtained at depth increments of 10 cm, counting all roots with a diameter between 1 mm and 10 mm. Roots smaller that 1 mm involve great uncertainty in identification and mapping, both in the field and by visual analysis, so they were excluded. Large roots, in contrast, may strongly affect RAR values without acting in accordance of the W&W reinforcement model, due to their stiffness. Schmidt et al. (2001), observed a greater average diameter in unbroken roots compared to the average diameter of broken roots, although their data do not show a clear size threshold.
Each trench at the same site was assumed to be part of a normally distributed sample. Thus, all the trenches were grouped according to the station, and mean values for each 10 cm layer were evaluated. RAR distributions were tested for normality by the ks-test and the correlation between RAR and depth was considered.
Also in this case, ANCOVA was used with RAR data considered as outcome variable respect to depth classes (covariate),
Root cohesion evaluation
In order to evaluate the stability of a hillslope, the root cohesion at the potential shear surface depth, \(c_{\text{r}}^Z \), must be known (depth-averaged values are not sufficient).
In the case of small volume landslides in shallow forested soils, the sliding mass must exceed both the resistance due to those roots crossing the basal shear surface and the resistance due to those roots intersecting the vertical plane at the detachment scarp (Riestenberg and Sovonick-Dunford 1983; Terwillinger and Waldron 1991; Schmidt et al. 2001; Keim and Skaugset 2003; Roering et al. 2003). In these cases, the total root cohesion, \(c_{\text{r}}^Z \), can be defined by the sum of the basal cohesion at depth Z, \(c_{{\text{bas}}}^Z \), and the lateral cohesion, \(c_{{\text{lat}}}^Z \), respectively. These are expressed as follows:
where N is the number of roots at the given depth, M is the number of depth classes of thickness Δz j .
The use of basal, lateral or the sum of the two root cohesions depends on the specific situation and on the sliding model adopted (infinite slope, slices, etc.). Eqs. 6a and b allow for estimation of the appropriate root cohesion value to be used in the different cases.
In the preset work, the additional cohesion at depth Z due to presence of roots was taken as the sum of basal and lateral root cohesions estimated by means of Eqs. 6a and 6b with some further consideration.
First, due to the uncertainty in the value of the root distortion angle, a cautionary value of 1.00 was assumed in Eq. 2, according to Waldron and Dakessian (1981).
Second, the behaviour of roots that break progressively, mobilising only a portion of the total tensile resistance with each break, can be described well using the FBM approach, although some of the hypotheses involved may not hold in all cases.
The original FBM approach considers a bundle of parallel fibres loaded parallel to the fibres direction, characterised by a statistically distributed strength. Fibres fail when the applied load exceeds a threshold value, and as consequence of failure, the load carried by the broken fibres is redistributed among the remaining intact fibres. This load redistribution consists of transferring stress from the broken to the unbroken fibres, inducing secondary failures that, in turn, induce tertiary ruptures, and so on. The failure avalanche is terminated when the unbroken fibres are able to withstand the entire load or when the material collapses.
One of the critical issues of the FBM approach is the criterion adopted for load redistribution, which can follow two different ways: the “democratic” distribution principle for all intact fibres (Equal Load-Sharing—ELS or Global Load Sharing—GLS) which implies an infinite range of interaction and then neglects stress enhancement in the vicinity of failed regions, and the Local Load-Sharing (LLS), where the load of the failed fibres is shared equally by all intact neighbours.
A second critical point is the time dependence of fibre strength. Fibres modelled by the FBM can be divided into two classes: “static” bundles, containing fibres whose strengths are independent of time, and “dynamic” bundles, where fibres are assumed to have time-dependent elements that capture creep rupture and fatigue behaviours.
In our case, the reduction factor k′′ that accounts for the behaviour of roots, was estimated by application of the static fibre bundle approach under equal load sharing as the ratio between root cohesion estimated by FBM approach and the original W&W model. The adopted approach has the same level of simplicity as the W&W model and can be easily integrated.
Results
Root tensile strength
As shown in Table 2, the root tensile strength for C. sativa and O. carpinifolia strongly decreases with root diameter according to a power law (Fig. 3) as in the case of the other species considered (Bischetti et al. 2005).

Tensile strength-diameter data and power law relationships for Castanea sativa Mill. and O. carpinifolia Scop
Figure 4, showing the power law curves of Eq. 3 for the considered species, suggests that there is a certain degree of similarity of root tensile strength between European hop-hornbeam and sweet chestnut and between Norway spruce and European larch. The two pairs of species are different between each other as well as in respect to European beech. The corresponding regression parameters are summarised in Table 2.

Tensile strength-diameter relationships for the considered species
The results of ANCOVA confirm such an idea. When all the species were considered, the curves resulted significantly different with regards to root diameter (F 4, 493 = 43.246, p < 0.001, ANCOVA). Considering European hop-hornbeam and sweet chestnut on one hand and Norway spruce and European larch on the other, the associated strength-diameter relationships were not statistically different at a 1% level of significance (respectively F 1,126 = 5.21, p = 0.024, ANCOVA and F 1,132 = 1.98, p = 0.16, ANCOVA).
For the species sampled at different sites (P. abies, C. sativa and F. sylvatica), differences in root tensile strength between stations were analysed, and in each of the cases they did not show statistically significant difference (p > 0.01).
Root Area Ratio (RAR)
The RAR data show that the variability for the same species at the same location and depth is very high, although the general trend is a decrease in RAR with depth, with the exception of the first two or three layers, where it generally increases (Fig. 5, 6, 7, 8, 9).

RAR values of Norway spruce (Picea abies (L.) Karst.) as function of depth for the surveyed trenches at the considered sites (Alpe Gigiai AG and M.te Pora MP)

RAR values of European larch (Larix decidua Mill.) as function of depth for the surveyed trenches at the considered sites (Alpe Gigiai AG and M.te Pora MP)

RAR values of European beech (Fagus sylvatica L.) as function of depth for the surveyed trenches at the considered sites (Alpe Gigiai AG, Morterone M and M.te Pora MP)

RAR values of sweet chestnut (Castanea sativa Mill.) as function of depth for the surveyed trenches at the considered sites (Alpe Bess AB and Malpe Giumelo AGM)

RAR values of European hop-hornbeam (Ostrya carpinifolia Scop.) as function of depth for the surveyed trenches at the considered site (Pasturo P)
The maximum rooting depths varied as follows: for Norway spruce, between 40 and 120 cm at MP (average 74 cm) and between 60 and 90 cm at AG (average 70 cm); for European larch, between 80 and 110 cm at MP (average 95 cm) and between 60 and 110 cm at AG (average 85 cm); for European beech, 90 cm at MP, between 60 and 70 cm at AG (average 63 cm) and between 100 and 110 cm at M (average 105 cm); for sweet chestnut, between 100 and 120 cm at AGM (average 104 cm) and between 60 and 90 cm at AB (average 90 cm); for European hop-hornbeam, between 70 and 120 cm at P (average 93 cm).
An analysis of the site-averaged RAR distribution with depth (Fig. 10) showed that for European larch the difference between the two locations is clear, whereas for sweet chestnut it is clear that there is no difference. In the case of Norway spruce and European beech, on the contrary, it is not apparent if the RAR distributions are different or not. ANCOVA has then be applied to verify the visual analysis.

Site-averaged RAR distributions for a P. abies (L.) Karst., b L. decidua Mill., c F. sylvatica L., d C. sativa Mill., e O. carpinifolia Scop
The preliminary analyses of parallelism requested for the application of ANCOVA showed that such condition is verified only in the cases of European larch and sweet chestnut and ANCOVA results confirm that for European larch RAR distributions between locations are statistically different at 1% level of significance (F 1,21 = 7.60, p = 0.01, ANCOVA), while for sweet chestnut they are not statistically different (F 1,20 = 2.30, p = 0.17, ANCOVA).
According to Conover and Iman (1982, p. 716) “However is not uncommon in an ANCOVA to assume that the slope are equal and to test only for equal intercepts”; therefore we performed ANCOVA also for the cases of Norway spruce and of European beech in order to have some additional information in respect to the visual comparison.
Results showed that both for Norway spruce and for European beech RAR the distributions can be considered statistically different between locations only imposing a significance level of 5% (F 1,20 = 4.14, p = 0.05, ANCOVA and F 2,29 = 3.43, p = 0.05, ANCOVA respectively).
The maximum RAR values are generally located in the shallower layers, between 20 and 30 cm, according to species and location (Fig. 10) and ranges between 0.4 and 0.6%. At 1 m depth, RAR is generally on the order of 0.01%, depending on the species.
The average RARs with respect to the entire profile are 0.24% (AG) and 0.07% (MP) for Norway spruce, 0.36% (AG), 0.18% (MP), and 0.09% (M) for European beech, 0.15% (AG) and 0.07% (MP) for European larch, 0.15, 0.15% (AGM) and 0.14% (AB) for sweet chestnut and 0.09% for European hop-hornbeam.
Estimated root cohesion
The values of root cohesion with depth \(\left( {c_{\text{r}}^Z } \right)\) reflected root tensile strength relationships, RAR distributions with depth and the values of k′ and k″. As previous illustrated, k′ was assumed to be 1.0, whereas k″ was estimated as the ratio between root cohesion obtained using the FBM approach and the original W&W model.
For the surveyed trenches, the obtained values of k″ varied between 0.32 and 1.00, predominantly as function of the number of roots (Fig. 11) and secondarily as a function of the heterogeneity of root diameter. It can be observed that k″ is greater than 0.5 for a root density less than 400 roots/m2 and it is between 0.5 and 0.32 for a root density between 400 and 1200 roots/m2. Because also in the case of shallow landslides phenomena the sliding layers are generally deeper than 0.5 m and then they are permeated by a small number of roots, we adopted a cautionary average value of 0.5. Due to the multiplicative form of the equation used for root cohesion estimation, the use of a fixed value for k″ will make future comparison to these results easier.

Values of k″ factor as function of the density of roots
The estimated values of root cohesion along soil profiles substantially reflected the RAR distributions, although some discrepancies could occur due to dependencies on root tensile strength and root diameter distribution.
As expected and according to RAR patterns, root cohesion values showed significant variability within the same species at a given location, but generally decreased with depth. Based on the site-averaged values, there were differences between locations in the cases of the Norway spruce, European larch and European beech, whereas similarity was observed for the sweet chestnut (Fig. 12).

a total root cohesion distribution for a P. abies (L.) Karst., b L. decidua Mill., c F. sylvatica L., d C. sativa Mill, e O. carpinifolia Scop. b Site-averaged (symbols) and specie-averaged (bars) values of root cohesion with depth for a P. abies (L.) Karst, b L. decidua Mill., c F. sylvatica L., d C. sativa Mill, e O. carpinifolia Scop
Site-averaged root cohesion values varied for Norway spruce between 58.6 kPa in the first layers and 14.3 kPa at the maximum depth of 90 cm at AG, and between 41.2 and 4.7 kPa at 130 cm at MP (Fig. 12). Profile-averaged values were thus 35.4 kPa at AG and 13.8 kPa at MP.
Similarly, European beech root cohesion was greater, but less deep, at AG compared to at MP. In the first case, root cohesion was greater than 100 kPa up to 40 cm and then suddenly decreases to about 40 kPa at the maximum depth of 70 cm; in the second case, root cohesion was around 40 kPa up to 50 cm and then decreased gradually to 11 kPa at the maximum depth at 1 m. At M, the maximum value of root cohesion was about 30 kPa at 20–30 cm and the minimum was about 5 kPa at 110 cm. Profile-averaged values were 86 kPa at AG, about 14.4 kPa at M and about 26.6 kPa at MP.
The European larch showed similar behaviour at the two locations, but had different values. The maximum values were reached in the first 20–30 cm, whereas the minimum values were reached at the maximum depth of 110 cm at both locations; these values are, respectively, in the order of 60 kPa and 15 kPa for AG and 30 kPa and 7 kPa for MP. The profile-averaged values are thus 38.3 kPa at AG and 17.4 kPa at MP.
For sweet chestnut, the values of root cohesion were similar for the two locations, but the root depths were different. At AB, the maximum value was 19.0 kPa at 30 cm and 6.4 kPa at 130 cm, whereas at AGM, the maximum value was about 19.6 kPa in the first layer and 8.1 kPa at 90 cm. The profile-averaged values were about 15 kPa for both locations (15.4 and 15.2 kPa respectively).
In the case of European hop-hornbeam, the maximum value of root cohesion was in the order of 30 kPa in the 20 cm and decreased gradually to 5.4 kPa at 120 cm. The profile-averaged value was 14.6 kPa.
Discussion
Root tensile strength
The results obtained for the considered species confirm the validity of the general power law equation for the relationship between root tensile strength and root diameter, in agreement with many other authors (Burroughs and Thomas 1977; Abe and Iwamoto 1986; Gray and Sotir 1996; Nilaweera and Nutalaya 1999; Bischetti et al. 2005; Genet et al. 2005).
Statistical analyses showed that, for the considered species and locations, tensile strength of sampled roots was not affected by location and can be considered a species-dependent parameter. Genet et al. (2006), on the contrary, found a significant difference for the same species sampled at different locations; in their case, however, the difference in elevation between the locations (in south-east Tibet) was very high with respect to the sites considered in the present study, and the Alpine environment in general. The difference in tensile strength, in fact, can be partially associated with a difference in cellulose content (Genet et al. 2005; 2006) and such an effect can possibly only be appreciated for very different environmental conditions, which Alpine species possibly do not encounter. The influence of local factors, like wind, exposure, slope or position of roots in respect of the main stem (upslope or downslope), could of course play a role in determining roots tissue composition. Our data, however, cannot support such hypotheses and they show that the tensile strength values of roots from different sites are not different from a statistical point of view.
The analysis of the tensile strength showed that there is a significant statistical difference between the considered species, in agreement with Genet et al. (2005). It can be observed (Fig. 4) that European beech seems to be the strongest species for small and medium-sized roots, followed by European larch, Norway spruce, sweet chestnut and European hop-hornbeam, which seem to be similar.
Statistical analysis showed that European hop-hornbeam and sweet chestnut on one hand and the two conifers on the other can be described by the same regression function. The differences or the similarities among tensile strength-diameter relationships for different species, should reflect differences and analogies in root anatomy and composition, which in turn should be related to the interaction between the genetic properties of species and the response to ecological conditions; also management could possibly exert a significant role. The interpretation of similar/different tensile strength-diameter relationships of different species, however, is a great challenge and requires further investigations and more thoughts based on an interdisciplinary approach.
Without clear evidences on such a point, in the present work root cohesion was estimated using the relationships specifically developed for each species.
Regression parameters obtained by Genet et al. (2005) for Norway spruce, sweet chestnut and European beech, in fact, were quite different from those obtained in the present study. According to the same authors, however, we deem that such differences can be ascribed to the presence of small roots (less than 0.9–1.0 mm). These small roots generally show very high strength values, a great variability, and, thus, a dramatic influence on the regression curves; very small roots, for example, generally involve a greater degree of uncertainty in measuring their diameter and then in calculating strength values.
The influence of fine roots on tensile strength-diameter relationships must be further investigated, with specific reference to the possible consequences on root cohesion estimation. The contribution of very fine roots (<1 mm) to soil resistance is questionable due to the length needed to avoid root slipping (Waldron 1977). Currently, only roots greater than 1 mm are generally considered in studies dealing with rooted soil reinforcement (Reubens et al. 2007), although there is no evidence of a well-defined critical threshold size.
RAR distribution
The RAR results showed that profile-averaged values vary between 0.07% and 0.36% and they are in the range of the values reported by other authors for different species and locations (Wu 1995; Stokes et al. 2008).
As expected, a great variability was observed within species with regard to depth, trench at the same location and location (Fig. 5–9). Such variability is due to the spatial heterogeneity of root system development, which is dependent upon the interactions of genetic and environmental factors (Stokes et al. 2008).
In spite of such variability, however, general patterns of RAR with depth (represented by the site-averaged RAR values) can be identified.
The general (and obvious) pattern is that the RAR tends to decrease with depth, showing maximum values on the order of 0.5% at a depths of 20–40 cm and minimum values of an order of magnitude less at the maximum depth (Fig. 10). When RAR, and root density in general, are used to estimate the root contribution to stability, then the use of average values should be avoided because they can lead to a dramatic overestimation of the additional cohesion at the sliding surface.
The maximum depth reached by roots in most cases was between 0.8 and 0.9 m and each of the considered species had at least one excavated trench over 1 m (average rooted depth of 84 cm ± 21, and maximum rooted depth of 1.3 m). The species-averaged rooted depths were around 90 cm, except for Norway spruce (73 cm). In general, according to Schiechtl (1980), in the Alpine environment 1 m can be considered a good reference depth for rooted soils.
The relationship between RAR and depth show different patterns between locations for the European larch and partially for the European beech and Norway spruce. For sweet chestnut, on the contrary, it is clear that RAR distributions of the two locations are statistically not different.
It must be noted that such differences always involve Alpe Gigiai trench points (AG). In general, at AG, the RAR values are greater with respect to the other locations but are concentrated in the upper portion of soil, with particular reference to European beech. Such behaviour is not easily explained with the available information. On one hand, at AG, soils are shallower and coarser than those at the other locations, which could keep roots in the first layers and hinder deeper penetration. On the other hand, at AG, the trees are about half the age of those at the other locations, and the effect of age on RAR and diameters should be investigated.
In the case of sweet chestnut trees, which were 20 and 30 years old, RAR distributions with depth were very similar for the two locations, although a small difference was observed in rooted depth.
Root cohesion distribution
Due to the multiplicative form of Eq. 6a and 6b, the values of the coefficients k′ and k″ assume a crucial role in estimating root cohesion. The value of k′ depends on the friction angle of the soil, ϕ, and the distortion angle of sheared roots, θ (see Eq. 1). It can be noted that for ϕ greater than 25° (which describes most soils; Day 2001) k′ is less than unity when θ is less than 40°; such conditions, however, occur only for roots characterised by high values of stiffness and diameter (Abe and Ziemer 1991; Wu 1995). Danjon et al. (2008), by means of 3-D measures of two trees of white oak (Quercus alba L.) and assuming the actual angle of intersection between roots and the potential shear surface as θ, found that the average value of k′ at different depths was always greater than 1.0 (1.03–1.13) for ϕ = 30°, whereas k′ values range between 0.88 and 1.02 for ϕ = 20°; the value of 1.2 is given only for ϕ = 40° (1.18 < k′ < 1.28). In any case, the value of unity for k′ assumed herein allows for adaptation of results when ϕ and/or θ are known.
Many authors have demonstrated that k″ can have values much lower than 1.0, dramatically affecting the value of root reinforcement (Waldron and Dakessian 1981; Operstein and Frydman 2000; Pollen and Simon 2005; Docker and Hubble 2008). The FBM approach, as already illustrated can account for the processes affecting k″ and be used to estimate its values. According to Pollen and Simon (2005), the magnitude of k″ estimated by the FBM approach is strongly associated with the number of roots considered. In the case of the surveyed trenches, k″ is always greater than 0.5 for a density smaller than 400 roots/m2 (Fig. 11), similar to the value suggested by Hammond et al. (1992) for forest species.
The site and profile-averaged values of root cohesion varied between about 15 and 80 kPa, according to the values reported in the literature (for a recent overview see Norris et al. 2008b).
Estimated values of root cohesion along soil profiles substantially reflected the RAR distribution patterns, although some discrepancies could occur due to the dependency on root tensile strength and root diameter distribution.
In general, the root cohesion values herein estimated are several tens of kPa in the first layers (10–40 cm) and still are significant at depths of more than 1 m (Fig. 12). Studies of root cohesion distribution with depth are scarce. Such values, however, can be considered consistent with the values reported in the literature for direct in situ shear tests and back-analysis, which generally are around a few kPa and seldom just less than 20 kPa (Wu 1995; Norris et al. 2008a), but lower with respect to those sometimes obtained by the W&W model (Schmidt et al. 2001). Most shear tests, in fact, excavating all around the soil block to test, measured root cohesion at a depth of 20–40 cm, but only for the basal component; Wu et al. (1988), who carried out direct shear test on lateral root cohesion, measured values of additional cohesion on the order of tens of kPa. Back-analysis, on the other hand, considers the cohesion of the entire profile at the shear surface, but likely at those points of the hillslopes where the root reinforcement is weaker. The values obtained with the W&W model can overestimate the root cohesion when the coefficients k′ and k″ are incorrectly evaluated. The values obtained in the present study, on the contrary, consider both basal and lateral cohesion (see Eqs. 6a and 6b) and are obtained in healthy forests, but reflect a cautionary estimation of k′ and k″.
In the case of European beech, the influence of root size in determining the root cohesion is particularly evident; the difference between site-average root cohesion values with depth are much greater than the difference between the associated RAR distributions. The very high values at AG can be attributed both to higher root density and, perhaps mainly, to the large amount of small roots (less than 3 mm), compared to the other two locations. The cause of such a structure is not clear and the point should be further investigated, considering different locations, tree ages and management practices.
European larch shows a greater reinforcement at all depths, with respect to other species (except European beech at AG in the first layers), due to a combination of a dense root system and high root strength. Considering the deeper layers, which are more relevant to hillslope stabilization, all the considered species contribute significantly. Considering a depth of 1 m as a reference for the use of vegetation for terrain stabilization in the Alps, the strongest species is the European larch (12.75 kPa), followed by the European beech (9.34 kPa not considering AG values), the European hop-hornbeam (6.30 kPa), the sweet chestnut (5.75 kPa) and the Norway spruce (2.56 kPa). Single trenches, however, can show greater values (e.g. 7.47 kPa at a depth of 120 cm for one trench of Norway spruce).
Conclusions
Measures of root tensile strength and Root Area Ratio and an estimation of root cohesion at different depth are provided for the Norway spruce (Picea abies (L.) Karst.), European larch (Larix decidua Mill.), European beech (Fagus sylvatica L.), sweet chestnut (Castanea sativa Mill.) and European hop-hornbeam (Ostrya carpinifolia Scop.) for some locations in the Lombardy Alps and Prealps.
Two conifers, Norway spruce and European larch, and two broadleaves, European hop-hornbeam and sweet chestnut, species showed statistically similar strength-diameter relationships. For the two couples and another broadleaf, European beech, on the contrary the strength-diameter relationships resulted statistically different. The comprehension of the causes of such differences and similarities were out of the goals of the work and they should be investigated in the future. The results obtained herein put the basis for such a work.
RAR distributions with depth, showed a great in-site variability. If site-averaged values were considered, however, RAR distributions resulted statistically different among the considered locations at 1% level of significance in the case of European larch and not different in the case of sweet chestnut. For Norway spruce and European larch the results did not show a clear response and future investigations should explore the possibility to include in the analyses local factors (soil depth, fertility, steepness, aspect, etc.).
Root cohesion values were estimated by means of a method based on the Wu (1976) and Waldron (1977) approach, implemented to account for roots of different size, root density distribution with depth and non-simultaneous breaking of roots. Since the method has been applied to forest species, where the soil is permeated by roots of different trees, and the most common sliding phenomena are shallow with a small volume, the total root cohesion was considered to be the sum of basal cohesion (due to roots present at shearing surface) and lateral cohesion (due to roots present along the whole rooted profile). The proposed method allows for estimation of the additional root cohesion for the considered species at different depths, whereas direct shear tests only give the basal and the lateral root cohesion at a defined depth (generally only few tens of cm) and back-analysis provides averaged values at sites that are probably weak. The FBM approach leads to a modified method that overcomes the simultaneous root breaking hypothesis of the original W&W model. The results obtained encourage further development of a model that implicitly incorporates FBM principles in order to estimate the actual root cohesion with respect to depth.
Estimated root cohesion values, according to RAR, show a great variance around the mean, even for the same species, depth, and location. When the site averaged values are considered, however, a clear trend can be observed. For most of the considered species, a great influence is also exerted by the location and perhaps by the local management practices. Currently, there is not enough data to completely explain this variance and this must be considered when root cohesion is added to soil cohesion in modelling hillslope stability.
Results show that, although most of the roots are concentrated in the first layers, forest vegetation can significantly increase soil cohesion at rather deep layers (over 1 m), accordingly to species. This effect is controlled by the root density and, above all, the root strength, which in turn, depends on species and root size.
In conclusion, the results obtained in the present work demonstrate that the revised W&W model proposed herein can be considered as an adequate tool for estimating root cohesion at the sliding surface, overcoming some limitation of the original scheme and maintaining its simplicity. The method is suitable for inclusion in stability models to quantitatively estimate how forests can be used to protect the mountainous landscape from hydrogeomorphic hazards, with particular reference to shallow landslides.
Moreover, the results provide input data for stability models including some of the most common forest species in the Alps and, in general, contribute to increase the knowledge about the reinforcement action of European Alpine vegetation. This further reveals the relationship between protection forests and mountain landscape stability.
References
Abe K, Iawamoto M (1986) An evaluation of tree-root effect on slope stability by tree-root strength. J Jpn For Soc 68:505–510
Abe K, Ziemer RR (1991) Effect of tree roots on a shear zone: modeling reinforced shear stress. Can J Res 21:1012–1019 doi:10.1139/x91-139
Abernethy B, Rutherfurd ID (2001) The distribution and strength of riparian tree roots in relation to riverbank reinforcement. Hydrol Process 15:63–79 doi:10.1002/hyp.152
Babu J, Herendra NP, Radhey ST (2001) Vertical distribution and seasonal changes of fine and coarse root mass in Pinus kesiya Royle Ex.Gordon forest of three different ages. Acta Oecol 22:293–300 doi:10.1016/S1146-609X(01)01118-3
Bischetti GB, Bonfanti F, Greppi M (2003) Misura della resistenza alla trazione delle radici: apparato sperimentale e metodologia d’analisi. Quad Idronomia Mont 241(1) (in Italian)
Bischetti GB, Chiaradia EA, Simonato T et al (2005) Root strength and root area ratio of forest species in Lombardy (Northern Italy). Plant Soil 278:11–22 doi:10.1007/s11104-005-0605-4
Burke MK, Raynal DJ (1994) Fine root growth phenology, production and turnover in a northern hardwood forest ecosystem. Plant Soil 162:135–146 doi:10.1007/BF01416099
Burroughs ER, Thomas BR (1977) Declining root strength in Douglas-fir after felling as a factor in slope stability. USDA For Serv Res Pap INT-190, 27 pp
Büttner V, Leuschner C (1994) Spatial and temporal patterns of fine root abundance in a mixed oak beech forest. For Ecol Manag 70:11–21
Daniels HE (1945) The statistical theory of the strength of bundles of threads. Proc R Soc Lond A Math Phys Sci 183:405 doi:10.1098/rspa.1945.0011
Danjon F, Barker DH, Drexhage M, Stokes A (2008) Using 3D plant root architecture in models of shallow slope stability. Ann Bot (Lond) 101:1281–1293 doi:10.1093/aob/mcm199
Day RW (2001) Soil Testing Manual: Procedures, Classification Data, and Sampling Practices. McGraw-Hill Professional
De Baets S, Poesen J, Reubens B et al (2008) Root tensile strength and root distribution of typical Mediterranean plant species and their contribution to soil shear strength. Plant Soil 305:207–226
Docker BB, Hubble TCT (2008) Quantifying root-reinforcement of river bank soils by four Australian tree species. Geomorphol doi:10.1016/j.geomorph.2008.01.009
Ekanayake JC, Phillips CJ (1999) A method for stability analysis of vegetated hillslopes: an energy approach. Can Geotech J 36:1172–1184 doi:10.1139/cgj-36-6-1172
Endo T, Tsuruta T (1969) Effect of trees’ roots upon the shearing strength of soil. 18th Annual Rpt of the Okkaido Branch. For Exp Stn, Tokio, Japan, 167–169
Frydman S, Operstein V (2001) Numerical simulation of direct shear of root-reinforced soil. Ground Improv 5:41–48 doi:10.1680/grim.5.1.41.39437
Genet M, Stokes A, Salin F et al (2005) The influence of cellulose content on tensile strength in tree roots. Plant Soil 278:1–9 doi:10.1007/s11104-005-8768-6
Genet M, Stokes A, Fourcaud T et al (2006) Effect of altitude on root mechanical and chemical properties of Abies georgei in Tibet, 5th Plant Biomech Conf—Stockholm, August 28–September 1 2006
Gray DH, Laiser AJ (1982) Biotechnical slope protection and erosion control. Van Nostrand Reinhold, New York. USA
Gray DH, Sotir RB (1996) Biotechnical Soil Bioengineering Slope Stabilization: A Practical Guide for Erosion Control. Wiley, New York, USA
Greenwood JR (2006) SLIP4EX—a program for routine slope stability analysis to include the effects of vegetation, reinforcement and hydrological changes. Geotech Geol Eng 24:193–202
Hammond C, Hall D, Miller S, Swetik P (1992) Level I, stability analysis (LISA) documentation for version 2.0. General Technical Report INT-285. USDA For Serv Intermt Res Stn
Hathaway RL, Penny D (1975) Root strength in some Populus and Salix clones. NZ J Bot 13:333–343
Hemmer PC, Hansen A, Pradhan S (2007) Rupture processes in Fiber Bundle Models. In: Bhattacharyya P, Chakrabarti BK (eds) Modeling Critical and Catastrophic Phenomena in Geoscience, Lecture Notes in Physics No. 705. Springer, Berlin, pp 27–55
Hendriks CMA, Bianchi FJJA (1995) Root density and root biomass in pure and mixed forest stands of Douglas-fir and Beech. Neth J Agric Sci 43:321–331
Keim RF, Skaugset AE (2003) Modelling the effect of forest canopies on slope stability. Hydrol Process 17:1457–1467 doi:10.1002/hyp.5121
Kun F, Raischel F, Hidalgo RC, Herrmann HJ (2007) Extension of Fiber Bundle Models. In: Bhattacharyya P, Chakrabarti BK (eds) Modeling Critical and Catastrophic Phenomena. in Geoscience, Lecture Notes in Physics No. 705. Springer, Berlin, pp 57–92
Lopez-Zamora I, Falcão N, Comerford NB, Barros NF (2002) Root isotropy and an evaluation of a method for measuring root distribution in soil trenches. For Ecol Man 166:303–310
Meyer FH, Göttsche D (1971) Distribution of root tips and tender roots of beech. In: Ellenberg H (ed) Ecological studies. Analysis and synthesis, vol. 2. Springer-Verlag, Berlin, pp 47–52
Morgan RPC, Rickson RJ (1995) Slope stabilization and erosion control—a bioengineering approach. Chapman & Hall, Univ Press, Cambridge, pp 221–264
Nilaweera NS, Nutalaya P (1999) Role of tree roots in slope stabilisation. Bull Eng Geol Environ 57:337–342 doi:10.1007/s100640050056
Normaniza OH, Faisal A, Barakbah SS (2008) Engineering properties of Leucaena leucocephala for prevention of slope failure. Ecol Eng 32:215–221 doi:10.1016/j.ecoleng.2007.11.004
Norris JE (2005) Root reinforcement by hawthorn and oak roots on a highway cut-slope in Southern England. Plant Soil 278:43–53 doi:10.1007/s11104-005-1301-0
Norris JE, Stokes A, Mickowski SB et al (2008a) Slope Stability and Erosion Control: Ecotechnological Solutions, Springer
Norris JE, Greenwood JR, Achim A et al (2008b) Hazard assessment of vegetated slopes. In Norris JE et al (ed) Slope stability and erosion control: Ecotechnological solutions. Springer, 119–166
Operstein V, Frydman S (2000) The influence of vegetation on soil strength. Ground Improv 4:81–89
Pollen N, Simon A (2005) Estimating the mechanical effects of riparian vegetation on streambank stability using a fiber bundle model. Water Resour Res 41:W07025 doi:10.1029/2004WR003801
Reubens B, Poesen J, Danjon F et al (2007) The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: a review. Trees (Berl) 21:385–402 doi:10.1007/s00468-007-0132-4
Raischel F, Kun F, Herrmann HJ (2008) Continuous damage fiber bundle model for strongly disordered materials. Phys Rev E Stat Nonlin Soft Matter Phys 77:046102 doi:10.1103/PhysRevE.77.046102
Riestenberg MM (1994) Anchoring of thin collovium by roots of sugar maple and with ash on hillslopes in Cincinnati. US Geol Surv Bull 2059-E
Riestenberg MM, Sovonik-Dunford S (1983) The role of woody vegetation in stabilizing slopes in the Cincinnati area Ohio. Geol Soc Am Bull 94:506–518 doi:10.1130/0016-7606(1983)94<506:TROWVI>2.0.CO;2
Roering JJ, Schmidt KM, Stock JD et al (2003) Shallow landsliding, root reinforcement, and the spatial distribution of trees in the Oregon Coast Range. Can Geotech J 40:237–253 doi:10.1139/t02-113
Sakals ME, Sidle RC (2004) A spatial and temporal model of root cohesion in forest soils. Can J Res 34:950–958 doi:10.1139/x03-268
Schiechtl HM (1958) Grundlagen der Grunverbauung. Mitteilungen der Forstlichen bundes-versuchsanstalt Mariabrunn. 55 Heft. 273 pp. (in German)
Schiechtl HM (1980) Bioengineering for land reclamation and conservation. Univ of Alberta Press, Edmonton, Canada
Schmid I, Kazda M (2001) Vertical distribution and radial growth of coarse roots in pure and mixed stands of Fagus sylvatica and Picea abies. Can J Res 31:539–548 doi:10.1139/cjfr-31-3-539
Schmid I, Kazda M (2002) Root distribution of Norway spruce in monospecific and mixed stands on different soils. For Ecol Man 159:37–47
Schmidt KM, Roering JJ, Stock JD et al (2001) The variability of root cohesion as an influence on shallow landslide susceptibility in the Oregon Coast Range. Can Geotech J 38:995–1024 doi:10.1139/cgj-38-5-995
Sidle RC, Ochiai H (2006) Landslides, Processes, Prediction, and Land Use Water Resour Monograph 18, AGU, Washington D.C., USA
Stokes A, Norris JE van Beck LPH et al (2008) How vegetation reinforces the soil on slopes. In Norris JE et al (eds) Slope stability and erosion control: Ecotechnological solutions. Springer, pp 65–118
Sun HL, Li SH, Xiong WL et al (2008) Influence of slope on root system anchorage of Pinus yunnanensis. Ecol Eng 32:60–67 doi:10.1016/j.ecoleng.2007.09.002
Terwillinger VJ, Waldron LJ (1991) Effects of root reinforcement on soil-slip patterns in the Transverse Ranges of southern California. Geol Soc Am Bull 103:775–785 doi:10.1130/0016-7606(1991)103<0775:EORROS>2.3.CO;2
Tosi M (2007) Root tensile strength relationships and their slope stability implications of three shrub species in the Northern Apennines (Italy). Geomorph 87:268–283 doi:10.1016/j.geomorph.2006.09.019
van Beek LP, Wint H, Cammeraat LH, Edwards JP (2005) Observation and simulation of root reinforcement on abandoned Mediterranean slopes. Plant Soil 278:55–74 doi:10.1007/s11104-005-7247-4
Vinceti B, Paoletti E, Wolf U (1998) Analysis of soil, roots and mycorrhizae in a Norway spruce declining forest. Chemospere 36:937–942 doi:10.1016/S0045-6535(97)10151-5
Vogt KA, Persson H (1991) Measuring growth and development of roots. Techniques and approaches in forest tree ecophysiology. CRC, Boca Raton
Waldron LJ (1977) The shear resistance of root-permeated homogeneous and stratified soil. Soil Sci Soc Am J 41:843–849
Waldron LJ, Dakessian S (1981) Soil reinforcement by roots: calculation of increased soil shear resistance from root properties. Soil Sci 132:427–435 doi:10.1097/00010694-198112000-00007
Wu TH (1976) Investigation on landslides on Prince of Wales Island. Alaska Geotech Rpt No 5 Dpt Civ Eng Ohio State Univ Columbus, USA
Wu TH (1995) Slope Stabilization. In: Morgan RPC, Rickson RJ (eds) Slope Stabilization and Erosion Control-A Bioengineering Approach. Chapman & Hall, Univ Press, Cambridge, pp 221–264
Wu TH, McKinnell WP III, Swanston DN (1979) Strength of tree roots and landslides on Prince of Wales Island, Alaska. Can Geotech J 16:19–33
Wu TH, Beal PE, Lan C (1988) In-situ shear tests of soil-root system. J Geotech Eng ASCE 114:1376–1394 doi:10.1061/(ASCE)0733-9410(1988)114:12(1376)
Xu YJ, Röhrig E, Fölster H (1997) Reaction of root systems of grand fir (Abies grandis Lindl.) and Norway spruce (Picea abies Karst.) to seasonal waterlogging. For Ecol Man 93:9–19
Yazici B, Yolacan S (2007) A comparison of various tests of normality. J Stat Comput Simul 77:75–183 doi:10.1080/10629360600678310
Ziemer RR (1981) Roots and stability of forested slopes. In Davies TRH, Pearce AJ (eds) Erosion and sediment transport in Pacific Rim steeplands. Int Assoc Hydrol Sci Pub 132:343–361
Acknowledgements
The research activity is part of the Research Program SISIFO funded by Regione Lombardia—Agricultural Directorate. The Authors wish to thank Dr. Giulio Zanetti for the field support, Dr. Eric Spelta for helping in some data elaboration, Prof. Claudio Gandolfi for the profitable discussions and the two anonymous reviewers for their helpful suggestions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Lars S. Jensen.
Rights and permissions
About this article
Cite this article
Bischetti, G.B., Chiaradia, E.A., Epis, T. et al. Root cohesion of forest species in the Italian Alps. Plant Soil 324, 71–89 (2009). https://doi.org/10.1007/s11104-009-9941-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-009-9941-0