
Abstract
Selenium (Se) is essential for humans and animals but is not considered to be essential for higher plants. Although researchers have found increases in vegetative growth due to fertiliser Se, there has been no definitive evidence to date of increased reproductive capacity, in terms of seed production and seed viability. The aim of this study was to evaluate seed production and growth responses to a low dose of Se (as sodium selenite, added to solution culture) compared to very low-Se controls in fast-cycling Brassica rapa L. Although there was no change in total biomass, Se treatment was associated with a 43% increase in seed production. The Se-treated Brassica plants had higher total respiratory activity in leaves and flowers, which may have contributed to higher seed production. This study provides additional evidence for a beneficial role for Se in higher plants.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Lower plants such as algae need Se for normal growth and development, with species from six classes having a Se requirement such that the element is either essential for growth or strongly promotes it (Price et al. 1987). The dinoflagellate Peridineum cinctum can even be used as a bioassay for Se in freshwater lakes (Lindstrom 1983), while selenate can induce activity of the important antioxidant selenoenzyme glutathione peroxidase (GSH-Px) in the green alga Chlamydomonas reinhardtii (Yokota et al. 1988).
For vascular (higher) plants, however, the question of Se’s essentiality remains unresolved after nearly 70 years of investigation. Selenium-deficient soils appear neither to inhibit plant growth nor to reduce crop yield (Reilly 1996; Shrift 1969). Some studies have demonstrated benefits of adding small amounts of Se, including increased tuber yield and increased starch concentration in young leaves in potato (Solanum tuberosum L.) (Turakainen et al. 2004), and increased growth in ryegrass (Lolium perenne L.) and lettuce (Lactuca sativa L.) exposed to UVB radiation (Hartikainen and Xue 1999). This response was associated with inhibition of lipid peroxidation through increased GSH-Px activity (Xue and Hartikainen 2000). Other studies have shown that Se can reduce lipid peroxidation at lower doses (Cartes et al. 2005; Djanaguiraman et al. 2005; Hartikainen et al. 2000), while acting as a pro-oxidant at higher doses (Hartikainen et al. 2000).
When a major selenoprotein was discovered in mung bean (Vigna radiata L.) seedlings supplemented with 2 mg L−1 of selenite, a role for Se in mitochondrial membrane functions was indicated (Easwari and Lalitha 1994). Mitochondrial GSH-Px was found to increase three-fold when fenugreek (Trigonella foenum-graecum L.) plants were supplemented with Se (Sreekala et al. 1999). It has been argued, however, that molecular evidence is yet to support the conclusion that any of these enzymes are true selenoproteins (Terry et al. 2000). On the other hand, further evidence for a Se role in normal plant metabolism was indicated by the discovery that a cysteine desulphurase NifS-like protein may be involved in selenoprotein synthesis in chloroplasts (Pilon-Smits et al. 2002), which, along with mitochondria, are subject to high levels of oxidative stress.
Despite the finding of increased vegetative growth in several species associated with added Se (Djanaguiraman et al. 2005; Hartikainen and Xue 1999; Turakainen et al. 2004; Xue et al. 2001), there is no definitive evidence to date in the published literature of increased reproductive capacity, in terms of seed production and viability. This is an important measure in evolutionary terms and in the production of cereals, oilseeds and legumes.
After promising results from trials using several plant species, the present study was conducted to evaluate seed production and growth response to the addition of Se to B. rapa in a carefully conducted hydroponic trial. Also, in view of the findings of Girton (1974) and Smrkolj et al. (2006) of increased respiration associated with added Se, we investigated the effect of Se addition on respiration and whether this is associated with seed production.
Materials and methods
Phytotron trial
A hydroponic trial was conducted in two phytotrons to examine the effect of selenite on a fast-cycling Brassica rapa cultivar (University of Wisconsin, Madison, Wisconsin). This plant has been selected to flower and set seed early and can produce seed in as little as 6 weeks. Sodium selenite was preferred as it was the most effective Se form for increasing seed production in B. rapa and Arabidopsis thaliana and in increasing early root growth in wheat in our earlier trials. The seeds used in this trial were derived from one plant, one of the control (low-Se) plants grown in a pilot trial, with a Se concentration of around 10 μg kg−1 DW. The seeds were surface sterilised with 70% ethanol and 4% hypochlorite (1 min and 5 min respectively) and germinated on Whatman ashless filter paper soaked in Milli-Q water, in 15 cm plastic petri dishes.
After 48 h, germinated seeds (with 1 cm radicles) were transferred to cotton wicks inside truncated Eppendorf tubes, one per tube, and four tubes were placed in the lids of each 1.1 l black polyethylene pot, which had been carefully acid-washed (5% nitric acid). Each pot contained 0.125-strength modified Hoagland solution (Epstein and Bloom 2005). On day 6, the largest and most uniform plants were selected and transplanted, one per pot, and randomised to 30 controls and 30 to be Se treated. The control and treated plants were placed in separate phytotrons to preclude transfer of volatile Se compounds from the treatments to the controls (Terry et al 2000). Aeration was provided to the solution in each pot. The conditions in each phytotron were closely monitored to ensure that the two groups of plants grew under identical conditions, which were: 23/15°day/night temperature, 14/10 h light/dark cycle, 65% mean relative humidity, and 280 μmol m−2 s−1 light intensity at plant height. Lighting was provided by four white 400 W metal halide light bulbs (OSRAM, Powerarc®, M95E, MIS400/BU-ONLY, USA).
To further verify the uniformity of the growth conditions, four pots of University of California (UC) mix growth medium (Barker et al. 1998), each containing four barley seedlings of equal size, were placed in each phytotron and harvested after 20 d. The mean (sd) whole tops dry weight (DW) per pot for barley grown in the control phytotron was 1.99 (0.09) g, compared with 1.93 (0.12) g for barley grown in the Se-treatment phytotron, i.e. they were essentially the same.
Throughout the trial, solutions were changed on days 6, 17, 23, 30, 37, 45, 52 and 58. At each solution change, the pots were re-arranged to reduce position effects in each phytotron. As the plants grew, solutions were topped-up with Milli-Q water between solution changes to replace transpiration losses. On day 17 the Hoagland’s nutrient solution concentration was increased from 0.125-strength to 0.25-strength, and on day 23 the concentration was further increased to 0.5-strength. This concentration was maintained for the rest of the trial. The 0.5-strength modified Hoagland’s solution contained: 4 mM KNO3; 2 mM Ca(NO3)2; 0.2 mM NH4H2PO4; 0.5 mM MgSO4.7H2O and micronutrients (μM): KCl (100), H3BO3 (25), MnSO4H2O (0.8), ZnSO4.7H2O (1), CuSO4.5H2O (0.2), HO2MoO4 (0.2), NiSO4.6H2O (0.2), CoSO4.7H2O (0.05), Na2SiO3.9H2O (500), NaFe ethylenediaminetetraacetic acid (EDTA) (10% Fe) (20). The pH of the solution was buffered at 6.0 by MES (2-[N-Morpholino] ethane-sulphonic acid)-KOH (MES 2 mM, KOH 0.872 mM).
Sodium selenite was added to the solution in the Se-treatment pots immediately after solution change on day 6 (22 μL of 0.001 M selenite added to 1.1 L of solution). Thereafter, 55 μL of 0.001 M selenite was added on days 12, 17, 23, 30 and 37. These concentrations and the frequency of Se supplementation were determined from the results of our previous trials using A. thaliana and B. rapa to maximise Se benefit to the plants. A dose-response trial using different amounts of added selenite (other than no dose v estimated optimal dose) was not conducted as growth room space was limited and a large number of pseudoreplications was desired. All operations were performed on control plants first, to avoid Se contamination from the treated plants.
In order to find whether Se addition affected leaf mitochondrial activity, leaf tissue samples (youngest and next-youngest mature leaves) were collected from a random sample of six controls and six Se-treated plants after 20 days, leaving 24 control and 24 treatment plants for measuring yield and Se concentration. Leaves were sliced into approximately 1 mm strips to yield between 0.05 and 0.10 g fresh weight, placed at once in assay buffer (15 mM trans—epoxysuccinate (TES), 0.2 mM CaCl2, pH 7.0) and incubated in the dark for 10 min to allow wound respiration to subside. Oxygen uptake was monitored using a Clark electrode with 3.0 mL of fresh assay buffer and black cloth to exclude light. Additions made to the assay were 5 mM KCN to inhibit the cytochrome pathway and 1 μM octyl-gallate to inhibit the alternate pathway. The total respiratory rate of the tissue was determined as being the respiratory rate in the absence of inhibitors minus the residual rate, which is the rate in the presence of both inhibitors.
Flower samples were collected from six randomly selected controls and treatments 22 days after germination. The relative amount of cytochrome oxidase protein present in flower extracts of control and Se-treated plants was determined using immunoblot method. Frozen tissue (0.1 g) was ground to a powder and double-strength SDS-PAGE sample buffer was added (300 µL). Samples were boiled for 5 min and centrifuged at 14,000 g for 10 min. Supernatant samples (20 µL, 6.7 mg protein) were loaded on a Laemmli-type 10% SDS-PAGE gel. Proteins were subsequently blotted onto nitrocellulose by standard methods. COX protein was detected using a primary anti-COX II antiserum from Agrisera, Sweden at 1:1,000 dilution in Tris-buffered saline (10 mM Tris, 120 mM NaCl, 5% Tween) for 1 hr at room temp. After extensive washing, the blot was incubated with 1:1,000 dilution of pig anti-rabbit IgG horseradish peroxidise conjugate, (DakoCytomation, CA) and the signal was detected using chemiluminescence.
Because this Brassica cultivar is self-incompatible, mechanical transfer of pollen between plants is required to achieve pollination. From day 26, pollination using a small artist’s brush was performed twice weekly on each flower, using pollen from another plant.
Pollen samples were collected on day 27 and stained with acetocarmine dye on slide glass. Pollen grains that stain red have viable cytoplasm. Pollen grains were assessed from three treatments and three controls, and a mean count from three microscope fields per sample was obtained. In addition, pollen collected on days 27 and 47 was examined under a scanning electron microscope to compare morphology of pollen grains from Se-treated and control plants.
Chlorophyll content was measured in a sample of the third-youngest leaves of each plant on day 42 using a SPAD meter (Minolta, Japan-SPAD-502).
The plants were harvested on day 80. Whole tops (including seed pods) were dried in an oven at 30°C for 3 days. At this temperature the seeds would remain viable. Then the seeds were removed, and the remainder of the tops were dried for a further 2 days at 70°C. Roots were dried for 2 days at 70°C.
Selenium was analysed by inductively coupled plasma mass spectrometry (ICP-MS) and Fe, Mn, Cu, Zn, Ca, Mg, Na, K, P and S by ICP optical emission spectrometry (ICP-OES) following acid-digestion (Zarcinas et al. 1987). The variation in recovery of Se from the certified reference material (NIST durum wheat flour 8436) was just 1%. The limit of accurate determination for Se was 4 μg kg−1, calculated as ten times the standard deviation of the blank.
Germination trial
A representative sample of 50 seeds from each B. rapa plant grown in the Se supplementation trial described above was placed on filter paper (Whatman qualitative circles 110 mm diameter) wetted with 10 ml Milli-Q pure water in 140 mm diameter plastic petri dishes. Thus there were 24 samples of seed from Se-treatment plants and 24 samples of seed from low-Se control plants. The dishes were placed on a laboratory bench with 12 h photoperiod and mean temperatures of 22°C day, 14°C night. The number of seeds which had germinated was counted 48 h after commencement of the experiment.
Statistical analysis
The two Se treatments, -Se and +Se, had to be assigned to two separate growth rooms to avoid cross-contamination of -Se plants with volatile Se compounds from +Se plants. Although the growth conditions were similar between the growth rooms, having the two Se treatments in two separate growth rooms resulted in pseudoreplication (Hurlbert 1984), making it impossible to perform proper statistical analysis of the data. Therefore, we only calculated means and standard error of the means. Despite the lack of statistical analysis, the similar growth response of test plant barley grown in the separate growth rooms suggests that differences between growth rooms are likely to be small and differences between Se treatments are worth reporting.
Results
Selenium concentration
Selenium concentrations in seeds, tops and roots were higher in the Se-supplemented plants (Table 1).
Seed production and growth
The Se-treated plants produced, on average, 43% more seed than the low-Se control plants. There was no difference between groups in individual seed weight, total biomass, root biomass or whole top biomass (Table 2).
Mineral nutrients and micronutrients
The mineral nutrients analysed by ICP-OES were all within normal limits in the leaf and seed samples tested, i.e. no nutrient deficiencies or toxicities were apparent. Moreover, there were no differences in concentrations of any mineral nutrient between groups (data not shown). The mean total biomass of all the plants in the experiment (15.63 g) was high (around three times the biomass commonly reported in experiments using these plants) for this fast-cycling cultivar of B rapa, and suggests optimum growing conditions for maximising total biomass.
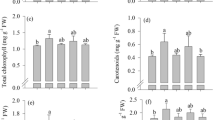
Chlorophyll content
For chlorophyll content measured in young leaves, there was no difference between groups, with a mean (SD, range) of 33.4 (16, 18–46) for controls, and 34.6 (18, 21–48) for treatment.
Pollen
Pollen from the control plants which was sampled and then treated with acetocarmine dye was found to have a mean of 14% unviable grains, compared to a mean of 2% unviable grains for the Se treated plants sampled (Table 3). However, electron microscopy revealed no apparent morphological differences in pollen grains from the two groups.
Respiration
Mitochondria from Se-treated plants showed higher total respiratory activity than control plants both in the leaves (140 (se 11) vs 86 (11) nmol O2 min−1 g−1 fresh weight (FW), a 63% increase) and in the flowers (64 (8) vs 22 (5) nmol O2 min−1 g−1 FW, a 200% increase) (Fig. 1).

Respiration rate in leaves and flowers of Brassica rapa plants grown in solution culture with or without sodium selenite. Vertical bars represent standard errors based on six pseudoreplications
Germination
Seed from selenite-treated plants had a mean (standard error) germination of 92% (1.6), compared to 81% (22) for control seed (Table 3).
Discussion
The experimental method was effective in minimising Se availability to the control plants, as no Se was detected in the control culture solutions or in control seeds or tops (plant tissue sample detection limit of 4 μg kg−1 DW). However, even with the stringent measures taken to exclude Se from the control culture solution, the control plants which were sampled had a detectable Se concentration in their roots (Table 1). It has been suggested that, if Se were essential to vascular plants, an available Se concentration as low as 1.0 μg kg−1 in growth media would be able to satisfy their Se requirement (Terry et al. 2000).
It appears that the Se-treated plants diverted proportionately more resources to seed production. Although the seed production appeared to be different (according to the means and standard errors), there was a high coefficient of variation (43%) despite the high number of replications (24), due largely to having one plant per replication. For example, total seed weight per plant ranged from 0.27–6.40 g in controls and from 1.81–7.19 g in treatments. Thus the results should be treated with caution and further research is warranted.
However, this study is not the only one conducted by our group to examine the effects of a wide range of Se doses on diverse plant species, including Arabidopsis thaliana L., Astragalus sinicus L. (milk vetch), Brassica rapa L. (mustard) and Triticum aestivum L. (wheat). The unpublished findings are similar to those of the current study and include: a 72% increase in A. thaliana fertile silique number (treatment mean [se] 193 [13] vs 112 [12] for controls) for plants treated with 0.003 µM selenite compared with low-Se controls, in a hydroponic trial. In A. sinicus the addition of 10 µg kg−1 Se as sodium selenate to soil in a glasshouse pot study resulted in a 65% increase in seed number (637 vs 385, p < 0.05). We observed a 50% increase in seed production in B. rapa in a pilot hydroponic trial using 0.05 µM selenite compared with low-Se controls. In addition, trials of Se and wheat yielded an 8% increase in root growth (as determined by total root length from seeds germinating in vitro) with the addition of 10 µM selenite (p < 0.05). Other studies have demonstrated a growth response to the addition of Se to wheat (Peng et al. 2000) and rice (Liu et al. 2004). However, no yield increases were observed in our field trials that investigated Se biofortification of wheat, presumably due to adequate available soil Se. Grain Se levels measured in these trials ranged from 60 µg kg−1 DW to 15 mg kg−1DW.
The increased seed production in the treatments did not appear to be associated with any difference in chlorophyll content. A similar result was found in a study using wheat (Munns and James 2003).
The finding of higher respiratory activity associated with low doses of Se is supported by a study of young pea (Pisum sativum L.) plants treated with Se (Smrkolj et al. 2006) and an earlier study using roots of maize (Zea mays L.) (Girton 1974). Respiration can occur by the participation of cytochrome oxidase (cytochrome pathway) or alternative oxidase (the alternative pathway). A small increase in the capacity of the alternative pathway was observed in the Se-treated plants: 31 (se 11) v 24 (6) nmol O2 min−1 g−1 FW. This suggests that the increase in the total respiratory pathway is due mostly to an increase in cytochrome pathway capacity, mediated by cytochrome oxidase (COX). Immunoblot analysis of protein extracts from the flowers of the control and Se-treated plants showed an increase in the relative amount of the COX II protein in flowers of Se-treated plants (Fig. 2). This observation would indicate an increase in the amount of COX complex.

Immunoblot analysis of COX protein in tissue extracts of flowers from selenium treated and un-treated plants. Total protein was extracted from 100 mg of tissue and 6.7 mg of protein loaded onto gel for transfer. Lane 1 is the control plant while lane 2 is the +Se plant. The result presented is typical for the six pseudoreplications
Mitochondria are a site of reactive oxygen species (ROS) production in the cell and thus are prone to damage. The antioxidant system exists within the cell, chloroplast and mitochondria to protect against oxidative damage (Moller 2001a). An increase in total respiratory activity in leaves and flowers of the Se-treated plants compared to that in leaves and flowers of control plants suggests that the mitochondrial activity in Se-treated plants is higher. This may be due to protection of the mitochondria in the Se-treated plants from damage by ROS by an upregulation of the cellular antioxidant defence system. The observed increase in respiration would be unlikely to be part of a general oxidative stress response as the Se concentrations were well below toxic levels. An artificial GSH-Px-like enzyme, Se-containing glutathione-S-transferase, was found to protect bovine heart mitochondria from oxidative stress (Yu et al. 2005).
It is well documented that reproductive processes, especially pollen development, are dependent on mitochondrial function for energy generation (Hanson 1991; Huang et al. 1994; Smart et al. 1994; Heiser et al. 1997). This has been shown by increased levels of mitochondrial protein in floral tissues compared to vegetative tissues. Further, extensive studies of cytoplasmic male sterility (CMS), along with antisense and other reverse genetic studies of mitochondrial proteins (Heiser et al. 1997; Gomez-Casati et al. 2002), have shown a strong correlation between mitochondrial function and pollen viability (Moller 2001b and references cited within). Thus it would seem likely that increased mitochondrial activity in the leaves and flowers would be reflected in pollen viability, leading to increased fertile seed set in the Se-treated plants.
Recent reports support this link between oxidative stress and pollen viability. A knockout mutant line of NADPH-dependent thioredoxin reductases in Arabidopsis thaliana showed reduced pollen fitness (Reichheld et al. 2007) and an analysis of ROS metabolism in cotton with CMS, showed higher ROS content and reduced gene expression of ROS scavenging enzymes co-incident with male cell death in the anthers (Jiang et al. 2007).
Although not conclusive, this suggests that the viability of the higher-Se seed was higher than that of the controls. This is less dramatic than the effects of severe Cu deficiency in higher plants, namely no seed production, whether viable or not, due to unviable pollen (Graham 1975).
However, the proposed mechanism discussed above does not explain why the application of adequate Se should promote dry matter partitioning into seeds, but not also into roots. The concentrations of Se (around 0.05 μM) used in this trial were orders of magnitude below those concentrations which could be toxic. Hence the observed increase in seed production would be unlikely to be part of a general oxidative stress response which could include greater allocation of resources to seed production at the expense of other plant tissues. Further studies that measure seed production and antioxidant enzymes in mitochondria and chloroplasts in other plant species supplemented with low doses of Se would be likely to further elucidate the role of Se in higher plants.
Conclusion
This study found that Brassica plants supplied with a theoretically ideal level of Se in the form of sodium selenite increased partitioning of dry matter into seed production, which was associated with increased respiration in leaves and flowers, compared to low-Se controls. The requirement for separate phytotrons for Se-treated and untreated plants created pseudoreplication, which precluded usual statistical analysis of the results of this trial. In addition, there was a high coefficient of variation in the pseudoreplications, despite the use of second-generation seed from a single parent. Hence, results need to be stated with caution. This study strengthens the evidence for a beneficial role for Se in higher plants.
References
Barker SJ, Stummer B, Gao L, Dsipain I, O’Connor PJ, Smith SE (1998) A mutant in Lycopersicon esculentum Mill. with highly reduced VA mycorrhizal colonisation: isolation and preliminary characterization. Plant J 15:791–797. doi:10.1046/j.1365-313X.1998.00252.x
Cartes P, Gianfreda L, Mora ML (2005) Uptake of selenium and its antioxidant activity in ryegrass when applied as selenate and selenite forms. Plant Soil 276:359–367. doi:10.1007/s11104-005-5691-9
Djanaguiraman M, Durga Devi D, Shanker AK, Sheeba JA, Bangarusamy U (2005) Selenium—an antioxidative protectant in soybean during senescence. Plant Soil 272:77–86. doi:10.1007/s11104-004-4039-1
Easwari K, Lalitha K (1994) Subcellular distribution of selenium during uptake and its influence on mitochondrial oxidations in germinating Vigna radiata L. Biol Trace Elem Res 48:141–160. doi:10.1007/BF02789188
Epstein E, Bloom AJ (2005) Mineral nutrition of plants: principles and perspectives. 2nd Ed. Sinauer, Massachusetts, USA
Girton RE (1974) Effects of selenite selenium on respiration in maize roots. Plant Soil 40:119–127. doi:10.1007/BF00011414
Gomez-Casati DF, Busi MV, Gonzalez-Schain N, Mouras A, Zabaleta EJ, Araya A (2002) A mitochondrial dysfunction induces the expression of nuclear-encoded complex I genes in engineered male sterile Arabidopsis thaliana. FEBS Lett 532:70–74. doi:10.1016/S0014-5793(02)03631-1
Graham RD (1975) Male sterility in wheat plants deficient in copper. Nature 254:514–515. doi:10.1038/254514a0
Hanson M (1991) Plant mitochondrial mutations and male sterility. Annu Rev Genet 25:461–486. doi:10.1146/annurev.ge.25.120191.002333
Hartikainen H, Xue T (1999) The promotive effect of selenium on plant growth as triggered by ultraviolet irradiation. J Environ Qual 28:1372–1375
Hartikainen H, Xue T, Piironen V (2000) Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil 225:193–200. doi:10.1023/A:1026512921026
Heiser V, Rasmusson AG, Thieck O, Brennicke A, Grohmann L (1997) Antisense expression of the mitochondrial NADH-binding subunit of complex I in transgenic potato plants affects male fertility. Plant Sci 127:61–69. doi:10.1016/S0168-9452(97)00118-0
Huang J, Struck F, Matzinger DF, Levings CS (1994) Flower-enhanced expression of a nuclear-encoded mitochondrial respiratory chain protein is associated with changes in mitochondrion number. Plant Cell 6:439–448
Hurlbert SH (1984) Pseudoreplication and the design of ecological field experiments. Ecol Monogr 54:187–211. doi:10.2307/1942661
Jiang PD, Zhang XQ, Zhu YG, Zhu W, Xie HY, Wang XD (2007) Metabolism of reactive oxygen species in cotton cytoplasmic male sterility and its restoration. Plant Cell Rep 26:1627–1634. doi:10.1007/s00299-007-0351-6
Lindstrom K (1983) Selenium as a growth factor for plankton algae in laboratory experiments and in some Swedish lakes. Hydrobiologia 101:35–48. doi:10.1007/BF00008655
Liu Q, Wang DJ, Jiang XJ, Cao ZH (2004) Effects of the interactions between selenium and phosphorus on the growth and selenium accumulation in rice (Oryza sativa). Environ Geochem Health 26:325–330. doi:10.1023/B:EGAH.0000039597.75201.57
Moller IM (2001a) Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu Rev Plant Physiol Plant Mol Biol 52:561–591. doi:10.1146/annurev.arplant.52.1.561
Moller IM (2001b) A more general mechanism of cytoplasmic male fertility? Trends Plant Sci 6:560. doi:10.1016/S1360-1385(01)02100-8
Munns R, James RA (2003) Screening methods for salinity tolerance: a case study with tetraploid wheat. Plant Soil 253:201–218. doi:10.1023/A:1024553303144
Peng A, Xu Y, Lu JH, Wang ZJ (2000) Study on the dose-effect relationship of selenite with the growth of wheat. Biol Trace Elem Res 76:175–181. doi:10.1385/BTER:76:2:175
Pilon-Smits EAH, Garifullina GF, Abdel-Ghany A, Kato S-I, Mihara H, Hale KL, Burkhead JL, Esaki N, Kurihara T, Pilon M (2002) Characterisation of a NifS-like chloroplast protein from Arabidopsis. Implications for its role in sulfur and selenium metabolism. Plant Physiol 130:1309–1318. doi:10.1104/pp.102.010280
Price NL, Thompson PA, Harrison PJ (1987) Selenium: an essential element for growth of the coastal marine diatom Thalassiosira pseudonana (Bacillariophyceae). J Phycol 23:1–9. doi:10.1111/j.1529-8817.1987.tb04493.x
Reichheld JP, Khafif M, Riondet C, Droux M, Bonnard G, Meyer Y (2007) Inactivation of thioredoxin reductases reveals a complex interplay between thioredoxin and glutathione pathways in Arabidopsis development. Plant Cell 19:1851–1865. doi:10.1105/tpc.107.050849
Reilly C (1996) Selenium in food and health. Blackie, London
Shrift A (1969) Aspects of selenium metabolism in higher plants. Annu Rev Plant Physiol 20:475–494. doi:10.1146/annurev.pp.20.060169.002355
Smart CJ, Moneger F, Leaver CJ (1994) Cell-specific regulation of gene expression in mitochondria during anther development in sunflower. Plant Cell 6:811–825
Smrkolj P, Germ M, Kreft I, Stibilj V (2006) Respiratory potential and Se compounds in pea (Pisum sativum L.) plants grown from Se-enriched seeds. J Exp Bot 57:3595–3600. doi:10.1093/jxb/erl109
Sreekala M, Santosh TR, Lalitha K (1999) Oxidative stress during selenium deficiency in seedlings of Trigonella foenum-graecum and mitigation by mimosine. 1. Hydroperoxide metabolism. Biol Trace Elem Res 70:193–207. doi:10.1007/BF02783829
Terry N, Zayed AM, de Souza MP, Tarun AS (2000) Selenium in higher plants. Annu Rev Plant Mol Biol 51:401–432. doi:10.1146/annurev.arplant.51.1.401
Turakainen M, Hartikainen H, Seppanen MM (2004) Effects of selenium treatments on potato (Solanum tuberosum L.) growth and concentrations of soluble sugars and starch. J Agric Food Chem 52:5378–5382. doi:10.1021/jf040077x
Xue TL, Hartikainen H (2000) Association of antioxidative enzymes with the synergistic effect of selenium and UV irradiation in enhancing plant growth. Agric Food Sci Finl 9:177–187
Xue TL, Hartikainen H, Piironen V (2001) Antioxidative and growth-promoting effect of selenium on senescing lettuce. Plant Soil 237:55–61. doi:10.1023/A:1013369804867
Yokota A, Shigeoka S, Onishi T, Kitaoka S (1988) Selenium as inducer of glutathione peroxidase in low-CO2-grown Chlamydomonas reinhardtii. Plant Physiol 86:649–651
Yu H, Liu J, Li J, Zang T, Luo G, Shen J (2005) Protection of mitochondrial integrity from oxidative stress by selenium-containing glutathione transferase. Appl Biochem Biotechnol 127:133–142. doi:10.1385/ABAB:127:2:133
Zarcinas BA, Cartwright B, Spouncer LR (1987) Nitric acid digestion and multi-element analysis of plant material by Inductively Coupled Plasma Spectrometry. Commun Soil Sci Plant Anal 18:131–146
Acknowledgements
The authors gratefully acknowledge funding from HarvestPlus (www.HarvestPlus.org), Washington DC, the Grains Research & Development Corporation (Australia) and the Molecular Plant Breeding Co-operative Research Centre, Australia. Analytical support from Waite Analytical Services, The University of Adelaide, South Australia is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Ismail Cakmak.
Rights and permissions
About this article
Cite this article
Lyons, G.H., Genc, Y., Soole, K. et al. Selenium increases seed production in Brassica. Plant Soil 318, 73–80 (2009). https://doi.org/10.1007/s11104-008-9818-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-008-9818-7