
Abstract
Global warming accelerates land surface drying, increasing the incidence of extreme climatic events such as severe droughts with detrimental effects on ecosystem functions and structure. We investigated the effects of an imposed severe drought (24 days) on fully established synthesized grassland communities with three species richness (S) levels (one, three or nine species), grown for 3 years at either ambient air temperatures (unheated) or ambient +3 °C (heated). Since water supply during these 3 years was equal in all treatments, heated communities experienced more frequent, short mild droughts, but it was unknown whether this conferred greater or smaller resistance for facing prolonged droughts. During the 24-day drought period, soil matric potential decreased in a similar way over time in both temperature treatments and was lower at increasing S-levels. Although green vegetation cover was significantly reduced by the drought in both temperature treatments, the decrease was higher in heated than unheated communities indicating a lower resistance of heated communities to the drought. After only 13 days of recovery, green vegetation cover of both temperature treatments approached values similar to those observed before the imposed drought, suggesting similar resilience in both treatments. Above-ground biomass was reduced by elevated temperature, consistently in all S-levels, showing that the drought period did not change the biomass production patterns observed in the treatments before the imposed drought. Our results suggest that, regardless of the continuous exposure to elevated temperatures and associated short mild droughts, heated communities had not developed clear mechanisms to better cope with extended summer droughts.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Global average surface temperatures are projected to increase by 1.8–4.0 °C by 2100 (best estimate of the range of emissions scenarios), as a consequence of the rising atmospheric concentrations of greenhouse gases (IPCC 2007b). Warming accelerates land surface drying and increases the potential incidence and severity of extreme climatic events such as droughts (IPCC 2007b). For increases in global average temperature higher than 1.5–2.5 °C, major changes are projected to affect also the structure and functioning of ecosystems and the ecological interactions between species, with predominantly negative consequences for biodiversity (IPCC 2007a).
Biodiversity plays an important role in the capacity of ecosystems to respond to global changes (Chapin et al. 1997). Specifically, the so-called ‘insurance hypothesis’ proposed that biodiversity positively influences the long-term stability of ecosystems productivity (Tilman and Downing 1994). Among the components of stability is the ability of a community to maintain the ecosystem functions during a stress event (resistance) or to recover rapidly from it (resilience; Loreau et al. 2002). Biodiversity provides a buffer against environmental fluctuations because different species respond differently to these fluctuations, leading to functional compensations among species (Yachi and Loreau 1999; Tilman et al. 2006). In particular, in the presence of an extreme drought, high species richness (S) levels may increase the probability of including in the ecosystem one or more resistant or resilient species, ensuring species persistence and ecosystem functions. High S-levels may increase community resistance to an extreme event also as a consequence of complementary use of resources, which arises from niche differentiation (Fridley 2001). For example, phenological differences may allow species to use resources at different times over the year (Gulmon et al. 1983), while deep-rooted species may have access to water and nutrients not available to shallow-rooted species (Berendse 1982). In case of an extreme drought, greater complementarity of species-rich communities will determine the use of a larger part of the available water and consequently maintain productivity longer as compared to monocultures of the individual species. The presence of a high number of species in a community would also increase the probability of including a single highly productive species (selection effect, Aarssen 1997; Huston 1997) but if these dominant species are either less resistant or have a low regeneration capacity after the stress, community resistance might decline. As a matter of fact, not all the studies support the positive relationship between biodiversity and stability of productivity. For instance, when synthesized grassland at the Swiss site of the BIODEPTH project, with S-level up to 32 species were exposed to a drought period, the absolute loss of biomass was larger at higher biodiversity, showing a decrease rather than an increase in resistance with higher S-levels (Pfisterer and Schmid 2002).
The current study investigates the effects of an imposed severe drought event on grassland communities with different S-levels, grown for a relatively long time (3 years) at either ambient air temperatures (unheated) or ambient temperatures +3 °C (heated). Specifically we investigated to what extent growing under elevated air temperature and increasing S-levels affected changes in the ecosystem resistance to a severe drought event. The communities grown for 3 years at elevated temperatures often experienced drier soil conditions, since the warming treatment was not combined with increasing precipitation (De Boeck et al. 2006). Having lower soil water content to begin with, heated communities might suffer more from an extended drought episode, but on the other hand these communities could also be better acclimated. Furthermore, elevated temperature itself might modify the responses to the imposed drought. The specific questions of this study were: (1) does soil water content decrease faster during the imposed drought in unheated vs. heated communities or in monocultures vs. mixtures? (2) Given the fact that S-level and temperature influence community water-use and biomass production, does the impact of the imposed drought on ecosystem evapotranspiration (ET), green vegetation cover and above-ground biomass vary with these factors? (3) Do communities grown for 3 years in a warmer climate, or with different S-levels, recover equally well from the imposed drought in terms of green vegetation cover and above-ground biomass?
Materials and methods
Site description
The study was performed in an experimental grassland platform established in 2003, containing 144 artificially assembled ecosystems. The platform was located at the Drie Eiken Campus in Wilrijk, University of Antwerp, Belgium (51° 09′ N, 04° 24′ E, 10 m elevation). At the location, the average annual precipitation is 776 mm, evenly distributed throughout the year, with average annual air temperature (T air) of 9.6 °C.
The platform consisted of six sunlit, climate controlled chambers, facing south. Since 2003, half of the chambers were exposed to ambient T air (unheated chambers) while the other half were continuously warmed at 3 °C above fluctuating ambient T air (heated chambers). Each chamber had an interior surface area of 2.25 m2 and the height of the north and the south sides were 1.5 m and 1.2 m, respectively. The chambers were covered on the top with a colorless polycarbonate plate (4 mm thick), and on the sides with polyethylene film (200 μm thick), both UV transparent. The distance between the chambers was maximized to avoid mutual shading and varied from a minimum of 2.2 m between rows of chambers and 3.1 m within a row of chambers. Each chamber had its individual air control group and was connected to a central cooling unit by isolated pipes. Aerators with regulated flow evenly distributed conditioned air providing ventilation inside the chambers. Relative humidity and T air were measured with a humidity-temperature sensor (Siemens, type QFA66, Germany) while photosynthetically active radiation (PAR) was measured with a quantum sensor (SDEC, type JYP1000, France). All the parameters, recorded in each individual chamber as well as outside, were automatically logged every half hour on a computer. From January 2006 to October 2006 the temperature of the unheated chambers was equal to ambient T air + 0.06 ± (SD) 1.23 °C while the temperature of the heated chambers was equal to ambient T air + 3.02 ± (SD) 0.99 °C. Further details regarding the experimental platform can be found in Lemmens et al. (2006) and De Boeck et al. (2007a).
Plant communities
Each of the six chambers contained the same 24 grassland communities in containers (24 cm inner diameter, 60 cm height), filled with sieved soil (76.3% silt, 14.8% clay and 8.74% sand; field capacity 0.39 m3 m−3, pH = 6.45, 1.6% total C, 0.17% total N) collected from an agricultural field in Brabant, central Belgium. Containers were buried into the soil to avoid unrealistic soil temperatures. Plant communities, established in June 2003, were assembled using nine grassland species, with different productivity, temperature and drought resistance. Three S-levels were created and each chamber contained: nine monocultures (S = 1), nine combinations of three species (S = 3), and six combinations of nine species (S = 9, each community with the same species composition but with a different internal arrangement). Species were selected from three functional groups, which were equally represented in each S-level: three grass species (Dactylis glomerata L., Festuca arundinacea Schreb., Lolium perenne L.), three N-fixing dicots (Trifolium repens L., Medicago sativa L., Lotus corniculatus L.), and three non-N-fixing dicots (Bellis perennis L., Rumex acetosa L., Plantago lanceolata L.).
Each community contained 30 individuals planted in a hexagonal grid at 4 cm distance, with interspecific interactions maximized by avoiding clumping. Each combination at S = 3 had one species from each of the functional groups, and the three species were represented by the same number of individuals. Each species in an S = 3 combination was used only once with any other species leading to three possible sets of nine different S = 3 communities. Only one randomly chosen set of the three possible ones was used in the experiment (De Boeck et al. 2007b). Communities with S = 9 had either three or four individuals per species. The internal arrangement of the communities inside each chamber was randomly assigned. No fertilizer was added and weeding was done manually throughout the experiment. Plants were treated regularly with commercially available products to control fungal infections and insect damage.
Water regime before the imposed drought
Water in the containers could drain freely, while drainage pipes installed below the chambers prevented capillary rise of soil water outside the containers. From the establishment of the ecosystems, and until the start of the imposed drought, unheated and heated chambers received the same amount of water through irrigation so that any enhanced water consumption due to the elevated temperature, would lead to drier soil conditions (De Boeck et al. 2006). Water was supplied twice a week. During the first year, the amount of water supplied was calculated weekly from the difference in soil water content (SWC) between inside and outside the unheated chambers, measured with a PR1-probe (Delta-T Devices Ltd., UK; Lemmens et al. 2006). This method was modified from the second year when the amount of water supplied was calculated based on precipitation data from a nearby weather station (Lint, Belgium, 51° 07′ N, 04° 29′ E, elevation 17 m), corrected for the increased ET inside the chambers (see De Boeck et al. 2006 for further details).
Imposed drought period
Grassland ecosystems of both temperature treatments were subjected to an imposed drought by withholding water from the ecosystems for a period of 24 days, from day of the year (DOY) 185 to 208 (the last irrigation was applied on July 3, 2006, DOY 184), until approximately 50% of the plants in the ecosystems showed severe signs of stress (advanced discoloration due to leaf senescence, leaf wilting and dehydration, and desiccation). At the end of the imposed drought, SWC was returned and kept near container capacity in both temperature treatments for the remaining part of the experiment to allow for full plant recovery.
SWC was measured from the start of the imposed drought, and then every 3–4 days until irrigation was restored. Measurements of SWC were taken with a PR2-probe (Delta-T Devices Ltd., UK) utilizing profile probe tubes (554 mm length), installed in 48 communities (each specific species composition per temperature treatment), across six chambers (three unheated and three heated) at the establishment of the ecosystems. For each community measured, the average SWC at four measurement depths (10, 20, 30, and 40 cm) was used to estimate the community ET with the approach used by De Boeck et al. (2006). The amount of ET per time interval (3–4 days) during the imposed drought period was estimated based on the container area, using the difference in SWC (ΔSWC) per pot between two consecutive measurements. Since no water was added during the period and leaching of water from the pots could be excluded due to the relatively low SWC at the beginning of the drought period, the ΔSWC could be considered an accurate estimate of the ecosystem ET.
In order to describe the relationship between the soil matric potential (Ψ m) and SWC, the soil water retention curve (SWRC) was determined. In order to obtain the SWRC, undisturbed soil samples were collected between 5 and 15 cm depth in four different containers with bare soil per each temperature treatment, using the core method with standard steel 100-cm3 Kopecky rings (height of 5 cm, diameter 5.3 cm; Cornelis et al. 2005). The SWRC was determined with pressure chambers between −20 and −1,500 kPa.
Ecosystem green vegetation cover
To evaluate the impact of the imposed drought and the subsequent recovery on total growth, the relative green cover of each community was estimated by means of digital photography using a Coolpix 5700 digital camera (Nikon Corporation, Tokyo, Japan). Images of individual ecosystems in all chambers were taken at midday under diffuse light from a height of 1 m at three different occasions: three days after the last irrigation (July 6, DOY 187), at the end of the imposed drought (July 26, DOY 207), and 13 days after first re-watering (August 9, DOY 221). Raw images (2560 × 1920 pixels) were analyzed with the Matlab 7 Image Processing Toolbox (The MathWorks, Inc., Natick, USA) using a procedure similar to the one described in Marchand et al. (2004). For effective color processing, images were converted to HSI (hue, saturation and intensity) stacks: hue represents the dominant wavelength of a single color in a mixture of light waves; saturation is the amount of white light mixed with light of a given hue; intensity is the brightness (Choi et al. 1995; Tang et al. 2000). A pixel value threshold was used to obtain the number of green pixels from the hue image. Relative green cover was calculated as the ratio of the total number of green pixels over the total number of pixels in the ecosystem pot area.
Above-ground biomass harvest and analysis of overyielding effects
Total above-ground standing biomass was harvested prior to the imposed drought in late spring, from June 12–14 (DOY 163–165), and after the drought in fall, from October 18–25 (DOY 291–298), 2006. At harvest, all plants in all containers were cut 3.5 cm above the soil surface, sorted by species, dried to constant mass (at least three days at 70 °C), and then weighed. Overyielding of mixtures was assessed calculating selection and complementarity effects using the additive partitioning method proposed by Loreau and Hector (2001). This method is based on the comparison of the observed yields of mixtures with the expected production based on the monoculture yields of each species in the mixture, and allows calculating the relative importance of complementarity and selection.
Statistical analysis
Analysis of variance (ANOVA) and repeated measures ANOVA (RM-ANOVA) were performed with SAS (version 9.1, SAS Institute Inc., Cary, NC, USA), using the mixed procedure (Littell et al. 2006). The statistical analysis was performed considering temperature and S-level treatments as fixed factors and chamber nested within temperature interaction [chamber(temp)], as a random factor. Normality distribution of each variable was tested with a Shapiro–Wilk test: relative green vegetation cover was arc-sin transformed while above-ground biomass was square-root transformed. The chamber factor was always included in the analysis and its effect tested and taken into account if significant. All statistical tests were considered significant at P < 0.05. When treatment effects or interactions were significant, means were separated using a posteriori Least-Squares means test, with Bonferroni corrections for multiple comparisons adjustment. Testing whether the mean complementarity or selection effect differed from zero was done with a one-sample t-test.
Due to interspecific competition over the course of the 3 years prior to the current experiment, the number of species before the imposed drought period had decreased in the mixed communities. Average realized S was 1.9 and 2.1 species in S = 3 unheated and heated communities, respectively, while in S = 9 the realized S was 4.7 and 4.2, respectively. We therefore tested if the realized S in S = 3 and S = 9 was different between temperature treatments. The frequency of distribution of the realized S classes in S = 3 and S = 9 was not significantly different in heated and unheated treatments (χ 2 = 0.201, P = 0.904 for S = 3 classes, χ 2 = 4.53, P = 0.209 for S = 9 classes). Since the realized S-levels were similar among temperature treatments, the communities were analyzed as a function of the initial categories (S-levels 1, 3 or 9). Average realized S for temperature treatments combined was 2 and 4.5 species for S = 3 and S = 9, respectively.
Results
Soil water stress and ecosystem evapotranspiration
During the imposed drought period SWC, and therefore soil Ψ m, was significantly lower in the heated treatment (RM-ANOVA, F 1, 4 = 18.74, P = 0.0125 for SWC, and F 1, 4 = 40.17, P = 0.0031 for soil Ψ m) in all the different days of measurement (F 6, 263 = 7.12, P < 0.0001 for soil Ψ m; Figs. 1 and 2). A principle finding of this study is that during the imposed severe drought event, soil Ψ m decreased in a similar way in both temperature treatments (Fig. 2), as indicated by the non-significant interaction between measurement day and temperature for soil Ψ m (F 6, 263 = 0.08, P = 0.99). Since, under simulated warming, soil Ψ m was lower on the day the drought was imposed, the heated communities consequently reached much greater stress levels by the time irrigation was restored.

Temporal course of soil water content (m3 m–3) during the imposed drought of grassland communities with species richness (S) levels 1 (○), 3 ( ) or 9 (▪; average realized S of 2 and 4.5 species for S = 3 and S = 9, respectively) exposed to a ambient temperature (unheated), and b ambient temperature +3 °C (heated). Each symbol represents the mean ± SE of nine plant communities for S = 1 and S = 3 and of six plant communities for S = 9 measured at four soil depths. Day of the year (DOY) 185 = July 4, 2006
) or 9 (▪; average realized S of 2 and 4.5 species for S = 3 and S = 9, respectively) exposed to a ambient temperature (unheated), and b ambient temperature +3 °C (heated). Each symbol represents the mean ± SE of nine plant communities for S = 1 and S = 3 and of six plant communities for S = 9 measured at four soil depths. Day of the year (DOY) 185 = July 4, 2006

Temporal course of soil matric potential (Ψ m) during the imposed drought of grassland communities with species richness (S) levels 1, 3 or 9 (average realized S of 2 and 4.5 species for S = 3 and S = 9, respectively) exposed to ambient temperature (○, unheated), and ambient temperature +3 °C (●, heated). Each symbol represents the mean ± SE of nine plant communities for S = 1 and S = 3 and of six plant communities for S = 9 measured at four soil depths. Day of the year (DOY) 185 = July 4, 2006
Overall, soil Ψ m significantly decreased with increasing S (F 2, 264 = 5.99, P = 0.0029) and the a posteriori analysis indicated that S = 1 was exposed to less water deficit (higher soil Ψ m) than S = 3 and S = 9 (Fig. 2). Differences between S = 3 and S = 9 were no longer significant. However, the analysis of the significant temperature × S interaction (F 2, 264 = 7.89, P = 0.0005) revealed that differences among S-levels were only significant in the unheated treatment (P < 0.0001 and P = 0.122 for unheated and heated treatments, respectively). Estimated ET was lower in heated than unheated only marginally (RM-ANOVA, F 1, 4 = 7.1, P = 0.056, data not shown). During the drought period, as soil progressively dried, ET significantly decreased over time in all communities (F5, 214 = 10.3, P < 0.0001, data not shown). Species richness effects on ET (F2, 215 = 16.6, P < 0.0001) were similar across temperature treatments, with monocultures having the highest ET rates compared to both S = 3 and S = 9 (a posteriori analysis, P < 0.0001) while no further differences were observed between S = 3 and S = 9 (data not shown).Average values of estimated ET during the imposed drought were equal to 1.05 ± 0.07 and 0.47 ± 0.04 L m−2 day−1 in unheated and heated communities, respectively.
Ecosystem green vegetation cover
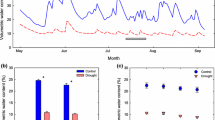
We next explored whether the more intense drought stress that was reached during the imposed drought event in the warmer climate was reflected in a change in the green cover. Overall, the ecosystem green vegetation cover was significantly affected by temperature (RM-ANOVA, F 1, 124 = 20.3, P < 0.0001), species richness level (F 2, 124 = 7.1, P = 0.0012), and date (F 2, 125 = 110.7, P < 0.0001). The a posteriori analysis of the significant temperature × date interaction (F 2, 125 = 15.6, P < 0.0001) indicated that at the start of the imposed drought period green vegetation cover was similar between the two temperature treatments (Fig. 3, DOY 187). Considering the different S-levels, S = 3 and S = 9 had similar green cover, both significantly higher than monocultures (a posteriori analysis, Fig. 3). Green vegetation cover was significantly reduced in both heated and unheated communities by the end of the imposed drought period (Fig. 3, DOY 207). Although the green vegetation cover decreased in both temperature treatments, the relative decrease in cover was higher in heated than unheated (ANOVA, F 1, 4.56 = 21.4, P = 0.0072) and was similar between the S-levels (F 2, 107 = 2.1, P = 0.13; Fig. 3). Surprisingly, after only two weeks of recovery, green vegetation cover significantly increased in both heated and unheated communities, reaching levels not significantly different than those prior to the imposed drought in both treatments (Fig. 3).

Relative green cover (fraction of total area) of grassland communities with species richness (S) levels 1, 3 or 9 (average realized S of 2 and 4.5 species for S = 3 and S = 9, respectively), grown at ambient temperature (unheated treatment, □) and ambient temperature +3 °C (heated treatment, ▪). Means ± SE by S-level and temperature treatment three days after the last irrigation (day of the year, DOY 187), at the end of the imposed drought (DOY 207) and 13 days after re-watering the ecosystems (DOY 221). Percentages on the bars represent the deviation in green cover from the beginning of drought (DOY 187)
Above-ground biomass
Considering that the stress aggravation was similar between temperature treatments, we next investigated if the imposed drought modified biomass patterns. For this purpose we compared total above-ground biomass prior (June) and after (October) the imposed drought. The total above-ground biomass of both harvests was significantly reduced by heating (ANOVA, F 1, 264 = 4.50, P = 0.0347), increased by species richness (F 2, 264 = 27.7, P < 0.0001) but not above S = 3 (a posteriori comparison), and was lower in October than in June (F 1, 264 = 11.6, P = 0.001; Fig. 4a and b). All the interaction terms, including temperature × time, were not significant, indicating that biomass differences observed between heated and unheated communities were similar prior and after the imposed drought (Fig. 4a and b).

Above-ground biomass of grassland communities with species richness (S) levels 1, 3 or 9 (average realized S of 2 and 4.5 species for S = 3 and S = 9, respectively) at (a) ambient temperature (unheated) and (b) ambient temperature +3 °C (heated) harvested in June (prior to the imposed drought, ●) and October (after drought and recovery, △) 2006. Means ± SE of temperature treatments are shown by S-level for June and October
Complementarity and selection effects were then calculated to evaluate whether both could explain the overyielding observed in mixtures and if the severe drought event impacted their relative contribution. In both harvests complementarity and selection were not affected by the temperature treatments (ANOVA, F 1, 166 = 0.26, P = 0.613 for complementarity and F 1, 166 = 3.43, P = 0.066 for selection) and were similar between S = 3 and S = 9 (F 1, 166 = 0.06, P = 0.808 for complementarity, and F 1, 166 = 2.81, P = 0.096 for selection). Overall, the complementarity effect was similar in the two harvests (F 1, 166 = 0.13, P = 0.722) and showed a positive value (0.53 kg m−2, one sample t-test P < 0.0001, Fig. 5).

Partitioning of above-ground biomass overyielding effect of grassland communities with species richness (S) levels 3 and 9 (average realized S of 2 and 4.5 species for S = 3 and S = 9, respectively) compared to average monoculture (S = 1) at ambient temperatures and ambient temperature +3 °C. Means ± SE of temperature treatments are shown combined because not significantly different. Complementarity effects were also not significantly different in June and October and averages are therefore combined
On the other side, the selection effect observed in mixtures in the two harvests was significantly different (F 1, 166 = 5.2, P = 0.024). Although in both harvests there was a positive selection effect, this was significantly reduced after the imposed drought (0.64 kg m−2, P < 0.0001 in June, 0.25 kg m−2, P = 0.003 in October, Fig. 5).
The effect of warming on biomass by species prior and after the imposed drought was calculated in terms of the initial (planted) number of individuals in the communities. Due to the large amount of variation, at the species level no statistical difference in biomass were detected between the two temperature treatments (P = 0.7578) and therefore their combined values are shown in Fig. 6. Species biomass was lower in October than in June 2006 (P = 0.0009) and species differed in total biomass (P < 0.0001), but only at S = 3 and S = 9 (Fig. 6). Overall, different S-levels did not affect species biomass (P = 0.1112), except in dominant species such as M. sativa, R. acetosa and D. glomerata where biomass was positively influenced by the S-level (a posteriori analysis of S-level × species interaction P = 0.0023).

Above-ground biomass of individual species in grassland communities grown at ambient temperature and ambient temperature +3 °C with richness (S) levels 1, 3 or 9 (average realized S of 2 and 4.5 species for S = 3 and S = 9, respectively). Means ± SE a before (June) and b after the imposed drought (October) of six chambers (temperature treatments combined). Species: Bellis perennis (Bp), Dactylis glomerata (Dg), Festuca arundinacea (Fa), Lolium perenne (Lp), Lotus corniculatus (Lc), Medicago sativa (Ms), Plantago lanceolata (Pl), Rumex acetosa (Ra), Trifolium repens (Tr)
Discussion
In this study we investigated the response of heated and unheated communities grown for the 3 years prior to this experiment in the respective climate scenario to a severe drought event. In the 3 years preceding this study, slightly drier soil conditions were consistently observed in the heated communities, as opposed to the unheated. During the period from August 2003 to October 2004, SWC in heated communities was approximately 0.01 m3 m−3 lower than unheated ones, with averages of 0.26 and 0.273 m3 m−3 for heated and unheated communities, respectively (De Boeck et al. 2006). Throughout the imposed drought, heated communities experienced higher water stress compared to unheated ones and drought aggravated similarly in both temperature treatments suggesting that, during the previous 3 years, communities under heated condition had not developed mechanisms to better protect themselves from a severe drought. As expected, the estimated ET during the imposed drought was lower in heated than unheated, confirming previous results on these ecosystems (De Boeck et al. 2006). The lower ET observed under the heated treatments could be explained as the result of several mechanisms. Stomatal regulation is usually one of the first responses to drought stress (Chaves et al. 2002; Flexas et al. 2006). Under the observed level of water stress, stomatal responses would have been triggered, especially in heated communities exposed to higher air vapor pressure deficit due to the elevated temperatures. Also, during the imposed drought plant growth was most likely reduced, particularly in heated communities, causing a lower total leaf area in heated than unheated communities. Under our experimental conditions, the lower leaf area of heated communities was also determined by the higher leaf mortality due to the drought stress as shown in the decrease in green vegetation cover at the end of the drought period.
Considering the differences between S-levels, lower SWC was observed in both S = 3 and S = 9 as compared to S = 1 in both heated and unheated communities. This could be the result of higher below-ground complementarity for water-use, which would allow mixtures to absorb more water from the soil. Our data support the hypothesis that plant diversity enhanced resource acquisition, similar to other studies on synthesized grassland communities exposed to severe or extreme droughts (Pfisterer and Schmid 2002; Van Peer et al. 2004). However, we cannot exclude that complementarity for nutrient uptake could also be partially responsible for the higher water consumption. Under our experimental conditions mixtures yielded higher above-ground biomass, which could partially explain the higher water loss observed. Overall, water stress was higher at increasing S-levels, although these differences proved to be significant only under unheated conditions. Contrary to what suggested by the ‘insurance hypothesis’, under our experimental condition drought stress was not buffered by increasing S-levels. Similarly, Van Peer et al. (2004) reported, in communities exposed to a combination of drought stress and heat wave, higher soil water stress at increasing S-levels due to the complementarity in resource use, which led to greater mortality.
Green vegetation cover was significantly reduced in both temperature treatments after the imposed drought, but its impact was higher in heated than unheated communities. In other words, the relative decrease in green cover was higher in the warming treatment indicating a lower resistance of heated communities to the severe drought. Despite having a more substantial decrease in green cover during the imposed drought, heated communities were able to return to initial values of cover in similar time than unheated ones, suggesting a higher regeneration rate as compared to unheated ones. However, since green vegetation cover reached similar values, the resiliance in the two temperature treatments was not different. Relative decrease in green cover was similar between the S-levels suggesting that ecosystem resistance was not affected by the S-levels, and after approximately two weeks of recovery, all the communities were able to reach similar values of green cover. In a study on model ecosystems of grass species differing in S-levels, individual mortality increased with increasing S-levels after the exposure to a combination of drought and heat wave (Van Peer et al. 2001). Number of tillers per plant increased at increasing S-levels and the total leaf area recovery per living plant (product of tiller number and leaf area per plant) was enhanced by S-levels (Van Peer et al. 2001). In our experiment although resistance to the imposed drought proved to be similar among S-levels, mechanisms such as higher mortality at increasing S-levels may have played a role, but these processes could not be discriminated based on the green vegetation cover observed.
Above-ground biomass was negatively affected by warming prior to and after the imposed drought indicating that the drought event did not change the production pattern observed in the temperature treatments before the imposed drought. This result is in agreement with the similar decrease in SWC and soil Ψm observed in the temperature treatments. Note that above-ground biomass in October included not only the imposed drought period but also the re-growth period after it. During the re-growth period, differences in seasonality between heated and unheated treatments may have acted as a confounding factor, precluding finding difference in biomass due to the drought event. Therefore, the fact that biomass loss was similar in heated and unheated in October should not be interpreted as similar resistance of the two temperature treatments to the imposed drought. After the drought period communities at S = 3 and S = 9 still produced more biomass than monocultures. In a 10-year experiment on grasslands communities with higher S-levels than the ones considered in our study (16 plant species in the highest S-level) the proportional change in biomass production was lower at higher S-levels; plots with the highest diversity were 70% more stable than monocultures (Tilman et al. 2006). However, our results on ecosystems biomass indicated that higher S-levels did not result in higher stability of biomass, in line with similar effects of S-levels on green vegetation cover and aggravation of soil drought.
The shift toward increasing values of root to shoot ratio is also a well-known drought avoidance strategy (e.g. Rodrigues et al. 1995) and total root biomass has been previously correlated with ecosystem stability (Tilman et al. 2006). However, since during the previous below-ground harvests of these grassland communities there were no significant differences in root to shoot ratios between the S-levels nor between the two temperature treatments (De Boeck et al. 2006 and 2007a), this mechanism could only have played a marginal role in the current experiment. Nevertheless, an imposed drought period in semi-natural grasslands induced higher carbon allocation to below-ground components at increasing S-levels, which contributed to maintain ecosystems functions (Kahmen et al. 2005).
After 3 years, the remaining species in the mixtures were mainly the most productive ones in terms of biomass, such as M. sativa and R. acetosa, and were probably responsible for the selection effects observed during the current experiment. Thus, in accordance with other studies (Roscher et al. 2005), both selection and complementarity effects contributed to a positive net biodiversity effect under our experimental conditions. After the imposed drought, the selection effect decreased and complementarity explained most of the overyielding effect. The dominant species in our communities were quite susceptible to drought stress and their decline in biomass was in part responsible for the lower selection effect observed after the drought event. For example, R. acetosa, a species from humid climate (Bruun and Ejrnæs 2000), was previously reported to yield a markedly lower WUE and productivity in heated compare to unheated communities (De Boeck et al. 2006) although under our conditions its biomass decreased after the imposed drought event under both temperature treatments.
In conclusion, our results indicate that communities exposed to warming and short drought events did not show a higher resistance to the imposed severe drought and that increasing richness levels did not enhance resistance either, failing to provide evidence for the ‘insurance hypothesis’. Our results highlight also important mechanisms of ecosystems responses to the combination of long-term exposure to global changes and severe stress events. Our study focused on the single and combined effects of temperature and increasing S-levels on the resistance of grassland communities to an imposed drought period. As concentration of atmospheric CO2 is projected to increase in the future climate together with temperatures, future studies should consider their combined effects. As a matter of fact, elevated CO2 could directly or indirectly affect plant water loss and it will be crucial to understand the combined effect of these climatic factors on ecosystem stability.
References
Aarssen LW (1997) High productivity in grassland ecosystems: affected by species diversity or productive species? Oikos 80:183–184
Berendse F (1982) Competition between plant populations with different rooting depths. III. Field experiments.. Oecologia 53:50–55
Bruun HH, Ejrnæs R (2000) Classification of dry grassland vegetation in Denmark. J Veg Sci 11:585–596
Chapin FS, Walker B, Hobbs RJ, Hooper DU, Lawton JH, Sala OE, Tilman D (1997) Biotic control over the functioning of the ecosystems. Science 277:500–504
Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ricardo CPP (2002) How plants cope with water stress in the field. Photosynthesis and growth. Ann Bot 89:907–916
Choi K, Lee G, Han YJ, Bunn JM (1995) Tomato maturity evaluation using color image analysis. Transactions ASAE 38:171–176
Cornelis WM, Khlosi M, Hartmann R, Van Meirvenne M, De Vos B (2005) Comparison of unimodal analytical expressions for the soil–water retention curve. Soil Sci Soc Am J 69:1902–1911
De Boeck HJ, Lemmens CMHM, Bossuyt H, Malchair S, Carnol M, Merckx R, Nijs I, Ceulemans R (2006) How do climate warming and plant species richness affect water use in experimental grasslands? Plant Soil 288:249–261
De Boeck HJ, Lemmens CMHM, Gielen B, Bossuyt H, Malchair S, Carnol M, Merckx R, Ceulemans R, Nijs I (2007a) Combined effects of climate warming and plant diversity loss on above- and below-ground grassland productivity. Environ Exp Bot 60:95–104
De Boeck HJ, Lemmens CMHM, Vicca S, Van den Berge J, Van Dongen S, Janssens IA, Ceulemans R, Nijs I (2007b) How do climate warming and species richness affect CO2 fluxes in experimental grasslands? New Phytol 175:512–522
Flexas J, Bota J, Galmes J, Medrano H, Ribas-Carbo’ M (2006) Keeping a positive carbon balance under adverse conditions: responses of photosynthesis and respiration to water stress. Physiol Plantarum 127:343–352
Fridley JD (2001) The influence of species diversity on ecosystem productivity: how, where, and why? Oikos 93:514–526
Gulmon SL, Chiariello NR, Mooney HA, Chu CC (1983) Phenology and resource use in three co-occuring grassland annuals. Oecologia 58:33–42
Huston MA (1997) Hidden treatments in ecological experiments: re-evaluating the ecosystem function of biodiversity. Oecologia 110:449–460
IPCC, Intergovernmental Panel on Climate Change (2007a) Climate change 2007: impacts, adaptation and vulnerability. Working Group II. Contribution to the Intergovernmental Panel on Climate Change IV Assessment Report. Summary for policymakers
IPCC, Intergovernmental Panel on Climate Change (2007b) Climate change 2007: the physical science basis. Summary for policymakers. Contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on Climate Change
Kahmen A, Perner J, Buchmann N (2005) Diversity-dependent productivity in semi-natural grasslands following climate perturbations. Funct Ecol 19:594–601
Lemmens CMHM, De Boeck HJ, Gielen B, Bossuyt H, Malchair S, Carnol M, Merckx R, Nijs I, Ceulemans R (2006) End-of-season effects of elevated temperature on ecophysiological processes of grassland species at different species richness levels. Environ Exp Bot 56:245–254
Littell RC, Milliken GA, Stroup WW, Wolfinger RD, Schabenberger O (2006) SAS® for mixed models, 2nd edn. SAS Institute, Cary, NC
Loreau M, Hector A (2001) Partitioning selection and complementarity in biodiversity experiments. Nature 412:72–76
Loreau M, Downing A, Emmerson M, Gonzalez A, Hughes J, Inchausti P, Joshi J, Norberg J, Sala O (2002) A new look at the relationship between diversity and stability. In: Naeem S, Loreau M, Inchausti P (eds) Biodiversity and ecosystem functioning. Oxford University Press, Oxford, pp 79–91
Marchand FL, Nijs I, Heuer M, Mertens S, Kockelbergh F, Pontailler J-Y, Impens I, Beyens L (2004) Climate warming postpones senescence in high-artic tundra. Arctic, Antarctic, Alpine Res 36:390–394
Pfisterer AB, Schmid B (2002) Diversity-dependent production can decrease the stability of ecosystem functioning. Nature 416:84–86
Rodrigues ML, Pacheco CMA, Chaves MM (1995) Soil–plant water relations, root distribution and biomass partitioning in Lupinus albus L. under drought conditions. J Exp Bot 46:947–956
Roscher C, Temperton VM, Scherer-Lorenzen M, Schmitz M, Schumacher J, Schmid B, Buchmann N, Weisser WW, Schulze E-D (2005) Overyielding in experimental grassland communities—irrespective of species pool or spatial scale. Ecol Letters 8:419–429
Tang L, Tian L, Stewart BL (2000) Color image segmentation with genetic algorithm for in field weed sensing. Transactions ASAE 43:1019–1027
Tilman D, Downing JA (1994) Biodiversity and stability in grassland. Nature 367:363–365
Tilman D, Reich PB, Knops JMH (2006) Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 441:629–632
Van Peer L, Nijs I, Bogaert J, Verelst I, Reheul D (2001) Survival, gap formation, and recovery dynamics in grassland ecosytems exposed to heat extremes: the role of species richness. Ecosystems 4:797–806
Van Peer L, Nijs I, Reheul D, De Cauwer B (2004) Species richness and susceptibility to heat and drought extremes in synthesized grassland ecosystems: compositional vs. physiological effects. Funct Ecol 18:769–778
Yachi S, Loreau M (1999) Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proc Nat Acad Sci USA 96:1463–1468
Acknowledgements
This research was funded by the Fund for Scientific Research—Flanders (Belgium) as project “Effects of biodiversity loss and climate warming on carbon sequestration mechanisms in terrestrial ecosystems” (contract G.0434.03N). C. Zavalloni is a beneficiary of a Marie Curie International Reintegration Grant (contract MIRG-CT-2005-031109), which partially financed this study. H.J. De Boeck holds a grant from the Institute for the Promotion of Innovation by Science and Technology in Flanders and B. Gielen is a post-doctoral researcher of the Fund for Scientific Research—Flanders (Belgium). We gratefully thank N. Calluy and F. Kockelbergh for technical assistance, prof. S. Van Dongen for statistical advice, and K. Maguylo for improving the English.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Tibor Kalapos.
Rights and permissions
About this article
Cite this article
Zavalloni, C., Gielen, B., Lemmens, C.M.H.M. et al. Does a warmer climate with frequent mild water shortages protect grassland communities against a prolonged drought?. Plant Soil 308, 119–130 (2008). https://doi.org/10.1007/s11104-008-9612-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-008-9612-6