
Abstract
Soybean cultivars show significant differences in cadmium (Cd) concentrations in seeds, due primarily to genetics, not environmental factors. We previously suggested that low-Cd cultivars accumulate Cd in their roots and thus prevent its translocation to the rest of the plant. Through grafting experiments, we drew the following conclusions about Cd absorption and translocation: (1) The amount of Cd accumulated in shoots is determined by the Cd accumulation capacity of roots: cultivars with a small capacity to accumulate Cd in roots translocate more Cd and accumulate it in shoots; (2) The Cd concentration in shoots is determined by the Cd accumulation capacity of roots and the shoot productive ability of the scion cultivar; (3) The Cd tolerance of shoots differs among cultivars. Enrei, with a high-Cd accumulation capacity of roots, had a low Cd tolerance of shoots compared with Suzuyutaka and Hatayutaka, with a low Cd accumulation capacity of roots; (4) Cultivars differ in their distribution of Cd to seed; (5) These results show that seed Cd concentration is influenced by the differences among cultivars in ease of translocation of Cd to seed and in Cd accumulation capacity of roots.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Recently, the Codex Alimentarius Commission of the United Nations Food and Agriculture Organization (FAO) and the World Health Organization (WHO) proposed a new international standard for cadmium (Cd) concentrations in foodstuffs. Soybean was dropped from the standards, because it is used mainly for extracting oil and is rarely eaten directly in Western countries (CCFAC 2004).
In Japan, however, soybean is a traditional and major food. The results of a large-scale survey of domestic agricultural products revealed that the Cd concentration of one-sixth of soybeans exceeded 0.2 mg kg−1, the international standard then proposed by the Codex Alimentarius Commission, a much higher proportion than in other field crops (MAFFJ 2002). A domestic standard for soybean Cd concentration is anticipated.
Soybeans showed higher Cd concentrations than cereals in a survey of six crops in the main producing districts in the United States (Wolnik et al. 1983) and in a comparison of crops grown in the same soil (Bingham et al. 1975; MacLean 1976). These results indicate that soybeans absorb Cd more easily than other crops. Differences in Cd concentration have been reported among cultivars of lettuce (John and van Laerhoven 1976; Crews and Davies 1985), tobacco (Wagner and Yeargan 1986), rice (Morishita et al. 1987; Arao and Ae 2003), corn (maize) (Hinesly et al. 1978), and sunflower (Li et al. 1995). We previously showed differences in seed Cd concentration among cultivars of soybeans (Arao et al. 2003). Therefore, to produce soybeans safely, in addition to soil treatments such as the spreading of liming materials, it is necessary to identify and breed low-Cd-absorbing cultivars.
The accumulation of Cd in seed of soybeans may be affected by several physiological processes, including Cd uptake from the soil solution, xylem translocation from root to shoot, and phloem movement into seed during maturation. Previously, we found that soybean cultivars with low seed Cd concentrations had low shoot Cd concentrations and high root Cd concentrations, and proposed that they have a mechanism to prevent the translocation of Cd to the shoots by accumulating it in the roots (Arao et al. 2003). Such a role of roots has already been reported in corn (Florijn and van Beusichem 1993) and tobacco (Wagner et al. 1988). We hypothesized that differences among cultivars in Cd accumulation capacity in roots determine the differences in seed Cd concentration, and tested this in grafting experiments.
Materials and methods
Cultivars
We tested soybean cultivars Suzuyutaka, Hatayutaka, Enrei, and Kanto 100 (formerly En-b0-1-2 and Sakukei 4). The seed Cd concentration decreased in that order. We confirmed that this ranking is not influenced by environmental factors, such as soil type and climate (Arao et al. 2003). The family tree of these four cultivars is shown in Fig. 1. Kanto 100 was derived from a mutant of Enrei, and Hatayutaka from a cross of Suzuyutaka and Enrei.

Origin of soybean cultivars tested in this study (boxed)
Grafting
Seeds were germinated in perlite. Grafting was done at the CV stage (Fehr and Caviness 1977), when the unifoliolate leaves were beginning to expand (10 days after sowing). The stem was cut 2 cm under the cotyledons, and grafts of each combination of cultivars were made (Cardwell and Polson 1972). The graft zone was fixed with a commercial graft clip, and seedlings were held indoors at ambient temperature (16–22°C) without artificial enhancement of light for 10 days to reduce transpiration. Ungrafted plants were grown as controls, but delayed by 2 weeks to allow the grafted plants to catch up. In both of experiments the soybeans were grown in a greenhouse at ambient temperature (18–30°C) under sunlight.
Experiment 1—hydroponics
Successfully grafted seedlings were grown in perlite and watered as needed with the following culture solution (mg l−1): N, 60 (NH4NO3); P, 20 (NaH2PO4·2H2O); K, 60 (K2SO4); Ca, 80 (CaCl2·2H2O); Mg, 40 (MgSO4·7H2O); Fe, 2 (Fe[III] EDTA); and Mn, 1 (MnSO4·7H2O). At stage V5 (Fehr and Caviness 1977), when the 4th trifoliolate leaf had expanded (grafted plants, 45 days after sowing; control plants, 31 days after sowing), seedlings were supplied with culture solution containing 0.1 mg Cd l−1. The solution was continually aerated and the pH was between 6.0 and 6.5. After 1 week’s exposure, plants were sampled (n = 6), washed thoroughly in distilled water, and divided into shoots and roots.
Experiment 2—pot experiment using polluted soil
Three cultivars (Suzuyutaka, Hatayutaka, and Enrei) were used in the second experiment. Four seedlings were transplanted per 3.7-l pot filled with 3 kg of air-dried Cd-polluted soil (paddy field Andosol; 0.1 M HCl-extractable Cd, 3.0 mg kg−1). The soil was polluted from irrigation water that had passed through mines. Seedlings were transplanted at 21 days (grafted) or 7 days (control) after sowing. A basal application of fertilizer supplied 0.6 g of N, 0.3 g of P, 0.3 g of K, and 0.1 g of Mg per pot in the form of ammonium sulfate, superphosphate, potassium chloride, and magnesium sulfate, respectively. Plants were thinned at 2 weeks after transplanting (V5) to 1 per pot (n = 12). Adventitious roots coming out of a graft were removed promptly.
Six plants of each treatment were harvested the shoots (including fallen leaves) at the full seed stage (R6, 116 days after sowing) (Fehr and Caviness 1977), and shoots were divided into seeds + pods and leaves + stems. The other six were grown until full maturity (R8) (Fehr and Caviness 1977), and ripened seeds were sampled.
Cadmium analyses of plant tissues and statistical analysis
Plant samples were dried in an oven at 60°C and ground to a fine powder. The powder (0.5 g) was digested in 10 ml HNO3–HClO4–H2SO4 (5:1:1, v/v) in a 2020 Digester (FOSS, Hillerød, Denmark). The Cd concentration was measured by inductively coupled plasma optical emission spectrometer (ICP-OES, Vista Pro, Varian, Mulgrave, Victoria, Australia).
All statistical analyses were performed using Excel 2002 (Microsoft, Redmond, WA, USA) with the add-in software Statcel v. 2 (Yanai 2004). When one-way analysis of variance (ANOVA) was significant among cultivars or among graft combinations as treatments, we used the Tukey–Kramer test to estimate the significance of differences among samples. To test for interactions between scion and rootstock cultivars, we used two-way ANOVA.
Results
Experiment 1—hydroponics
We conducted the hydroponic experiment to reveal differences in Cd absorption by roots among cultivars and the influence of root Cd absorption on shoot Cd concentration. No deficiency symptoms were observed during the experiment. The results of self-grafted and ungrafted soybeans are shown in Table 1. Only two differences were evident: only self-grafted Kanto 100 had significantly lighter roots than the ungrafted control, and self-grafted Suzuyutaka, Hatayutaka, and Enrei had significantly higher shoot Cd concentrations than the ungrafted controls. Whether self-grafted or ungrafted, Cd concentrations showed the same order. The shoot Cd concentration followed the order Suzuyutaka > Hatayutaka > Enrei ≥ Kanto 100. The root Cd concentration followed the order Kanto 100 > Enrei > Suzuyutaka ≥ Hatayutaka, effectively the opposite to that in shoots. The cultivars showed the same rankings in Cd accumulation, also, in both shoots and roots.
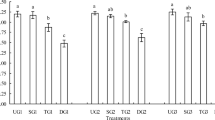
Table 2 shows the influence of scion and rootstock cultivars on dry matter, Cd concentration, and Cd accumulation in cross-grafted cultivars at vegetative stage V8. Neither scion nor rootstock cultivars affected shoot dry weight. Scion cultivars affected root dry weight: Enrei as a scion produced a lower root weight. Rootstock cultivars affected the Cd concentrations of both shoots and roots. The shoot Cd concentration again followed the order Suzuyutaka ≥ Hatayutaka > Enrei > Kanto 100. The root Cd concentration followed the reverse order. The interaction of scion × rootstock also affected the Cd concentrations of both shoots and roots. The Cd concentrations of shoots and roots in all graft combinations are shown in Fig. 2. On both Enrei and Suzuyutaka rootstock, shoot Cd concentration differed between Suzuyutaka and Hatayutaka as scion. When Suzuyutaka was used as a scion on Kanto 100 rootstock, the root Cd concentration was lower than when the other two cultivars were used as scions.

Cd concentrations of grafted soybeans at the vegetative stage (hydroponics). Growth stage was V8: 7th trifoliolate leaves had fully deployed. Error bars indicate standard deviation (n = 6). Means followed by different letters are significantly different (Tukey–Kramer test, p < 0.05)
Rootstock cultivars influenced the Cd accumulation of shoots and roots (Table 2). Shoot Cd accumulation followed the order Suzuyutaka ≥ Hatayutaka > Enrei ≥ Kanto 100, virtually the same order as shoot Cd concentration. Root Cd accumulation followed the order Kanto 100 > Enrei > Suzuyutaka ≥ Hatayutaka, again virtually the same order as root Cd concentration, and the opposite of shoot Cd accumulation.
Experiment 2—pot experiment using polluted soil
To test the hypothesis that differences in the Cd accumulation capacity of roots among cultivars determine differences in the seed Cd concentration among cultivars, we grew grafted plants in polluted soil. At the full seed stage (R6) and full maturity stage (R8), there were no differences between self-grafted and ungrafted soybeans in any parameter (Table 3). Table 4 shows that both scion and rootstock cultivars influenced the dry matter weight, Cd concentration, and Cd accumulation of cross-grafted soybeans at R6 and R8. Table 5 shows the dry matter weight and the Cd accumulation of all graft combinations. The shoot dry matter weight of Hatayutaka as scion was larger than that of the other cultivars as scion (Table 4), and the shoot dry matter weight of Suzuyutaka as rootstock was smaller than that of other cultivars as rootstock.
Rootstock cultivars influenced seed + pod dry weight at R6. That on Suzuyutaka as rootstock was again the smallest (Table 4). Scion cultivars influenced the seed dry weight at R8 in the order Hatayutaka > Suzuyutaka > Enrei. Moreover, the interaction of scion and rootstock cultivars influenced the dry weights of shoots and seeds + pods at R6, and of seeds at R8. Dry weights of Suzuyutaka shoots and seeds + pods at R6 followed the order Hatayutaka ≥ Enrei ≥ Suzuyutaka as rootstock (Table 5). Dry weights of Hatayutaka and Enrei seeds at R8 followed a different order: Enrei ≥ Hatayutaka ≥ Suzuyutaka as rootstock, but there was no difference with Suzuyutaka as scion.
The Cd concentrations of shoots and seeds + pods at R6 and of seeds at R8 were influenced by both scion and rootstock. Hatayutaka as scion had lower Cd concentrations in seeds + pods at R6 and in seeds at R8 than the other cultivars as scions (Table 4). The shoot Cd concentration at R6 showed the same tendency (Table 4). The concentrations as determined by rootstocks all showed the same order: Suzuyutaka > Hatayutaka > Enrei. Moreover, the interaction of scion and rootstock influenced the seed Cd concentration at R8 (Table 4). The seed Cd concentration of all graft combinations at R8 is shown in Fig. 3. When Suzuyutaka was used as rootstock, there was no difference in seed Cd concentration among scion cultivars.

Seed Cd concentration of grafted soybeans at the full maturity stage (pot experiment using polluted soil). Error bars indicate standard deviation (n = 6). Means followed by different letters are significantly different (Tukey–Kramer test, p < 0.05)
Rootstock cultivars influenced the shoot Cd accumulation at R6 in the order Hatayutaka ≥ Suzuyutaka > Enrei (Table 4). Scion cultivars influenced the Cd accumulation in seeds + pods at R6 in the order Enrei > Suzuyutaka > Hatayutaka. Both scion and rootstock cultivars influenced the seed Cd accumulation at R8 in the order Suzuyutaka ≥ Hatayutaka ≥ Enrei. Moreover, the interaction of scion and rootstock influenced the Cd accumulation in shoot and seed + pod at R6 and in seed at R8 (Table 4). When Enrei was used as scion, there was no difference in shoot Cd accumulation at R6 among rootstock cultivars (Table 5). When Suzuyutaka was used as rootstock, there was no difference in seed + pod Cd accumulation between Enrei and Suzuyutaka as scion at R6. There was no difference arising from rootstock in seed Cd accumulation at R8 in Enrei as scion.
Discussion
Influence of roots on Cd translocation to shoots
Comparison of ungrafted and self-grafted soybeans verified that the grafting operation hardly affects dry matter production and the absorption of Cd by soybeans at the vegetative and reproductive stages (Tables 1, 3). At the vegetative stage, the root Cd concentration was influenced by rootstock cultivars in the order Kanto 100 > Enrei > Suzuyutaka ≥ Hatayutaka, both collectively (Table 2) and in each scion separately (Fig. 2). Although the interaction of scion and rootstock cultivars also influenced root Cd concentration (Table 2), the interaction did not limit the influence of the main effect, i.e., rootstock cultivar. Since self-grafted soybean showed the same cultivar ranking of root Cd concentration as above (Table 1), it is clear that there is a difference among cultivars in the Cd concentration of roots, and that this concentration is little influenced by shoots, and does not change with different scions.
Shoot Cd concentration also was influenced by rootstock cultivar, in the order Suzuyutaka ≥ Hatayutaka > Enrei > Kanto 100 (Table 2). The interaction of scion and rootstock also influenced shoot Cd concentration (Table 2). In every scion, the shoot Cd concentration as determined by rootstock cultivar showed virtually the same ranking as the above (Fig. 2), showing that the interaction did not limit the main influence of rootstock cultivar on shoot Cd concentration.
Wagner et al. (1988) found clear differences among tobacco cultivars in shoot Cd concentration and in the influence of rootstock cultivars on shoot Cd concentration. They reported differences among cultivars in the root/shoot concentration ratio, and that cultivars with a higher ratio had a lower shoot Cd concentration. We found clear differences as well: in soybeans at vegetative stage V8, the root/shoot Cd concentration ratio of control plants was 80 in Kanto 100, 58 in Enrei, 22 in Hatayutaka, and 20 in Suzuyutaka (based on data in Table 1). Brown et al. (1958) found that soybean roots controlled iron absorption and translocation to shoots in similar grafting experiments. Similarly, our results show that Cd accumulation in roots reduces Cd translocation and thus the Cd concentration in shoots. Root cells have an important role in cell wall binding of Cd and in limiting its translocation into shoots via the xylem (Wagner 1993; Grant et al. 1998; and references therein). The results of desorption of Cd bound to roots by sequential extraction with BaCl2 and Na2EDTA showed that soybean cultivars with low seed Cd concentration are likely to possess a high affinity and capacity for Cd retention on root cell walls (Ishikawa and Ae 2003). When Suzuyutaka and Hatayutaka, with low Cd accumulation capacity in roots, were used as rootstocks, the shoot Cd concentration became high, independent of the scion cultivar. Conversely, when Kanto 100 and Enrei, with high Cd accumulation capacity in roots, were used as rootstocks, shoot Cd concentration became low. The Cd accumulation in both shoots and roots was also influenced by only rootstock cultivar (Tables 1, 2). This means that the Cd accumulation capacity of roots determined the shoot Cd concentration.
At reproductive stage R6, rootstock cultivar influenced shoot Cd accumulation (excluding Enrei as scion), in almost the same order as at the vegetative stage: Hatayutaka ≥ Suzuyutaka > Enrei. Since the Cd concentration in shoots had the same order as at the vegetative stage (Suzuyutaka > Hatayutaka > Enrei), the Cd accumulation capacity of roots at the vegetative stage did not change at the reproductive stage, and the shoot Cd concentration and accumulation were determined by the Cd accumulation capacity of roots (Tables 4, 5).
However, the influence of scion cultivar appeared in the shoot Cd concentration. Salt et al. (1995) suggested that root-to-shoot translocation of Cd in Brassica juncea was due to transpiration, as abscisic acid (ABA) prevented shoot Cd accumulation. Hart et al. (2006) reported that movement of Cd through the root and into the transpiration stream may be the cause of differential Cd partitioning in two isolines of durum wheat. Since Hatayutaka had more shoot dry matter than the others, we presumed that Hatayutaka as scion accumulated more shoot Cd driven by mass flow due to transpiration. However, the concentration in Hatayutaka as scion was lower than that in the other two cultivars as scion. The shoot Cd accumulation determined by rootstock cultivar might have been diluted by the higher shoot biomass of Hatayutaka (Tables 4, 5). Florijn and van Beusichem (1993) found no correlation between the root/shoot ratio of Cd and transpiration in inbred maize lines, and a similar result in durum wheat was reported (Chan and Hale 2004). We did not investigate the difference in transpiration among soybean cultivars, but transpiration might have some influence on Cd translocation from root to shoot.
Influence of roots on shoot dry matter
The shoot dry matter weight on Suzuyutaka rootstock at R6 was less than that on other rootstocks (Table 4). Some scions grown on Suzuyutaka showed purplish-red veins and leaf stalks and curled leaf tips. These are signs of Cd toxicity (Boggess et al. 1978). The shoot Cd concentration on Suzuyutaka rootstock was higher than that on Hatayutaka and Enrei rootstocks (Table 4), because Suzuyutaka roots have a low-Cd accumulation capacity and thus allow translocation of Cd to shoots. The resultant Cd toxicity reduced the shoot dry matter weight on Suzuyutaka rootstock. If damage had not occurred, the shoot matter dry weight at R6 would have shown the influence of scion cultivar.
Scion cultivars showed different reactions to excess Cd. The growth of Enrei as scion on different rootstocks is shown in Photo 1, and that of Suzuyutaka in Photo 2. The shoot Cd concentration of Enrei on Suzuyutaka (8.4 mg kg−1) was almost the same as that of Suzuyutaka as scion (8.3 mg kg−1) on Suzuyutaka rootstock. Damage was intense: almost all leaves aged and fell early (Photo 1). However, such symptoms were barely apparent in Suzuyutaka as scion (Photo 2). Moreover, the shoot matter dry weight of Enrei on Suzuyutaka was about 2/3 that of Enrei on Enrei. However, there was no difference in shoot matter dry weight of Suzuyutaka as scion between Suzuyutaka and Enrei as rootstocks (Table 5).

Growth of Enrei as scion on different cultivars as rootstock at the full seed stage (pot experiment using polluted soil). Values in parentheses are seed Cd concentrations of Enrei at full maturity (mg kg−1)

Growth of Suzuyutaka as scion grafted on different cultivars as rootstock at the full seed stage (pot experiment using polluted soil). Values in parentheses are seed Cd concentrations of Suzuyutaka at full maturity (mg kg−1)
These results show that the shoot Cd tolerance of Suzuyutaka is higher than that of Enrei and suggest differences in shoot Cd tolerance among cultivars. We think that this difference caused the interactions at the reproductive stage.
Influence of roots and shoots on Cd translocation to seed
The Cd concentrations of seeds + pods at R6 and of seeds at R8 differed considerably among rootstocks, in the order Suzuyutaka > Hatayutaka > Enrei (Table 4). The interaction between scion and rootstock influenced seed Cd concentration at R8, but the influence of rootstock was seen in every graft combination, and the seed Cd concentration showed the same ranking as above (Fig. 3). Moreover, since this ranking was the same as that for the shoot Cd concentrations at V8 (Fig. 2) and R6 (Table 4), it is clear that the Cd concentration of seeds + pods is determined mainly by the Cd accumulation capacity of roots, as is the shoot Cd concentration.
Scion cultivar also influenced the Cd concentration of seeds + pods at R6 and of seeds at R8 (Table 4). Moreover, in self-grafted soybeans, the Cd concentration of seeds + pods at R6 and of seed at R8 followed the order Suzuyutaka > Enrei > Hatayutaka, which is different from that according to rootstock cultivar. The seed + pod/shoot Cd concentration ratios of self-grafted plants at R6, indicating the ease of translocation of Cd in shoots to seed, were Enrei 0.78 > Suzuyutaka 0.64 > Hatayutaka 0.55 (derived from the data in Table 3). Except for plants on Suzuyutaka rootstock, in which symptoms of Cd toxicity appeared in scions, the seed + pod/shoot Cd concentration ratios differed among scion cultivars: Enrei 0.77 > Suzuyutaka 0.67 > Hatayutaka 0.58 (data not shown). Since these rankings and values were almost the same, the ease of translocation of Cd in shoots to seed is characteristic of the scion cultivar. Until the R6 stage at least, we consider that Enrei translocates Cd in shoots to seeds and pods more easily than Hatayutaka can. Harris and Taylor (2001) introduced 109Cd to the phloem of low- and high-Cd accumulating isolines of durum wheat via a leaf flap, and concluded that the ability to remobilize Cd from leaves and stem to maturing grain may control differences in Cd accumulation in grain on a genetic basis. In soybean, the results of grafting experiments showed that Sr, Ca, P, Mg, Mn, and B contents of seeds and other shoot organs were peculiar to scion cultivars (Kleese 1967, 1968; Kleese and Smith 1970; Polson and Smith 1971). Cd translocation to seeds is also peculiar to scion cultivars.
It is clear that the seed Cd concentration is controlled by the rootstock cultivar (Fig. 3). Cross-grafting combinations showed that it was determined mainly by the Cd accumulation capacity of roots, and that it was further influenced by the ease of translocation of Cd to seeds, which was peculiar to each cultivar.
Figure 1 shows that Kanto 100, with the lowest seed Cd concentration, is derived from a mutant of Enrei with high root Cd accumulation capacity, and that Hatayutaka, with low root Cd accumulation capacity, was derived from a cross of Suzuyutaka and Enrei. Suzuyutaka, with a low root Cd accumulation capacity and the highest seed Cd concentration, was derived from Harosoy, which can accumulate even more Cd in seed (Arao et al. 2003). As the seed Cd concentration in durum wheat is based on a single recessive gene (Clarke et al. 1997), we hypothesize that the soybean seed Cd concentration and the Cd accumulation capacity of roots are heritable. Further study should reveal whether this is the case.
References
Arao T, Ae N (2003) Genotypic variations in cadmium levels of rice grains. Soil Sci Plant Nutr 49:473–479
Arao T, Ae N, Sugiyama M, Takahashi M (2003) Genotypic differences in cadmium uptake and distribution in soybeans. Plant Soil 251:247–253
Bingham FT, Page AL, Mahler RJ, Ganje TJ (1975) Growth and cadmium accumulation of plants grown on a soil treated with a cadmium-enriched sewage sludge. J Environ Qual 4:207–211
Boggess SF, Willavize S, Koeppe DE (1978) Differential response of soybean varieties to soil cadmium. Agron J 70:756–760
Brown JC, Holmes RS, Tiffin LO (1958) Iron chlorosis in soybeans as related to the genotype of rootstock. Soil Sci 86:75–82
Cardwell VB, Polson DE (1972) Response of ‘Chippewa 64’ soybean scions to roots of different genotypes. Crop Sci 12:217–219
CCFAC (Codex Alimentarius Commission) (2004) Report of the 36th session of the Codex Committee on Food Additives and Contaminants. Rotterdam, Netherlands. Available online at http://www.codexalimentarius.net/download/report/614/al04_12e.pdf25
Chan DY, Hale BA (2004) Differential accumulation of Cd in durum wheat cultivars: uptake and retranslocation as sources of variation. J Exp Bot 55:2571–2579
Clarke JM, Leisle D, Kopytko GL (1997) Inheritance of cadmium concentration in five durum wheat crosses. Crop Sci 37:1722–1726
Crews HM, Davies BE (1985) Heavy metal uptake from contaminated soils by six varieties of lettuce (Lactuca sativa L.). J Agric Sci 10:5591–5595
Fehr WR, Caviness CE (1977) Stages of soybean development. Special report 80, Iowa State University of Science and Technology, Ames, Iowa, pp 1–11
Florijn PJ, van Beusichem ML (1993) Uptake and distribution of cadmium in maize inbred lines. Plant Soil 150:25–32
Grant CA, Buckley WT, Bailey LD, Selles F (1998) Cadmium accumulation in crops. Can J Plant Sci 78:1–17
Harris NS, Taylor GJ (2001) Remobilization of cadmium in maturing shoots of near isogenic lines of durum wheat that differ in grain cadmium accumulation. J Exp Bot 52:1473–1481
Hart JJ, Welch RM, Norvell WA, Kochian LV (2006) Characterization of cadmium uptake, translocation and storage in near-isogenic lines of durum wheat that differ in grain cadmium concentration. New Phytol 172:261–271
Hinesly TD, Alexander DE, Zeigler EL, Barrett GL (1978) Zinc and Cd accumulation by corn inbreds grown on sludge amended soil. Agron J 70:425–428
Ishikawa S, Ae N (2003) Immobilization of cadmium at root cell wall of soybean cultivars with low seed cadmium. 1st Int Symp Japan–Korea Research Cooperation, Abstract, 89–90. NIAES, Tsukuba
John MK, van Laerhoven CJ (1976) Differential effects of cadmium on lettuce varieties. Environ Pollut 10:163–173
Kleese RA (1967) Relative importance of stem and root in determining genotypic differences in Sr-89 and Ca-45 accumulation in soybean (Glycine max L.). Crop Sci 7:53–55
Kleese RA (1968) Scion control of genotypic differences in Sr and Ca accumulation in soybeans under field conditions. Crop Sci 8:28–129
Kleese RA, Smith LJ (1970) Scion control of genotypic differences in mineral salts accumulation in soybeans (Glycine max L. Merr.) seeds. Ann Bot 34:183–188
Li Y-M, Chaney RL, Schneiter AA, Miller JF (1995) Genotypic variation in kernel cadmium concentration in sunflower germplasm under varying soil conditions. Crop Sci 35:137–141
MacLean AJ (1976) Cadmium in different plant species and its availability in soils as influenced by organic matter and additions of lime, P, Cd and Zn. Can J Soil Sci 56:129–138
MAFFJ (Ministry of Agriculture, Forestry and Fisheries of Japan) (2002) Survey of the cadmium contained in agricultural products. Outline of result. Available online at http://www.maff.go.jp/www/press/cont/20021202press_4.pdf
Morishita T, Fumoto N, Yoshizawa T, Kagawa K (1987) Varietal differences in cadmium levels of rice grains of japonica, indica, javanica, and hybrid varieties produced in the same plot of a field. Soil Sci Plant Nutr 33:629–637
Polson DE, Smith LJ (1971) Nature of scion control of mineral accumulation in soybeans. Agron J 64:381–384
Salt DE, Roger CP, Pickering IJ, Raskin I (1995) Mechanisms of cadmium mobility and accumulation in Indian mustard. Plant Physiol 109:1427–1433
Wagner GJ (1993) Accumulation of cadmium in crop plants and its consequences to human health. Adv Agron 51:173–212
Wagner GJ, Yeargan R (1986) Variation in cadmium accumulation potential and tissue distribution of cadmium in tobacco. Plant Physiol 82:274–279
Wagner GJ, Sutton TG, Yeargan R (1988) Root control of leaf cadmium accumulation in tobacco. Tobacco Sci 32:88–91
Wolnik KA, Fricke FL, Capar SG, Braude GL, Meyer MW, Satzger RD, Bonnin E (1983) Elements in major raw agricultural crops in the United States. 1. Cadmium and lead in lettuce, peanuts, potatoes, soybean, sweet corn, and wheat. J Agric Food Chem 31:1240–1244
Yanai H (2004) Statcel—the useful add-in software forms on Excel, 2nd edn. OMS, Tokyo
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sugiyama, M., Ae, N. & Arao, T. Role of roots in differences in seed cadmium concentration among soybean cultivars—proof by grafting experiment. Plant Soil 295, 1–11 (2007). https://doi.org/10.1007/s11104-007-9252-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-007-9252-2