
Abstract
Darjeeling teas are the highest grown teas in the world and preferred for its flavour, aroma and quality. Apart from the genetic makeup of the plant, earlier reports suggest that insect infestation, particularly jassids and thrips triggers the aroma and flavour formation in Darjeeling tea. The present work encompasses the identification of the genes/transcriptomes responsible for the typical flavour of Darjeeling tea, besides understanding the role of jassids and thrips in particular, in producing the best cup character and quality. The quantitative real time PCR analysis was based on a suppression subtractive hybridisation forward library of B157 (tea clone infested with thrips), providing us transcripts related to aroma and flavour formation. We observed the expression of genes like leucine zipper, ntd, nced, geraniol synthase, raffinose synthase, trehalose synthase, amylase, farnesyl transferase, catalase, methyl transferase, linalool synthase, peroxidases, elicitor responsive proteins, linamarase, nerolidol linalool synthase 2, 12-oxophytodienoate reductase, glucosidase, MYB transcription factor, and alcohol dehydrogenase, highly regulated due to insect infestation, manufacturing stresses and mechanical injury. The first report on gene expression dynamics in thrips infested Darjeeling tea leaves can be extrapolated with increase in volatiles which is responsible for enhancing the quality of Darjeeling tea, specially the flavour and aroma of the infusion. We hope to model these responses in order to understand the molecular changes that occur during Darjeeling tea flavour formation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
India is one of the main tea growers, exporting more than 12% of the world’s tea and with over 521,000 hectares under cultivation. The amount of tea sold as Darjeeling worldwide every year exceeds 40,000 tonnes, while the annual tea production of Darjeeling itself is estimated at only 8,000–11,000 tonnes, including local consumption.
Darjeeling tea has traditionally been prized above all other black teas, especially in the UK and the countries comprising the former British empire. In recent years a high percentage of top quality Darjeeling tea has been bought by Japanese consumers at relatively high prices. When properly brewed it yields thin-bodied, light-colored liquor with a floral aroma and somewhat astringent, tannic characteristics, and spiciness often referred to by tea connoisseurs as ‘muscatel’. Regarded as the “Champagne of Teas,” Darjeeling tea is grown on 100 estates on the foothills of the Himalayas, in over 18,000 hectares at about 7,000 ft. Light and delicate in flavour and aroma, and with undertones of muscatel, Darjeeling is an ideal complement to dinner or afternoon tea. The first “flushes” (pluckings) are thought to produce the best Darjeeling vintage but all crops are of very high quality.
Second flush Darjeeling tea is picked between May and June and produce excellent quality teas that are considered by some to be better than the first flush as they have a fruitier, less astringent flavour than the earlier teas. The leaves are darker brown and contain plenty of silvery tips. It has been reported in literatures that concomitant with the second flush, the population of tea thrips (Scirtothrips dorsalis Hood) and tea jassids (Empoasca flavescens Fabricius) also increases. Whereas the mouth parts of jassids are of piercing and sucking type, this type of mouth parts are developed for drawing fluids from plats by puncturing the epidermis, the mouthparts of thrips consist of rasping and sucking type. As such, in acute infestations the damages are more profound in case of thrips than that of jassids.
It has been reported that emission of volatiles and expression of genes involved in volatile biosynthesis is induced by stresses such as insect attack and wounding in various plant species (Arimura et al. 2005; Bartram et al. 2006; Howe and Schilmiller 2002). Cho et al. (2007) reported chemical profiling and gene expression during the manufacturing process of Taiwan Oolong tea “Oriental Beauty”. The study concentrated on the manufacturing process and the changes during and observed the enhancement of various volatile compounds in the leaves such as Linalool, Geraniol, Methyl salicylate, Linalool oxide I, II, III and IV, Benzyl alcohol, (Z)-3-Hexenol, Hotrienol and 2,6-dimethyl-3,7-octadiene-2,6-diol (DOD).
So far, no reports are there on enhancement of aroma through induced stresses, molecular mechanism during flavour formation and during manufacturing in Darjeeling tea. It was believed that characteristic aroma formation in Darjeeling tea is based on stress-responsive reactions of tea leaves due to infestation by the jassids. In our studies the concept was proven and further, the role of thrips during the infestation was enumerated for the first time. Identification of most of the major floral tea aroma precursors as disaccharide glycosides such as ß-primeverosides in fresh leaves of cultivars led to suggestions that some specific glycosidase should be responsible for the floral tea aroma formation.
The present work encompasses the identification of the genes/transcriptomes responsible for the typical flavour of Darjeeling tea, besides understanding the role of jassids and thrips in particular, in producing the best cup character and quality. The present study provides us information, which can be utilised for production of a new brand of black tea, to sustain the industry by opening newer vistas and diversified products. In addition to the differential expression approach, we also considered the concomitant production of volatile flavory constituents and their relation with respect to the genes identified. We also analysed the gene expression profiles in Darjeeling tea leaves upon mechanical wounding for major genes pertaining to quality, thereby simulating the field conditions of thrips infestation, which may pave the way for a new and unique type of tea in future.
Results and discussion
The experimental setup involved performing the experiment in field condition, where the collection of leaves was done from Darjeeling gardens during the first flush and the second flush of Darjeeling tea. Care was taken so that the samples were subjected to minimum injury post plucking. Sampling was also done at each step of the manufacturing process to determine and monitor the gene expression along with aroma analysis during manufacturing and during mechanical injury experiment. The manufacturing processes involved steps where total RNA and volatile flavour components (VFCs) were extracted from samples collected at each step of the process (Figs. 1, 2a–f).

Flow chart showing stages of the manufacturing process of Darjeeling black tea and the sampling (S2–S9) done at Sungma Tea Estate, Darjeeling, West Bengal (India). Each sample had 3 replicas, and the experiment was repeated twice during 2nd Flush period of Darjeeling tea (May–June, 2007–2008)

a–f Various stages of manufacturing of Darjeeling tea affected by jassids and thrips infestation. a Jassids infested leaves collected separately from Sungma T.E., Darjeeling, India. Inset shows a single leaf with the characteristic pigmentation of the veins. b Thrips infested leaves from the same. Inset shows the characteristic parallel striations on the leaf lamina, as a result of thrips infestation during early leaf morphogenesis from bud. c Withered leaves of second flush during the process of Darjeeling tea manufacturing. d Final Darjeeling tea product ready for the market. e An electron micrograph of an adult thrips (Scirtothrips dorsalis) showing the mouth parts. f The prepared concoction known as the champagne of teas
Analysis of suppression subtractive hybridisation (SSH) and identification of genes
Tea molecular biology is a field where much genomic information on the crop has not been obtained. Therefore genomic information on this health drink can help us determine the factors associated with its beneficial effects on human health. The perennial nature of the crop makes it unsuitable for integrating it into conventional breeding approaches. Hence, unravelling its genomic dataset can be of tremendous application in alternate approaches for application in tea commercialisation and from the crop improvement point of view. We studied the expression of some of the most probable genes (reported in other crops), potential determinant for flavour and aroma in tea and also expression of transcripts obtained upon subtraction of thrips infested B157 leaves from control B157 using SSH approach. The selection of transcripts was done based on available data in other crops and their involvement in defence related functions. Our study involved profiling for monoterpenoid, flavour compounds encoding genes and carbohydrate metabolism genes using quantitative PCR approach.
The present analysis was performed to identify specific transcripts associated with the quality trait in tea. The sequences showed direct or indirect association with different pathways engaged in flavonoid biosynthesis, monoterpenoid and carotenoid biosynthesis, and also few other related to secondary metabolite biosynthetic pathways. These transcripts coded quality related enzymes like alcohol dehydrogenase, phenylalanine lyase, phytoene dehydrogenase, catalase, lipoxygenases, 1-deoxy-d-xylulose 5-phosphate synthase and a few transcription factors. There were few transcripts related to the MEP and Isoprenoid pathway, which are reported to contribute to flavour and aroma in different fruit crops like grapes, strawberry and apple.
More than 1,000 clones from the subtractive library were selected and sequenced. After removal of vector backbone, the sequences were qualified and annotated by BLASTX and BLASTN database searches. Among 621 clones, the deduced amino acid sequence of 472 clones showed significant homology to known functional proteins, the other 149 clones showed uncharacterized proteins (Unknown proteins 37 clones; unnamed proteins, 35 clones; and hypothetical proteins, 77 clones). The sequences of the clones were grouped by DNASIS program. Each consensus sequence of the grouped clones was annotated by BLASTX database search. In addition, a list of selected candidate genes associated with important processes, such as protein metabolism, hormone biosynthesis and signal transduction, transport, and regulation of transcription, and preferentially expressed in the infested leaves of B157, are shown in Table 1.
The percentage of sequence which had a sequence homology with sequences in the public databases was 63% of the total sequence aligned using the BlastX tool at NCBI. Furthermore, we also performed a Blast2GO global analysis by gene ontology (GO) groups. The annotation of the sequences was performed using the InterProScan and BlastX was used to search for sequence homology in the GenBank. A substantial number of sequences pertained to have role in metabolic processes, these metabolic processes were those involved in alcohol catabolism, aromatic amino acid metabolism, carbohydrate metabolism, cellular metabolic processes, macromolecular metabolic processes and others, these sequences were annotated to be involved in regulating function like antioxidant activity (oxidoreductase activity, acting on peroxides as acceptor peroxide), binding activity, catalytic activity, transporter, and molecular function as listed in Table 2. The organellar distribution and abundance of the sequences in enlisted in details in Table 3, these distribution depicted a majority of sequence to be localized in the chloroplast compartment.
The most economically important part of the tea plant is the leaf tissues, so we considered identifying the transcript of tea leaf tissue for the study. Moreover, earlier evidence for Darjeeling quality relative to insect infestation prompted us to consider a hypothesis that insect infestation particularly thrips enhanced the aroma and flavour composition of Darjeeling tea. The subtracted libraries showed overrepresentation of sequences related to flavour and aroma determining pathways. The sequences have been submitted at NCBI with accession numbers HS396832 to HS397309 (sequences in continuity). The raw sequence files were processed to remove vector, adaptor sequences, only sequence larger than 100 bp length and a QV (quality value) of 20 were considered (around 350 sequences). The average length of the sequences was 324 base pairs approximately. A total of 55 contigs and 54 singletons were obtained, which can be together termed as unigenes.
To gain insights into the function of differentially expressed genes, the total transcripts obtained from SSH were categorized into different classes based on their putative roles during infestation. The major functional category corresponded to genes involved in defence, secondary compound synthesis, and cell wall fortification. In addition, another category comprised genes involved in signalling and gene regulation and a significant fraction of genes were involved in detoxification. Genes were also found to play a role in protein synthesis, abiotic stress, photosynthesis or energy metabolism and a major fraction are listed as miscellaneous. Genes, whose function was not ascertained, were categorized as ‘unknown functions’ and considered being Scirtothrips-responsive. This is also to be mentioned that for some of the genes the functional categorization might be arbitrary and there may be some overlaps. Secondary metabolites such as phytoalexin, radical scavengers, and structural barriers serve a vital role in pathogen and insect defence. Several genes potentially involved in secondary metabolite synthesis were identified, including leucoanthocyanidin dioxygenase, cytochrome P450, phenylalanine lyase and methyltransferase. Furthermore, methyltransferase is known to be up-regulated in response to elicitation of insect oral secretions (Giri et al. 2006). Endo-1, 4-β-d-glucanase encoding proteins that function in cell wall fortification were also up-regulated. Genes potentially involved in protection of cells from oxidative stress were up-regulated on insect attack viz. thioredoxin and metallothionein-like protein. Thioredoxins are a group of small proteins functioning in the regulation of redox status of the cell during oxidative stress (Gelhaye et al. 2004). Some of the clones homologous to ATPase were also induced upon infestation. ATPase is reported to be up-regulated in poplar on insect attack and the function assigned to them may be actively transporting a range of ions into or out of the vacuoles or cells to support many biological functions. Moreover, ATPase proteins function as molecular switches in disease signalling pathways. Specific binding and hydrolysis of ATP has been shown for the NBS domains of two tomato CNLs, I2 and Mi. ATP hydrolysis is thought to result in conformational changes that regulate downstream signalling. Ralph et al. (2006) suggested that aphid feeding could induce the expression of H+ ATPase in a resistant plant indicating its role in defence.
It is important to mention that the formation of the flavour and aroma imparting short chain aldehydes and alcohols occur through the action of lipases, hydrogen peroxide lyases and cleavage enzymes on lipid components. These broken down components are acted upon by alcohol dehydrogenase (Galliard et al. 1977). The up-regulation of the lipases in the library supported the excellent quality observed in clones upon infestation by Jassids and thrips, as more the amount of lipases more will be generation of aromatic alcohols and hence flavour and aroma enhancement. The transcripts considered for quantitative PCR expression assays were found in multiples in the representative libraries after sequencing.
Gene expression variation
Mixed infestation
The gene expression patterns in all clones upon infestation showed an up regulation in majority of genes. Alcohol dehydrogenase showed a significant increase in expression upon infestation in all the clones under study. Other genes which showed a significant increase in expression were trehalose-6-phosphate synthase and amylase. Another gene which showed relatively higher expression in infested leaf samples was leucine zipper 9. Expression of raffinose synthase was found to be higher in all the clones, both in control and infested leaf samples (Fig. 3a).





Heat map and hierarchical cluster analysis showing relative changes in gene expression (after normalization) along with production of Volatile Flavoury Components (VFCs, percentage of total; inset) in various tissue samples and clones of Darjeeling tea. [All values considered are normalized using geNorm and heat map generated using GenePattern. Each sample had 3 replicates and Students’ T test was performed for all data considered]. a Mixed infestation samples of different clones; b Jassids and thrips infested samples as compared to mixed infestation and uninfested samples from Sungma1; c Thrips infested and uninfested samples from the three Darjeeling clones; d Profiles at different stages of manufacturing process of black tea; e Scratched samples (mechanical injury)—upper, lower and both surfaces of leaves as compared to uninjured samples
In this study, the samples had both thrips and jassids infestation (mixed infestation). To study the effect of the above infestations separately, samples were collected separately in order to analyze gene expression and flavour formation due to thrips and due to jassids.
Jassid and thrips infestation
For this study, clone Sungma 1 was considered and sampling was done for mixed infestation, jassids infestation, thrips infestation and a control (uninfested). Genes like NCED, Farnesyl transferase, Catalase, Peroxidase and Caffeine synthase hardly showed any variation in expression between all types of infested or control samples (Fig. 3b). All the other genes showed higher expression in infested samples as against control samples. Trehalose-6-phosphate synthase and alcohol dehydrogenase showed a 300 fold increase in thrips infested samples over the control samples. Moreover a 3 fold increase was also observed in the gene expression values of the above two genes when compared with jassids and mixed infestation samples. Most of the other genes showed higher expression in thrips infested samples when compared with jassids infested and mixed infestation samples.
This result confirmed the unique role of thrips in enhancing flavor (confirmed by GC analysis of VFCs) by expression of genes directly or indirectly related to flavour. But it was not clear why the expression of genes in mixed infested samples was less than both jassids and thrips infested samples. Ideally, it should have had an additive effect, maybe the tea plant is unable to bear two infestations at the same time and the capacity to express genes or to produce flavour is somewhat masked due to mixed infestation.
Comparison between clones as a result of thrips infestation
In this study only thrips infested samples were considered for the three clones, along with uninfested (control) samples. All the genes showed characteristic higher expression in the infested samples as against their controls. When we compared the three infested clones, alcohol dehydrogenase showed a 2-fold higher gene expression in Sungma 1 (Fig. 3c). Trehalose synthase, amylase, linamarase and Leucine Zipper 9 showed a similar trend in Sungma 1. It may be mentioned here that Sungma 1 is supposed to produce a unique Darjeeling quality of tea, going by its export market and popularity.
During manufacturing stages
Trehalose-6-phosphate synthase showed higher activity during the different stages of black tea manufacturing. The highest expression being of the withered leaf samples and slowly the expression decreased as depicted by the normalized values of expression represented by colour codes. The other gene which showed an increase was caffeine synthase during the processing of black tea. Linamarase also showed a steady up regulation of expression throughout the manufacturing process. Almost all the genes showed an upregulation in the unwithered, infested samples of Sungma1. A comparative study was done between the expression of genes in this sample with other infested samples during all the stages of black tea processing and the uninfested samples at different stages of manufacturing (Fig. 3d). Besides, trehalose synthase, linamarase, nerolidol linalool synthase 2, and 12-oxophytodienoic acid synthase showed high expression levels during withering and rolling, the last two genes showing such pattern of expression only during manufacturing. This is supported by the fact that although various types of linalool are produced at high levels in field conditions, but the percentage of linalool goes up only during the manufacturing process (withering and rolling stages) (Fig. 3d). This result also suggests that the volatile profiles do go down during downstream processing of black tea, particularly during drying. If linalool and geraniol are compounds that are important for the typical flavour of Darjeeling tea, perhaps re-engineering of the last two steps of manufacturing may restore some of the important compounds that are being lost.
Heirarchical clustering showed that all the infested samples at the different stages of black tea manufacturing formed a separate group. Within that group, withered infested sample (S2) showed the maximum expression for all the genes considered in this study, and the gene expression reduced progressively with further manufacturing. This may be due to the fact that up to withering, the tea leaves are not mechanical or physically damaged, which is done in the downstream steps like drying. It was also observed that the 1st rolling (S4) and the second rolling (S6) had comparable gene expression for all the genes with respect to S2. Overall, withering and the rolling stages produced the best comparative expression profiles for only the infested samples during black tea manufacturing process.
Mechanical injury
The mechanical injury experiments were performed to study the influence of mechanical stress in comparison with insect infestation (thrips), on the expression of genes related to aroma and flavour. The patterns for gene expression revealed upon analysis with GenePattern, grouped thrips infested and scratched leaf surface (lower side) into a single group, which showed higher normalized expression levels for majority of the genes under study. The upper-side scratched and both-side scratched leaf surface samples showed a relatively lower expression for all the genes and they grouped together. Unscratched, uninfested leaf sample were placed alone in a third group. This hierarchical clustering justifies our hypothesis that thrips (which make characteristic scratch on the leaf lower surface) trigger a set of genes which are related to flavour and aroma formation, as a result of the plants inherent defense strategy. Our finding suggests that apart from this they also switch a cascade of events leading to production of volatile and their emission may cause reduction in subsequent herbivory attack (Fig. 3e). This study also helps us to determine whether the thrips infestation can be simulated by mechanical injury. Although a lot of fine tuning needs to be done so far as simulating such a natural phenomenon, preliminary gene expression data does suggest that in-depth studies need to be taken up in a similar line for producing a unique and new type of Darjeeling tea with added or newer flavour attributes.
Comparison of gene expression and VFCs
In order to compare the variation in specific attributes of VFCs in different samples of our experiment, we considered t-2-hexanal, linalool oxide I, linalool oxide II, linalool, phenyl acetaldehyde and phenyl ethanol. These compounds varied significantly between various samples in all the experiments (Fig. 4). Although the percentage production of linalool in all the tea samples was maximum followed by geraniol, but the maximum variation was observed in 2-phenyl ethanol and phenyl acetaldehyde. We correlate this with the gene expression of alcohol dehydrogenase, which is responsible for conversion of phenyl acetaldehyde to 2-phenylethanol. To draw a correlation between formation of the various components of tea flavour with the few genes that we have been able to identify so far would be far-fetched at this point of the investigation. By application of the aroma extract dilution analysis on the volatile fraction isolated from a black tea infusion (Darjeeling Gold Selection), Schu and Schieberle (2006) revealed, in particular, the previously unknown tea constituent (E,E,Z)-2,4,6-nonatrienal as a key odorant in the infusion and confirmed the important role of linalool and geraniol for the tea aroma. Quantitative measurements indicated that, in particular, geraniol, but also eight further odorants were significantly increased in the infusion as compared to their concentration in the leaves.

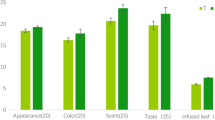
VFC profiles of 3 green leaf samples from Sungma1. In the lower panel, the appearance of 2-phenyl ethanol and phenyl acetic acid is noticeable, besides various other known and unknown components were observed due to thrips infestation. The histogram shows percentage wise variation of the major components between the three samples (error bars are for three replicates per sample)
Other genes and their expression
Other interesting genes preferentially expressed were those potentially involved in phospholipid-mediated signal transduction and transport, such as a protein containing a lipid transfer related domain and related kinases, as well as a MYB transcription factor, genes related to jasmonic acid biosynthesis (allene oxide synthase and SAM-dependent methyltransferase). A number of ESTs corresponding to genes encoding cell wall modification enzymes were found such as two different polygalacturonases, two different xyloglucan-endotransglycosylases, a pectin-methylesterase, a pectate lyase, and a β-galactosidase, in addition to an acidic cellulase that was previously associated with abscission in citrus (Burns et al. 1998). Similarly, genes encoding for cell wall biosynthesis proteins, namely, a UDP-glucose:protein transglucosylase, involved in cellulose biosynthesis, and two genes involved in nucleotide-sugar metabolism, (a UDP-glucose dehydrogenase and a UDP-glucuronic acid decarboxylase), except a cellulose synthase, were also preferentially expressed in B157 infested leaves. A set of 15 ESTs corresponding to two different lipid transfer proteins (LTPs) genes (namely, LTP1, and LTP3) were also preferentially expressed. A set of pathogen-related protein genes including a stress related protein, four different chitinases (10 ESTs), two different glutathione-S-transferases (GSTs) and a putative β-1,3-glucanase was found. ESTs corresponding to five different genes encoding oxidative stress-related enzymes also showed higher gene expression. These were catalase, glutathione dehydrogenase, ascorbate peroxidase, and two independent peroxidases. Interestingly, different genes related to selective ubiquitin mediated protein degradation (i.e. RING-finger protein and an F-box/LRR repeat protein) were also differentially expressed. The expression profiles for SSH derived transcripts showed a higher expression for many of the differentially expressed transcripts in thrips infested leaf samples. The abundance of certain flavour and aroma biosynthesis related transcripts in the thrips infested cDNA libraries supported that there is an enhancement of flavour and aroma metabolism upon infestation by thrips in Darjeeling tea. There was around 59 folds increase in expression of certain transcripts like tryptophan synthase in infested samples than their uninfested counterparts. The abundance of transcripts in this library and the consistent aroma and flavour biosynthesis in these samples suggest that there is a role to be played by these transcripts for this enhanced quality of made tea. The transcripts that were expressed were tubulin specific chaperone, 60s ribosomal protein, dormancy/auxin family related protein, ferredoxin, calmodulin ion binding, phytoene dehydrogenase, translocase of outer membrane, zinc finger, salt tolerance protein, glutathione peroxidases, crt/dre, ATP synthase, PRLi interacting factor, phenylalanine ammonia lyase, Arabidopsis protein targeted to chloroplast protein, plastoquinone oxidoreductase, photosystem II polyprotein, late ethylene and jasmonate biosynthesis, plant synaptotagmin, 3-deoxy-d-arabino-heptulosonate 7-phosphate synthase, MYB transcription factor, lipases, serine/threonine phosphatase 7, cis prenyl transferase, ubiquitin conjugating enzyme, mitogen activated protein kinases, xylulose phosphate synthase, tryptophan synthase. Some were induced profusely upon infestation while others were negatively regulated. The thrips infested samples were clustered in the same group indicating that a similar pattern of gene expression was exhibited upon thrips infestation (Fig. 5).

Heat Map showing gene expression variation between thrips infested and uninfested samples of B157 and AV2. Transcripts considered for this analysis were represented in multiple copies in the SSH library constructed from B157
Analysis of flavour
As a very preliminary work for flavour analysis of infested and uninfested Darjeeling tea, we collected the samples from four different stages of black tea processing namely (1) withered (S2 & S3); (2) 1st rolled (S4 & S5); (3) 2nd rolled (S6 & S7); and (4) fermented black tea (S8 & S9) (Fig. 1). A significant difference in flavour compounds was observed between infested and uninfested samples at all the stages of tea processing. A highly increasing trend of t-2-hexenal, linalool oxides, linalool and geraniol was observed till 2nd rolling and then declined during drying. Changes of α-ionone, β-ionone, and other compounds during tea processing were not remarkable though changes in infested and uninfested samples was prominent.
Preliminary aroma analysis of tea samples prepared in a series at various stages of the manufacturing process clarified that much more volatiles were liberated from the tea samples with the insect-infestation than those without the infestation as the tea processing proceeds. Identification of most of these major floral tea aroma precursors as disaccharide glycosides such as ß-primeverosides in fresh leaves of cultivars led to suggestions that some specific glycosidase should be responsible for the floral tea aroma formation. The present work encompasses the identification of the presence of the precursors in the germplasms studied, the stage at which these precursors are being converted, the enzymes that play a significant role in these conversions and the genes/transcriptomes responsible for the enzymes. The ß-primeverosidase never encounters with the substrates until the cell membrane is damaged. This strongly suggests that the floral aroma formation is partly a self-defense mechanism of tea plants. The understanding of the stage at which the integrity of the cellular structure is changed, the framework collapse leading to direct contact of the enzymes with their substrates would pave the way for applying such knowledge in regulating and retaining the flavour content in processed tea. A monoterpene, 2, 6-dimethyl-3, 7-octadiene-2, 6-diol (DOD) for the first time in the fresh leaves infested by the insect was isolated by Sakata et al. (2005) which is closely related to hotrienol. Hotrienol is considered to be one of the characteristic volatiles and a biochemical marker. These observations indicate that the stresses caused by the insect infestation are very important for the characteristic aroma formation of tea. Stress-responded genes are expressed at this stage and at an early stage of the tea manufacturing during withering and the 1st rolling. There were many kinds of highly induced and moderately induced genes such as ß-amylase, raffinose synthase, mitochondrial protein, senescence-associated protein, jasmonic acid induced protein etc. We have been able to identify upregulated genes due to stress caused by injury which shows similarity with the work reported by Kinoshita et al. (2005). These results indicate that stresses caused by injury induce the expression of various genes and the induced genes result in formation of various enzymes, which are then transported across the membranes or the integrity of the membranes is lost, as was indicated by electron microscopic observations. The enzymes coming in contact with the precursors result in the enhancement of the characteristic aroma products of second flush Darjeeling tea.
Fresh infested tea leaves and infested tea leaves of the fifth turnover were collected and subjected to the processes on and after panning to obtain dried materials. In the final product (black tea) 23 volatile compounds, including 18 alcohols, 2 aldehydes, 3 acids and 1 ester, were identified. Fourteen volatile compounds were detected before the manufacturing process, although the levels were very low, except for linalool and geraniol. Most volatile compound contents increased during the manufacturing process. Aromatic alcohols such as benzyl alcohol and 2-phenylethanol were found to be at the highest level at S6 and dramatically increased, about 400-fold, at S8 as compared to those at S2. The production of monoterpene alcohols such as geraniol and linalool was greatly promoted during manufacture, and linalool oxides and linalool derived compounds also increased significantly. In contrast, diol increased only 1.7 fold and furthermore hotrienol decreased a little during the manufacturing process. Six-carbon (C6) volatiles such as hexanal, hexanol, hexenols and hexenoic acids were hardly found at S2 but increased greatly at S6. Methyl jasmonate was found at S2 but decreased to a trace at S6. Hazarika et al. (1984) studied volatile constituents in orthodox black teas of various clones and flushes in NE India and observed well marked variations in VFC due to differences in weather conditions during the growth periods and manufacturing techniques. Second flush teas contained a higher amount of monoterpenes as linalool, their oxides and geraniol compared to rain flush teas of both low grown and high grown areas. Non-terpenoids such as hexanol, Z-3-hexenol, benzaldehyde and benzyl alcohol were higher in rain flush teas than in second flush teas of only high grown areas but we observed an increase in 2-phenylethanol and phenyl acetaldehyde. Hazarika and Mahanta (1984) revealed the increase or decrease of flavour during different seasons is the consequence of enzymatic reaction sequences controlling the dynamic metabolic system and altering the leaf physiology, which are modified during processing to initiate biochemical reaction sequences resulting in VFC formation. Flavour volatiles, which are synonymous with lipid, fatty acids and lipoxygenase activity, can be a helpful tool to evaluate the quality of clonal cultivars as an objective measurement of organoleptic properties of black tea. Mahanta et al. (1995) observed the distribution of chemical constituents such as total lipid and fatty acid components of fresh tea leaf shoots was maximum during the second flush period to give the superior and distinctive VFC in made tea, compared to pre-second flush, monsoon and end-season flushing period. However, the aroma of tea produced from the China variety in Darjeeling is found to be higher than that of the Assam race (Bhuyan et al. 1991; Horita and Owuor 1987; Mahanta et al. 1993a, b; Venkatakrishna et al. 1976). It would be worthwhile to try the mechanical wounding in Assam type leaves, either before or during the manufacturing process and find out what genes are being upscaled and check the VFC profile. It has been observed by Mahanta et al. (1985), Bhuyan and Mahanta (1989) that the lipid content of tea increases from the first to the third leaf, a fact which might be important in influencing the level of volatile flavour constituents in the various shoot components. Kawakami et al. (1995) found a more complicated aroma pattern in the Darjeeling brewed extract consisting of four linalool oxides, linalool, geraniol, hexanoic acid, benzyl alcohol, 2-phenyl ethanol, trans-geranic acid, (E)-2-hexenoic acid, (Z)-3-hexenoic acid and 2,6-dimethyl-3,7-octadiene-2,6-diol. The Darjeeling SDE extract consists of seven main components which include geraniol, linalool, four linalool oxides and methyl salicylate. The brewed extract of Chan Pin Oolong (red oolong tea), which is similar in aroma to Darjeeling and is made from tea leaves infested with green flies (Empoasca flavescens) in the same way as Darjeeling, contains very high amounts of 2,6-dimethyl-3,7-octadiene-2,6-diol. In continuation of the above, the same working group collected 35 kinds of Darjeeling tea plucked at the tea estates in Darjeeling and estimated high levels linalool oxides I,II,III,IV, linalool, geraniol, benzyl alcohol, 2-phenyl ethanol, hexanoic acid, (E)-2-hexenoic acid, (Z)-3-hexenoic acid, t-geranic acid, dihydroactinidiolide, N-ethylsuccimide, 2,6-dimethyl-3,7-octadiene-2,6-diol and 3,7-dimethyl-1,5,7-octatrien-3-ol. The last two components have apple-Muscat grape like flavour and fresh greenish aroma and contribute to Darjeeling Muscat grape flavour. A review by Drynan et al. (2010) summarising the chemistry of low molecular weight polyphenols isolated and characterised from black tea, including some mentioned above, discusses both structures identified and likely mechanisms of their formation.
Terpene-diol compounds are also components of Muscat grapes, and they have been reported as the thermal degradation in grape juices (Williams et al. 1980). The characteristic VFC are mostly formed by the hydrolytic action of enzymes on cell constituents during leaf maceration (Takeo 1981; Renold et al. 1974). Thus the processing technique largely determines the quantity of flavour components in a made tea. This is indicated by the fact that Assam orthodox teas contain greater amounts of VFC than CTC teas (Takeo and Mahanta 1983). This difference has been attributed to the greater rate of polyphenolic oxidation in CTC teas (Hazarika and Mahanta 1984). On the other hand orthodox Darjeeling teas contain an even greater amount of VFC than orthodox Assam teas, signifying harder processing conditions in the former (Hazarika et al. 1984). The extent of flavour development also depends on the amount of linalool and its derivatives having flower fragrance characteristic aroma of muscatel flavour. There are two major ways in which terpenoid-related volatiles are produced: one involves the enzyme-initiated oxidative breakdown of carotenoids and the other hydrolysis of terpenoid glycosides (Mick and Schreier 1984; Shimizu 1982; Williams et al. 1980; Fischer et al. 1987; Takeo 1981). Yamanishi et al. (1990) investigated the change in flavour constituents during black tea manufacturing and shown that various types of alcohol had increased after withering and that, after fermentation, the aldehyde of (E)-2-hexenal had greatly increased, whereas that of (Z)-3-hexenol had decreased to less than half its level in withered leaves. These results suggest that the glycosidically bound aroma precursors are present in fresh leaves and that glycosidases play the main role in forming black tea aroma (Yano et al. 1990). Characteristic aroma formation in Formosa Oolong tea “Oriental Beauty” is based on stress-responded reactions of tea leaves especially by the tea leafhopper (Jacobiasca formosana) in Taiwan (Kinoshita et al. 2005). Takeo et al. (1985) and Yamanishi (1995) experimental results on analysis of different types of made teas, steamed green tea, perched green tea and oolong tea showed that volatile flavour components are known to contribute to the floral tea aroma and Nishikitani et al. (1996), Sakata (2000), Kobayashi et al. (2001) and Ma et al. (2001) identified the most of these major floral tea aroma precursor as disaccharide glycosides such as ß-primeverosides in fresh leaves of cultivars and these facts strongly suggest that some specific glycosidases should be responsible for the floral tea aroma formation (Ogawa et al. 1995; Pare and Tumlinson 1999; Mizutani et al. 2002). Sakata et al. (2005) isolated and identified aroma precursors of many kinds of the floral tea aroma such as linalool, geraniol, 2-phenylethanol, benzyl alcohol etc. mainly as ß-primeverosides (6-o-ß-d-xylopyranosyl-ß-d-glucopyranosides) and then isolated a diglycosidase, ß-primeverosidase, which can hydrolyze the aroma precursors to release the floral tea aroma. These observations indicate that the stresses caused by the insect infestation are very important for the characteristic aroma formation of the tea.
Conclusion
The basic idea behind the work was to better understand and profile various transcripts and volatile compounds during insect feeding, different stages of manufacturing of Darjeeling tea and mechanical injury. The differential expression profiling was essential and important to selectively determine those transcripts which got regulated due to jassids and thrips infestation. This is the first report on gene expression dynamics in thrips infested Darjeeling tea leaves. The results also provide clues as to which signal pathways are selectively induced by feeding insects. Having deciphered the regulation and expression of the major genes for flavour and aroma in Darjeeling tea, we can implement the knowledge in further processing and exploiting the pathway mechanisms for new nutraceuticals and for producing a unique type of quality in tea.
Materials and method
Plant material
Young two leaves and a bud from Sungma; Tukvar and Ging Tea estates (Darjeeling, West Bengal, India) were collected for gene expression profiling. We took into consideration some of the most popular Darjeeling clones which were believed to be high in flavour and aroma which fetched very high price worldwide. The clones were namely B157, AV2 and Sungma1 (Table 4). AV2 is a fairly hardy hybrid clonal cultivar, doing well at all elevations although leaf gets smaller at higher elevations. The clone has less spreading frame, of erect growing habit, requires closer spacing, leaf small, semi-erect, light green in colour with good tips, shoots medium with high density. It is a high yielder with above average quality having high tip content and good appearance. It is known to be fairly resistant to mites and blister blight, good resistance to drought and moderately susceptible to red spider. B157 is an easy rooter, grows slow initially but starts growing vigorously from second year onwards. B157 is a medium sized, dark green, glossy leafed hybrid clone. It has medium sized frame “shaving brush” type with dense plucking points and many trailing lower branches. Tea manufactured from this clone has very good flavour and above average yield potential of over 1,400 kgs of made tea per hectare. This clone is strongly resistant to red spider, drought, but susceptible to blister blight. It is a very early flusher and keeps flushing till late December. Sungma 1 is a hybrid clone with small dark green leaves. All three clones were evaluated for the typical Darjeeling muscatel flavour quality and are well established and popular as quality clones.
Manufacturing stages and sampling
Tea leaves infested by jassids and thrips and un-infested leaves were plucked in Sungma Tea Estate, Darjeeling. The manufacturing was carried out in the same manner as followed at the Sungma tea factory. For biochemical profiling of volatile compounds, sampling was done at each step of the manufacturing process. In this study we obtained samples for each stage of manufacturing and stored them for both biochemical and gene expression profiling. The schematic representation of the sampling performed at various stages of the manufacturing process is provided in Fig. 1.
Jassids and thrips Infested samples
Jassids and thrips infestation is very prominent in Darjeeling second flush tea leaves. We considered jassids and thrips infested leaves from the gardens in Darjeeling during the second flush (May–June, 2006–2008) to determine the influence of insect infestation on tea quality.
GC analysis
Isolation and analysis of volatile compounds—Volatile compounds from deactivated green leaf and black tea infusions were separated by a column adsorptive method (Mahanta et al. 1985). The concentrated elutes are put together and further concentration is conducted to about 200 μl.
Gas Chromatography–Mass Spectrometry (GC–MS)—Quantification of volatile compounds is carried out on the basis of total ion chromatograms (TIC) in electron impact ionization (EI) mode. The conditions for GC–MS analysis are as described in Hazarika et al. 2005).
Extraction, identification and relative quantification of volatile flavor constituents
The aroma concentrate was prepared from 100 g of black tea by steam distillation under reduced pressure for 45 min using the method reported earlier (Takeo and Mahanta 1983). The concentrate thus obtained was analyzed in Varian GC model CP 3800 equipped with flame ionization detector (FID) and 60 m × 0.25 mm CP-wax 52CB column. Both the injector and detector temperatures were kept at 250°C. The column oven temperature was maintained from 60 to 200°C at the rate of 2°C/min and total run time was 105 min. More than 50 peaks were obtained in the GC profile. Out of these peaks, depending on the availability of standards the individual components were identified by comparing with authentic compounds from Sigma Chemicals. The amount of each component was determined and expressed as the ratio of each peak area to that of internal standard and percentage was determined based on the peak area in the chromatogram (Hazarika et al. 2005).
Total RNA isolation and quality
Total RNA was isolated from the leaf samples (three replicas pulled together) using RNA aquous kit (Ambion, USA). The good quality of RNA was ensured by running a denaturing agarose gel. The quality was also checked using a Spectrophotometer (Eppendorf, Germany).
mRNA solation
mRNA was isolated from the high quality total RNA using PolyATtract® mRNA isolation systems (Promega, USA) following the protocol mentioned in the user manual. The purity and concentration of eluted mRNA was checked using a spectrophotometer (Eppendorf, Germany).
Subtractive cDNA library construction
A suppression subtractive library was constructed taking B-157 infested (thrips), as tester against B-157 uninfested as the driver. It may be mentioned here that all three clones could have been considered for the SSH library construction as all three produce the best quality Darjeeling muscatel flavour. Thus the selection of the clones in the experiment was based on biochemical properties exhibited by the clones upon analysis and also from the excellent quality of processed tea, besides its easy availability in most of the Darjeeling gardens.
First-strand cDNA synthesis
The first strand synthesis for the samples were carried out in duplicates from 2 μg (3 μl) of Poly A+ RNA. The Poly A+ RNA was added to a sterile 0.5 ml microcentrifuge tube containing 1 μl of cDNA synthesis primer (10 μM) and 1 μl of sterile deionised water. After mixing and brief spin the tube was incubated for 2 min in a thermal cycler (Eppendorf) at 70°C. This was followed by addition of 2 μl 5X first strand buffer, 1 μl dNTP Mix (10 mM each), 1 μl sterile water and 1 μl AMV Reverse Transcriptase (20 units/μl). After mixing and brief spin all the tube were incubated at 42°C for 1.5 h in an air incubator. The reactions were terminated by placing the tubes in ice.
Second-strand cDNA synthesis
The second strand synthesis reaction was performed by adding 48.4 μl sterile water, 16 μl 5X second-strand buffer, 1.6 μl dNTP mix (10 mM) and 4 μl of 20X Second-strand enzyme cocktail to the to the above first strand reaction mixtures and incubated at 16°C for 2 h in water bath (Pharmacia). The reactions were terminated by adding 4 μl of 20x EDTA/Glycogen mix. The reaction mixtures were purified by Phenol:chloroform:isoamylalcohol followed by ammonium acetate precipitation. The pellets were washed, dried and dissolved in 50 μl of sterile water.
Preparation of driver cDNA
The purified second-strand cDNAs of all the four samples were digested in a 50 μl reaction mixture containing 15 units of RsaI enzyme (New England Biolabs, UK) for 1.5 h to produce blunt-ended fragments. The cDNAs were phenol-extracted, ethanol-precipitated and then resuspended in 5.5 μl deionised water.
Preparation of tester cDNA
RsaI-digested ds tester cDNA was prepared as described above for the driver. Digested tester cDNA (1 μl) was diluted in 5 μl of deionised water. The diluted tester cDNA (2 μl) was then ligated overnight to 2 μl of adapter 1 (10 μM) and 2 μl adapter 2R (10 μM) in separate ligation reactions in a total volume of 10 μl at 16°C using 0.5 units of T4 DNA ligase (New England Biolabs, UK) in the buffer supplied by the manufacturer. After ligation, 1 μl of 0.2 M EDTA was added and the sample was heated at 70°C for 5 min to inactivate the ligase and stored at −20°C.
Subtractive hybridization
To obtain differentially expressed cDNAs, two rounds of hybridizations were performed. The purpose of the first round of hybridization was to equalize and to enrich the differentially expressed sequences. The objective of the second round was to produce double-stranded tester molecules with different adaptors on each end: 1.5 μl of RsaI-digested driver ds cDNA was added to each of two tubes containing 1.5 μl of adapter1- and adapter 2R-ligated tester cDNA. The samples were mixed, ethanol precipitated and resuspended in 1 μl of hybridization buffer (50 mM HEPES, pH 8.3; 500 mM NaCl; 0.02 mM EDTA, pH 8.0; 10% [w/v] PEG 8000). The solution was overlaid with mineral oil, the DNA was denatured at 98°C for 1.5 min and then allowed to anneal for 8 h at 68°C. After this first hybridization, the two samples were combined and a fresh portion of heated-denatured driver was added to 1 μl of hybridization buffer. The sample was left to hybridize for an additional 10–12 h at 68°C. The final hybridization was then diluted in 200 μl dilution buffer (20 mM HEPES, pH 6.6; 20 mM NaCl; 0.2 mM EDTA, pH 8.0), heated at 72°C for 7 min and stored at −20°C.
PCR amplification
Two rounds of PCR amplifications were performed for each subtraction. In the first amplification, PCR was suppressed, whereby only differentially expressed sequences can be amplified exponentially. In the second procedure, the background was reduced to enrich the differentially expressed sequences: The primary PCR was conducted in 25 μl containing 1 μl of diluted subtracted cDNA, 1 μl of PCR primer 1 (10 μM) and 23 μl of PCR master mixture prepared using the 50 × Advantage cDNA PCR Kit (Clontech, Palo Alto, CA, USA). The PCR was performed with the following parameters: 75°C for 5 min, 27 cycles at (94°C for 10 s, 68°C for 30 s, 72°C for 1.5 min) and a final extension at 72°C for 5 min. The amplified products were diluted by a factor of 10 in deionised water. The primary PCR-diluted products (1 μl) were then used as templates in secondary PCR for 10 cycles under the same conditions as for the primary PCR, with the exception that PCR primer 1 was replaced by nested PCR primers 1 and 2R. The PCR products were analyzed on a 2.0% Agarose/EtBr gel. All of the primers (PCR primer 1 and nested PCR primers 1-2R) for the PCR were in the kit.
PCR analysis of subtraction efficiency
The efficiency of subtraction was tested by doing a PCR taking equal quantity of subtracted and unsubtracted cDNAs of tester as template. The gene specific primer of housekeeping gene 26S was used to compare its abundance in the subtracted and unsubtracted cDNAs of tester. The equal abundance of transcripts of 26S in control and experimental samples were ensured.
Cloning of subtracted cDNA
The subtracted cDNAs, obtained from the second PCR amplification, were cloned into the pGEM-T Easy vector (Promega) and electroporated into E. coli DH10β cells (Invitrogen, USA). The transformed cells were plated onto LB agar culture plates containing ampicillin (100 μg/ml), IPTG (125 mM), X-Gal (40 μg/ml) and were incubated at 37°C overnight. The transformants were selected based on α-complementation. White colonies were randomly picked up and inoculated in 4 ml LB medium containing ampicillin (0.1 μg/ml) at 220 rpm at 37°C overnight. Next day the cells were harvested and plasmid was isolated by alkaline lysis method. The inserts in the extracted plasmids were verified by digesting with EcoRI.
DNA sequencing and homology search
The inserts in the extracted plasmids were sequenced in GA3130xl Genetic Analyzer (Applied Biosystems) using BigDye Terminator V3.1 cycle sequencing Kit (Applied Biosystems) following the instructions given in the manual. The adaptar and vector sequences were removed from the selected good quality (quality value ≥20 and length ≥100 nt) sequences using Sequence Scanner software (Applied Biosystems) and considered for further analysis. A similarity search of all the ESTs, before contig assembly, was performed using the BLASTX programme at the National Centre for Biotechnology Information (http://blast.ncbi.nlm.nih.gov/Blast.cgi/) at default parameters against non-redundant protein database (nr) database.
Functional annotation of differentially expressed ESTs
EST sequences of all the four libraries were used for contig assembly by using a web interface (http://mobyle.pasteur.fr/cgi-bin/portal.py#forms::cap3) of the CAP3 software at default parameters. The unigenes were individually annotated according to the three main vocabularies of the gene ontology (GO): cellular component, biological process and molecular function. Functional annotation of ESTs was performed using the Blast2GO software V.2.4.8 at default parameters. InterPro Scan was performed to find functional motifs and related GO terms by using the specific tool implemented in the Blast2GO software. Finally, the ‘Augment Annotation by ANNEX’ function was used to refine annotations (http://www.goat.no).
Enrichment analysis was performed using Gossip package integrated into Blast2GO to identify the functional categories that are over- or under-represented in the set of differentially expressed genes between libraries of different stages at default p value (i.e. ≤0.05). This package employs the Fisher’s exact test and corrects for multiple testing. For this analysis, the involved sequences with their annotations were loaded in the application. Blast2GO returns the GO terms under or over represented at the specified p value.
Selection of internal control genes for normalization
For internal standard we had selected four housekeeping genes, viz, 26S rRNA, 18S rRNA, Camellia tubulin and Ribulose-1, 5-bisphosphate carboxylase/oxygenase and tested their variability of expression in different samples under consideration. The gene sequences were retrieved from public database and primer was designed using primer3 (http://frodo.wi.mit.edu/primer3/) online software. The details of accession numbers and designed primers are given in Supplementary Table 1.
Selection of genes for qRT-PCR analysis
The list of EST sequences and their homology with other proteins selected for qRT-PCR analysis and their involvement in stress response can be found in Table 1.
Primer design
All primers were designed using DNASTAR and PRIMER3 primer designing softwares. List of the primers used in the study is presented in Supplementary Table 2. Apart from analyzing gene expression pattern for different genes selected from the SSH library sequences there were some additional gene sequences selected for expression studies which were involved in major pathway like the phenylpropanoid pathway, monoterpenoid biosynthesis pathway and those genes/enzymes involved in carbohydrate metabolism. This group of genes were believed to contribute to tea quality in concert.
Standardization of PCR parameters and product size verification
The PCR parameters for all the housekeeping genes and the selected drought induced genes were standardized before going into the qRT-PCR analysis in a gradient thermal cycler (Eppendorf Mastercycler). Specific annealing temperatures were found out at which only single amplicon of desired product size with no primer dimers were formed. The formation of specific desired size amplicon for all the genes were ensured by running an agarose gel. These standardised PCR profiles were used during qRT-PCR analysis.
Two step qRT-PCR
1 μg total RNA was reverse transcribed using transcriptor first strand cDNA synthesis kit (Roche, Germany) following the manufacturers protocol. Briefly, 1 μg of total RNA and 1 μl of anchored-oligo(dT)18 primer and required volume of nuclease free water to make the final volume of 13 μl were gently mixed in a 0.2 ml nuclease free PCR tube and incubated at 65°C for 10 min in a thermal cycler (ABI, 2720) to denature the template-primer mixture. The tube was immediately chilled on ice. To the above mix, 4 μl of 5X transcriptor reverse transcriptase reaction buffer, 0.5 μl protector RNase inhibitor (40 U/μl), 2 μl deoxynucleotide mix (10 mM each) and 2 μl transcriptor reverse transcriptase (20 U/μl) were added, mixed, briefly centrifuged and incubated at 50°C for 1 h in a thermal cycler. The reverse transcriptase enzyme was deactivated by heating the tube at 85°C for 5 min. The first strand was stored at −20°C for downstream application.
Quantitative real time PCR (qRT-PCR)
LightCycler 480 SYBR green I Master kit (Cat. No. 04 707 516 001, Roche Diagnostics, Germany) was used to carry out the expression assays using a LightCycler 480 II real time machine (Roche, Germany). The reverse transcribed first strand cDNA was used as template. All the reactions were carried out in a 20 μl volume prepared according to the protocol mentioned in the kit manual. Three technical replicates of each reaction were used to reduce the error rate. The following amplification programme was used: pre incubation at 95°C for 5 min, this was followed by 45 cycles of amplification with 10 s denaturation at 95°C, annealing for 5–20 s (primer dependent) and extension at 72°C for 30 s. This was followed by one cycle of melting curve analysis to check specificity of amplified product. The temperature for melting curve analysis was 95°C for 5 s, 65°C for 1 min and 97°C for continuous acquisition. This was followed by one cycle of final cooling at 40°C.
Data acquisition
Data on the expression levels were obtained in the form of crossing point (CP), the point where the sample’s fluorescence curve turns sharply upward. The data acquisition was done employing the ‘2nd derivative maximum method as computed by the software of LightCycler (Roche Diagnostics) Carousel-based system.
Computation of normalization factor
We have used geNorm (http://medgen.ugent.be/~jvdesomp/genorm/) software to find out the most stable genes among the set of housekeeping genes at three stages of drought. geNorm calculates a gene expression normalization factor for each tissue sample based on the geometric mean of a user defined number of reference genes. It eliminates the most unstable genes from the group of housekeeping genes step by step and end with two most stable reference genes. In the actual process the raw Cp values of all the housekeeping genes are transformed into relative quantities using delta Ct method. These values are than used for computation of normalization factor in geNorm for all the biological replicates.
Relative quantification
In relative quantification assays, the expression of the target gene is expressed as ratio of target-to-reference gene in the same sample. Here the mean CP values (obtained from three technical replicates) of all the target drought induced genes of all the biological replicates (seven biological replicates in our case) under consideration were transformed into relative quantities using delta Ct method. The normalized expression levels of all the drought induced genes in all the biological replicates were calculated by dividing the quantities by respective normalization factor. This normalised expression value of a gene in different biological replicates was compared to know the relative expression level.
Hierarchical clustering
The hierarchical clustering was performed to study the expression profile of genes across different biological replicates to identify groups of samples that share similar expression profiles. This was performed using GenePattern online software (http://genepattern.broadinstitute.org/gp/pages/index.jsf). Clustering was performed using GenePattern and calculated using Pearson correlation and average linkage as agglomeration rule. The different tonalities of yellow and blue represent up-regulation and down-regulation levels respectively.
References
Arimura G, Kost C, Boland W (2005) Herbivore-induced, indirect plant defences. Biochim Biophys Acta 1734:91–111
Bartram S, Jux A, Gleixner G, Boland S (2006) Dynamic pathway allocation in early terpenoid biosynthesis of stress-induced lima bean leaves. Phytochemistry 67:1661–1672
Bhuyan LP, Mahanta PK (1989) Studies on fatty acid composition in tea Camellia sinensis. J Sci Food Agric 46:325–330
Bhuyan LP, Tamuly P, Mahanta PK (1991) Lipid content and fatty acid composition of tea shoot and manufactured tea. J Sci Food Agric 39:1159–1162
Burns JK, Lewandowski DJ, Nairn CJ, Brown GE (1998) Endo-1,4-β-glucanase gene expression and cell wall hydrolase activities during abscission in Valencia orange. Physiol Plant 102:217–225
Cho JY, Mizutani M, Shimizu B, Kinoshita T, Ogura M, Tokoro K, Lin M, Sakata K (2007) Chemical profiling and gene expression profiling during the manufacturing process of Taiwan Oolong tea “oriental beauty”. Biosci Biotechnol Biochem 71(6):1476–1486
Drynan JW, Clifford MN, Obuchowicz J, Kuhnert N (2010) The chemistry of low molecular weight black tea polyphenols. Nat Prod Rep 27:417–462. doi:10.1039/B912523J
Fischer N, Nitz S, Drawert F (1987) Free and bound aroma compounds of green and black tea. Z Lebensm Unters Forsch 185:195–201
Galliard T, Matthew JA, Wright AJ, Fishwick MJ (1977) The enzymatic breakdown of lipids to volatile and non-volatile carbonyl fragments in disrupted tomato fruits. J Sci Food Agric 28:863–868
Gelhaye E, Rouhier N, Gerard J, Jolivet Y, Gualberto J, Navrot N, Ohlsson P, Wingsle G, Hirasawa M, Knaff DB (2004) A specific form of thioredoxin h occurs in plant mitochondria and regulates the alternative oxidase. Proc Natl Acad Sci, USA 101:14545–14550
Giri AP, Wünsche H, Mitra S, Zavala JA, Muck A, Svatoš A, Baldwin IT (2006) Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. VII. Changes in the plant’s proteome. Plant Physiol 142:1621–1641
Hazarika M, Mahanta PK (1984) Compositional changes of chlorophylls and carotenoids during four flushes of tea in North East India. J Sci Food Agric 35:298–303
Hazarika M, Mahanta PK, Takeo T (1984) Studies on some volatile flavour constituents in orthodox black teas of various clones and flushes in North East India. J Sci Food Agric 35:1201–1207
Hazarika M, Bhuyan LP, Pathak SK (2005) Extraction, identification and quantification of volatile flavoury constituents (VFC) in TRA released Darjeeling clones and standardization of process variables in Darjeeling tea with special reference to VFC. In: Proceedings of the fourth workshop of national tea research foundation, tea research: a new perspective, pp 38–49
Horita H, Owuor PO (1987) Comparison and characterization of volatile components of Kenyan clonal black teas and various black teas from other producing areas of the world. Bull Natl Res Inst Veg Ornam Plants Tea 55–65
Howe GA, Schilmiller AL (2002) Oxylipin metabolism in response to stress. Curr Opin Plant Biol 5:230–236
Kawakami M, Ganguly SN, Banerjee J, Kobayashi A (1995) Aroma composition of Oolong tea and black tea by brewed extraction method and characterizing compounds of Darjeeling tea aroma. J Agric Food Chem 43:200–207
Kinoshita T, Cho JY, Mizutani M, Shimizu B, Tsai HT, Chen YL, Lin ML, Sakata K (2005) Gene expression profiling during the fermentation process of Oriental Beauty. In: 2005 international symposium on innovation in tea science and sustainable development in tea industry, pp 541–545
Kobayashi A, Kubota K, Wang D, Yoshimura T (2001) Specificity of glycosidases from tea leaves toward glycosidic tea aroma precursors In: Takeoka G, Guntert M, Engel KH (eds) Aroma active compounds in food. American Chemical Society, Washington, DC
Ma SJ, Mizutani M, Hiratake J, Hayashi K, Yagi K, Watanabe N, Sakata K (2001) Substrate specificity of β- primeverosidase, a key enzyme in aroma formation during oolong tea and black tea manufacturing. Biosci Biotechnol Biochem 65:2719–2729
Mahanta PK, Hazarika M, Takeo T (1985) Flavour volatiles and lipids in various components of tea shoots Camellia sinensis L.O. Kuntze. J Sci Food Agric 36:1130–1132
Mahanta PK, Tamuly P, Bhuyan LP (1993a) Changes of fatty acid contents, lipoxygenase activities and volatiles during black tea manufacture. J Agric Food Chem 41:1677–1683
Mahanta PK, Baruah SK, Baruah HK, Kalita JN (1993b) Changes of polyphenol oxidase and peroxidase activities and pigment composition of some manufactured black teas Camellia sinensis L. J Agric Food Chem 41:272–276
Mahanta PK, Tamuly P, Bhuyan LP (1995) Comparison of lipoxygenase activity and lipid composition in various harvests of Northeastern Indian tea. J Agric Food Chem 43:208–214
Mick W, Schreier P (1984) Additional volatiles of black tea aroma. J Agric Food Chem 32:924–929
Mizutani M, Nakanishi H, Ema J, Ma SJ, Noguchi E, Inohara-Ochiai M, Fukuchi-Mizutani M, Nakao M, Sakata K (2002) Cloning of β-Primeverosidase from tea leaves, a key enzyme in tea aroma formation. Plant Physiol 130:2164–2176
Nishikitani M, Kubota K, Kobayashi A, Sugawara F (1996) Geranyl 6-O-β-l-arabinopyranosyl-β-d-glucopyranoside isolated as an aromaprecursor from leaves of a green tea cultivar. Biosci Biotech Biochem 60:929–931
Ogawa K, Moon JH, Guo W, Yagi A, Watanabe N, Sakata K (1995) A study on tea aroma formation mechanism: alcoholic aroma precursor amounts and glycosidase and activity in parts of the tea plant. Z Naturforsch 50:493–498
Pare PW, Tumlinson JH (1999) Plant volatiles as a defense against insect herbivores. Plant Physiol 121:325–331
Ralph SG, Oddy C, Cooper D (2006) Genomics of hybrid poplar (Populus trichocarpa × deltoides) interacting with forest tent caterpillars (Malacosoma disstria), normalized and full length cDNA libraries, expressed sequence tags, and a cDNA microarray for the study of insect-induced defences in poplar. Mol Ecol 15:1275–1297
Renold W, Naf-Muller R, Keller U, Willhalm B, Ohloff G (1974) An investigation of tea aroma. Part I. New volatile black tea constituents. Helv Chim Acta 57:1301–1308
Sakata K (2000) In: Parliament TH, Ho C, Schieberie P (eds) Caffeinated beverages, health benefits, physiological effects and chemistry, vol ACS symposium series 754. American Chemical Society, Washington DC, pp 327–335
Sakata K, Mizutani M, Ahn YO, Shimizu B (2005) Floral aroma of Oolong tea are results of stress-responded reactions in tea leaves during the tea processing. In: 2005 international symposium on innovation in tea science and sustainable development in tea industry, 11–15 Nov 2005, organized by Tea research institute, Chinese academy of agricultural sciences, China tea science society and Unilever (China) Ltd., Hangzhou, China, pp 607–617
Schu C, Schieberle P (2006) Characterization of thekey aroma compounds in the beverage prepared from Darjeeling Black Tea: quantitative differences between tea leaves and infusion. J Agric Food Chem 54:916–924
Shimizu J (1982) Identification of volatiles in grape musts and characteristic terpenoid constituents. Agric Biol Chem 46:2265–2274
Yamanishi T, Kobayashi A, Silva, JDE, Botheju WS (1990) Aroma formation of Sri Lanka black tea. In: Charalambous G (ed) Flavours and off-flavours 1989. Proceedings of the 6th international flavour conference, Crete, Greece, 5–7 July 1989, Elsevier Science, Amsterdam, The Netherlands, pp 169–177
Takeo T (1981) Production of linalool and geraniol by hydrolytic breakdown of bound forms in disrupted tea shoots. Phytochemistry 20:2145–2147
Takeo T, Mahanta PK (1983) Comparison of black tea aromas of orthodox and CTC teas and black teas made from different varieties. J Sci Food Agric 34:307–310
Takeo T, Tsushida T, Mahanta PK, Tashiro M, Imamura Y (1985) Study on the food chemistry of aroma of oolong tea and black tea. J Rep Tea Study 20:91–180
Venkatakrishna S, Premachandra BR, Cama HR (1976) Comparative study of the effects of processing on the carotenoids composition of Chinese (Thea sinensis) and Assamese (Thea assamica) tea. Agric Biol Chem 40:2367–2371
Williams PJ, Strauss CR, Wilson B (1980) New linalool derivatives in Muscat of Alexandria grapes and wines. Phytochemistry 19:1137–1139
Yamanishi T (1995) Flavour of tea. Food Rev Int 11:477–525
Yano M, Okada K, Kubota K, Kobayashi A (1990) Studies on the precursors of monoterpene alcohols in tea leaves. Agric Biol Chem 54:1023–1028
Acknowledgments
This work was supported by generous funding from Department of Biotechnology, Govt. of India, Tea Board, Govt. of India and CSIR, Govt. of India. Dr. Vibha Malhotra-Sawhney, Scientist, CSIR, Govt. of India and Dr. Anamika Gambhir, Principal Scientist, DBT for constant encouragement and support.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Gohain, B., Borchetia, S., Bhorali, P. et al. Understanding Darjeeling tea flavour on a molecular basis. Plant Mol Biol 78, 577–597 (2012). https://doi.org/10.1007/s11103-012-9887-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-012-9887-0