
Abstract
Kernel texture of wheat is a primary determinant of its technological properties. Soft kernel texture phenotype results when the Puroindoline a and Puroindoline b genes are present and encode the wild-type puroindolines PINA and PINB, respectively, and various mutations in either or both gene(s) result in hard phenotypes. A wealth of information is now available that furthers our understanding regarding the spatial and temporal regulation of expression of Puroindoline genes. Through the use of model membranes and synthetic peptides we also have a clearer understanding of the significance of the cysteine backbone, the tryptophan-rich domain (TRD) and the helicoid tertiary structures of PIN proteins in relation to their membrane-active properties. Many studies suggest individual yet co-operative modes of action of the PIN proteins in determining kernel texture, and significant evidence is accumulating that the proteins have in vivo and in vitro antimicrobial activities, shedding light on the biological roles of this unique ensemble of proteins. The puroindolines are now being explored for grain kernel texture modifications as well as antimicrobial activities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The leading defining characteristic of common ‘bread’ wheat (Triticum aestivum L., AABBDD), in terms of world trade and food technological applications, is its grain hardness, or endosperm/kernel texture. The kernel texture phenotype is associated with variations in a group of ∼13 kDa proteins, comprised of two major proteins, puroindoline a and b (PINA and PINB) (Morris 2002). The two Puroindoline (Pin) genes Pina-D1 and Pinb-D1 (encoding PINA and PINB, respectively), and the gene Grain softness protein-1 (Gsp-1) encoding the third minor component of this protein group, GRAIN SOFTNESS PROTEIN-1 (GSP-1), are all associated with the Hardness (Ha) locus on chromosome 5D in common wheat. However, the homoeologous loci on chromosomes 5A and 5B lack both Pin genes, these having been deleted during the evolution of durum and thus common wheat. In contrast, both Gsp-1 genes are retained in durum wheat and all three in common wheat. The absence of softness-conferring Pin genes is considered responsible for the very hard texture of durums, soft texture being restored in common wheat through contribution of Pina-D1 and Pinb-D1 genes from the soft-textured D genome donor, Ae. tauschii (DD). Soft texture requires both PINA and PINB proteins in a ‘functional’ state, encoded by the wild-type alleles, designated Pina-D1a and Pinb-D1a, and diverse mutations in either or both genes have been associated with a shift to hard texture. The discovery, purification and properties of PIN proteins and the earlier-known alleles of Pin genes have been reviewed in Morris (2002) and further alleles in wheat and alleles in other Triticeae are discussed in the accompanying paper (Bhave and Morris 2007). A much greater understanding has developed in recent years regarding the expression and regulation of Pin genes in different types of wheat, the biochemical properties and interactions of PIN proteins, and their roles in determining kernel texture. PINs have also been shown to exert significant antimicrobial activities and are being considered for developing applications related to these. These areas are discussed in this review.
Regulation of expression of Puroindoline and GSP-1 genes
Temporal regulation of expression
Gautier et al. (1994) first reported the expression of both Pin genes to be up-regulated in developing wheat seed, with transcripts starting to accumulate at 8–12 days after flowering (DAF), a significant increase at 15–18 DAF, a peak at 26–33 DAF, then a rapid decline and disappearance by 40 DAF. Many further studies (Giroux and Morris 1997; Digeon et al. 1999) report similar observations for Pin and/or Gsp-1 genes, with minor variations in the time of onset or peak of expression, often based on the time points studied or techniques used by the investigators; for example, Hogg et al. (2004) noted a steady increase for Pinb transcripts from 7 to 28 DAF and highest levels of Pina during 14–28 DAF, while Amoroso et al. (2004) showed expression of both genes peaking at 19–26 DAF. Transcripts of Pinb and Gsp-1 were noted at 14 DAF (the only time point studied) in wheat and three Aegilops species (Kan et al. 2006). PINA protein has been detected at 20 DAF (the earliest time point examined; Dubreil et al. 1998), increasing to 40 DAF, then plateauing; later studies using enzyme-linked immunosorbent assays (ELISA) could detect PINA protein at 10 DAF and PINB at 15 DAF (Turnbull et al. 2003a). The transcriptome analyses of developing wheat seeds appears to be particularly sensitive in detecting expressions at very early time points, for example, from 3 to 9 DPA (Clarke and Rahman 2005; Drea et al. 2005; Laudencia-Chingcuanco et al. 2007; McIntosh et al. 2007). Laudencia-Chingcuanco et al. (2007) identified Gsp-1 amongst the genes most differentially expressed and involved in grain filling, along with metabolic or defence functions (e.g., storage proteins, alpha-amylase inhibitors) and expression of PINs and defence proteins correlated with dry weight accumulation. Taking the various observations together, it appears that Pin genes are expressed most actively during the middle stages of endosperm development, Gsp-1 genes possibly a little earlier, and transcription then appears to be terminated but all three proteins are present in mature seeds.
Spatial regulation of expression
A number of efforts have focussed on identifying the (sub)cellular location of PIN proteins in order to better understand their roles in endosperm texture. Some have led to unexpected observations. Transcripts of either Pin gene were not found in roots and coleoptiles after seed germination (Gautier et al. 1994; Dubreil et al. 1998), indicating strict seed-specific expression, confirmed by Pin expression in various seed tissues but not in the embryo, leaf, root or shoot (Digeon et al. 1999). Gautier et al. (1994) localised PINA protein to the vacuoles of aleurone cells in developing seeds. Dubreil et al. (1998) found PINA localised to the starchy endosperm and aleurone, around the intracellular protein-rich aleurone grains, in rehydrated mature seeds. This result was obtained using polyclonal antibodies but not monoclonal, suggesting some cross-reactivity of the polyclonals with PINB; thus PINA may be localised only in the starchy endosperm and PINB in this tissue and/or aleurone (as there were no anti-PINB antibodies used). In seeds just preceding dehydration (40 DAF), PINA was found in the protein matrix and around the starch granules, while in the dehydrated mature seed it occurred only in the protein matrix. Digeon et al. (1999) showed Pinb-D1 transcription was restricted to the endosperm, aleurone and pericarp layers in the developing seed. Capparelli et al. (2005) confirmed localisation of both PINs to the aleurone and endosperm in mature seeds and Drea et al. (2005) confirmed expression of Pinb-D1 and Gsp-1 in endosperm and pericarp from very early stages of development (3–9 DPA). The widely accepted roles of PINA and PINB in determining the texture of mature kernels suggest they would be at the surface of starch granules; thus the reason for their localisation in aleurone is unclear. One theory is that PINs may be synthesised in the aleurone but later transported to endosperm (Gautier et al. 1994). Further work is needed on the cellular and intracellular localisation of PINs at different stages of seed development. Generally, these studies are limited by the specificity and availability of antisera.
Relative expression in different types of wheat
An early suggestion was that the quantity of ‘GSP’ (earlier called friabilin) effected the difference between soft and hard wheats. This was later discounted as seeds of both soft and hard wheats were found to contain significant levels of GSP. However, more total GSP was found in soft kernels and a much greater proportion of it co-isolated with starch, compared to hard wheats (Morris 2002). Many studies have since focussed on comparative studies of expression of the Pin genes and quantities of total and starch-bound puroindolines between soft and hard wheats, to gain a better understanding of the biochemical basis of kernel texture. The results do not yet provide a complete picture, but some general trends emerge.
In the case of soft wheats (Pina-D1a/Pinb-D1a), the extent of transcription of Pina-D1a gene and amount of PINA protein are consistently found to exceed those of Pinb-D1 (Gautier et al. 1994; Giroux and Morris 1997; Turnbull et al. 2000; Turnbull et al. 2003a; Amoroso et al. 2004; Hogg et al. 2004; Gazza et al. 2005; Ikeda et al. 2005; Day et al. 2006), although some difference may be attributable to the extraction processes or analytical techniques used (Turnbull et al. 2003a; Gazza et al. 2005).
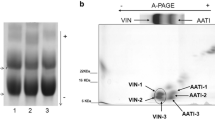
Some interesting observations have been made regarding the relative transcription levels of Pin genes and amounts PIN proteins between soft and hard wheats and the effects of mutations in Pin genes on the quantities of PIN proteins. The transcript levels of Pina-D1 or Pinb-D1 genes have been noted to be identical between soft wheats and Pina-D1a/Pinb-D1b hard wheats (Giroux and Morris 1997), and transcription analyses at different developmental stages of seeds of near-isogenic lines (NILs) in soft (Heron) or hard (Falcon; PINA null) backgrounds showed no significant differences in expression of Pinb-D1 between soft and hard kernel NILs (Clarke and Rahman 2005). Turnbull et al. (2000) reported that amounts of PINA and PINB proteins in soft wheats were marginally to moderately higher than those in Pina-D1a/Pinb-D1b hard wheats, but PINB was drastically reduced in PINA null (Pina-D1b/Pinb-D1a) hard wheats. Corona et al. (2001), using acid-PAGE (A-PAGE), found the quantity of starch-bound PINB was higher in soft compared to hard wheats, but that of PINA was similar between them (except in case of Pina-D1b/Pinb-D1a, i.e. PINA null hard wheats). Further, PINB was present in the meal of PINA-null hard cultivars, indicating that PINA was required for co-operative binding of PINB to starch. The quantity of starch-bound puroindolines was significantly reduced in Pinb-D1b hard wheats compared to most soft wheats; but total kernel puroindoline did not necessarily differ. Capparelli et al. (2003), using flow cytometry and a PINA+PINB-specific monoclonal antibody, found that soft wheats had similar contents of kernel puorindoline, which was generally higher than that in hard wheats, and most of the soft wheat puroindoline was associated with starch. PINA-null (Pina-D1b/Pinb-D1a) hard wheats had very low levels of total puroindoline, and none associated with starch. Pinb-D1b hard wheats had a variable amount of total puroindoline, on occasion exceeding that in soft wheat, and its proportion associated with starch was also highly variable. Further, Pina-D1a mRNA expression was reduced in Pinb-D1b hard wheats compared to soft, suggesting some regulation of its expression and that expression of Pina-D1 regulates association of puroindolines with starch.
The quantitative real-time (RT)-PCR studies of Amoroso et al. (2004) added further interesting results. The transcript levels of Pina-D1a were comparable between soft cultivars and Pinb-D1b hard cultivars (as noted by Giroux and Morris 1997), and transcript levels of Pinb alleles (Pinb-D1a and Pinb-D1b) were similar between soft wheats, Pina-D1b/Pinb-D1a hard wheats, and Pina-D1a/Pinb-D1b hard wheats. However, both Pinb-D1 alleles were generally expressed at a lower level compared to Pina-D1a, and the amount of starch-bound PINA was greater than starch-bound PINB in soft cultivars. Most notably, amount of starch-associated PINB was significantly reduced in PINA-null hard wheats, despite its mRNA levels being similar to soft wheats, suggesting again that PINA is required to stabilise PINB and act in cooperative binding, supporting Corona et al. (2001) and Capparelli et al. (2003). Amoroso et al. (2004) then showed that the total amount of starch-bound puroindolines and the amount of total kernel PINA and PINB had decreased in Pina-D1a/Pinb-D1b hard wheats, but the decrease in PINA was more marked. The results indicate that wild-type PINB might similarly exert some cooperative effect on binding of PINA to starch granules, perhaps by affecting its conformation or affinity for lipids. Assumedly the Gly-to-Ser mutation, which causes alterations to such properties of PINB (Clifton et al. 2007), may be responsible for this effect. The inter-relation was further confirmed by Gazza et al. (2005, 2007), and it was asked whether the wild type PINB synthesized by PINA-null cultivars was preferentially transferred to the protein matrix (rather than to the starch granule surface) (Gazza et al. 2005). Wheat transformation/complementation experiments confirm a major role for PINB and demonstrated that both proteins are needed for kernel softness (discussed below).
No differences were found in the accumulation of Gsp-1 transcripts in soft and hard common wheats or durum wheat, or cDNA sequences from soft and hard wheats (Rahman et al. 1994; Giroux and Morris 1998; Turnbull et al. 2000). However, some recent results suggest interesting possibilities. Clarke and Rahman (2005) reported up-regulation of Gsp-1 transcripts in hard Heron NILs compared to soft Heron NILs. The amount of GSP-1 protein was also reported to be higher in two common wheat cultivars lacking PINA (Ikeda et al. 2005; alleles Pina-D1b and Pina-D1k; Morris and Bhave 2007) and in a durum, compared to common wheats expressing PINA. In light of the reductions seen in PINB in the PINA-null wheats (see above), these data indicate a need for further work on these enigmatic genes.
Promoter sequences in hexaploid wheat
The use of inverse-PCR (IPCR) to clone the flanking sequences, followed by progressive deletions and expression of reporter gene constructs, enabled Digeon et al. (1999) to identify the upstream regulatory regions of Pinb-D1 of common wheat. A number of features were identified including the TATA and CAAT boxes in the −60 to −100 nt region but absence of a GC box, the critical nature of the −380 to −210 region for tissue-specific expression, presence further upstream (up to −560 nt) of specialised sequences such as an E-box (or CANNTG motif; recognised by certain DNA-binding proteins), an ACGT motif recognised by bZIP transcription factor (possibly involved in regulation of seed storage protein genes), a (CA) n element (seed specificity), endosperm box (endosperm specificity) and multiple enhancers. Lillemo et al. (2002) noted that the promoter of both Pin genes had the important prolamin box (endosperm box) but without the GCN4 motif found in storage protein genes, as also noted for Gsp-1 promoters (Turner et al. 1999). The CAAT box of Pina-D1 was found to be inverted and additional regulatory sequences for both Pin genes were reported, i.e., NtBBF1 and dyad repeats (tissue specificity), DOF core recognition sequence (signal transduction and/or tissue specificity) and Amybox1 (response to gibberellin; also found upstream of alpha-amylase genes) (Lillemo et al. 2002). The two promoters shared significant identity (∼78%) in 400-nt upstream sections, decreasing further upstream. The differences might be relevant to differences in the relative extents or tissues of expression of the two genes (see above). Simeone et al. (2006) found the Pina-D1 or Pinb-D1 promoters of a soft wheat and a Pinb-D1b type hard wheat to be identical or very similar, suggesting there was no difference in transcriptional regulation of these genes in these two types of wheat, supporting Amoroso et al. (2004). It will be useful to analyse the promoters of individual Gsp-1 genes in different genomes/ploidy levels due to observations such as the extensive sequence diversity in the gene sequences (Bhave and Morris 2007), a pseudogene in Ae. longissima (Chen et al. 2005), the possible minor effect of only the Gsp-B1 allele on texture (Tranquilli et al. 2002) and silencing of homoeologous genes in wheat (Bottley et al. 2006).
Promoter sequences in diploid progenitors of wheat and wild Triticeae
Amongst the diploid Triticeae taxa, Lillemo et al. (2002) noted that the identity between promoter regions of the hexaploid and diploid Pina genes was much higher than that of their Pinb promoters. The Pina-D1, Pinb-D1, and Gsp-1 promoters of Ae. tauschii shared only 50% identity over −900 nt (Turnbull et al. 2003b), similar to promoters of Pina-D1 and Pinb-D1 genes of hexaploid wheat (Lillemo et al. 2002). The extensive comparative analyses by Simeone et al. (2006) of Pina and Pinb promoters of several diploid Triticeae provided a significant amount of information. Over upstream −280 nt regions, the Pina promoters from all 2n genomes shared ∼93% identity whereas Pinb promoters had a much lower identity (66.5%), confirming the results of Lillemo et al. (2002). Intra-genome group identities were much higher, Pina promoters and Pinb promoters being most conserved amongst the DD (99.6% identity) and AA (99.6%) taxa respectively and both being least conserved amongst SS taxa. Variations were seen in sequences of TATA boxes and the number and sequences of CCAAT boxes, compared to the near-perfect conservation of the prolamin box (except for minor inter-species variations) and one of the (CA) n elements. All Pinb promoters lacked the Amy-box (found in hexaploid Pinb-D1) and P-binding site (also missing in hexaploid Pinb-D1), while all Pina promoters lacked the NtBBf1 motif (also missing in hexaploid Pina-D1). Certain regulatory elements differed between the two genes (e.g., prolamin boxes, E-Box, AACA motif and (GA) n repeats occurred upstream of only Pina genes) and some elements were restricted to only some taxa or genomes. Most of the regulatory sequences occurred in the −300-nt region, except the further upstream Amy-Box, ACGT-Box, P-binding site and a (GA) n repeat in Pina promoters, as in hexaploid wheat. It will be interesting to see whether the variations amongst the Pina and Pinb promoters of the diploid species affect their expression, perhaps as a synergistic factor, considering the coding sequence variations amongst them and the hard kernel texture of only one accession of Ae. sharonensis (Chen et al. 2005). Further, it will be useful to investigate the promoters in accessions of barley and rye considered hard and soft kernels, respectively (Bhave and Morris 2007).
Tertiary structure of puroindolines and biochemical basis of kernel texture and other roles
Tertiary structure, lipid-binding and antimicrobial properties of the PIN-related non-specific lipid-binding proteins
The lipid-binding proteins of the wheat kernel are of interest due to their relevance to lipid functionality in bread-making and brewing, especially their surface activity. They have been investigated as potential factors to improve lipid spreading and stability and thus bread volumes (reviewed in Douliez et al. 2000). The wheat starch lipid-binding proteins are small, cysteine-rich albumins that share structural similarities with proteins called non-specific lipid-transfer proteins (ns-LTPs), which are small (7 or 9 kDa) basic proteins ubiquitous to, but restricted to, the plant world. They function during normal plant growth and development and are involved in inter-membrane lipid trafficking at cellular and extracellular levels and mobilizing monomers of materials that make up the external protective layers (Kader 1996; Garcia-Olmedo et al. 1998; Douliez et al. 2000; Blein et al. 2002). Some may also have technological applications; for example, barley LTP1 is surface-active and aids in the formation of beer foam (Douliez et al. 2000; Perrocheau et al. 2006). Of particular interest is the fact that a number of ns-LTPs exhibit antibacterial, antifungal or antiviral activities, some isoforms being up-regulated under pathogen infections or involved in signalling pathways (Kader 1996; Garcia-Almedo et al. 1998; Blein et al. 2002; Cameron et al. 2006). The ns-LTPs (and other defence-related proteins) are expressed at early stages of seed development (Boutrot et al. 2005; Drea et al. 2005; McIntosh et al. 2007).
The antimicrobial properties of ns-LTPs appear to emanate from their characteristic tertiary structures that allow membrane interaction. The ns-LTP1 proteins have eight conserved Cys residues (but lack Trp residues) and exhibit in their secondary structure a bundle of four helices separated by flexible loops, held together by four disulphide bridges (Marion et al. 1994; Kader 1996; Douliez et al. 2000). A hydrophobic cleft in the C-terminal half comprises the lipid-binding site that mediates lipid transfers. Curiously, certain LTPs (e.g., onion Ace-AMP1) appear to have evolved such that they retain the ability to bind to lipid bilayers and change membrane permeability, features required for antimicrobial activities, but lack the hydrophobic cavity required for lipid transfer (Tassin et al. 1998), a situation analogous to puroindolines. Acc-AMP1 can interact with bilayer vesicles but with less efficiency than PINA (Tassin et al. 1998). Other lipid-binding proteins such as purothionins and inhibitors of exogenous alpha-amylases also show similar structures and have roles in plant defence (Garcia-Olmedo et al. 1998). Purothionins are shown to cause lysis of mammalian cells by formation of cation channels, which, interestingly, may be composed of polymeric complexes (Hughes et al. 2000).
Tertiary structure and membrane interaction of puroindolines
As the puroindolines and related proteins are highly conserved in the closely related cereal species (e.g., wheat, barley, rye, oat) but absent in durum wheat and more distantly related cereals (e.g., maize, sorghum, rice) (Bhave and Morris 2007), they are unlikely to have physiological roles that are obligatory for plant survival. The PINs (and GSP-1) are clearly lipid-binding proteins, indicated by properties such as extraction with the non-ionic detergent Triton X-114 (Morris 2002), interaction with phosphatidylcholine (Le Guerneve et al. 1998) and ability to bind to bilayer vesicles (see below). Both PINA and PINB contain a backbone of 10 Cys residues including a Cys-Cys pair and a Cys-X-Cys triplet, and form a tertiary structure very similar to that of ns-LTPs, comprised of four alpha helices separated by loops and stabilised by five disulphide bridges (instead of four, due to the two extra Cys compared to ns-LTPs). Both proteins also contain a conserved Tyr residue in helix 1 which may be functionally important (Le Bihan et al. 1996).
The similarity between structures of ns-LTPs and PINs has resulted in expectations that they would share certain biochemical roles. PINs, ns-LTPs, and amylase and protease inhibitors are all suggested to be members of a large family that originated from a common ‘helicoidal’ ancestral protein and have inhibitory activities against plant predators and pathogens (Le Bihan et al. 1996; Shewry et al. 2004). Both PINs contain an additional tryptophan (Trp) rich domain (TRD) compared to ns-LTPs, composed of five Trp residues in PINA but only three in PINB. The Trps occupy a surface loop, the TRD forming an extension of it, stabilised by a Cys28/Cys48 disulphide bridge in PINA and Cys29/Cys48 in PINB (Le Bihan et al. 1996). In order to investigate the mechanisms of interaction of PINs with lipid membranes, purified PINs as well as model peptides including those mimicking their TRD, and model membranes or bilayer vesicles of different compositions, have been used extensively. The Trp residues as well as the basic amino acid residues interspersed between them, and the basic nature of the proteins in general, are proposed to be important for the lipid binding and membrane insertion properties of PINs (Kooijman et al. 1997; Le Guerneve et al. 1998). The Trps are shown to comprise the lipid-binding site (Kooijman et al. 1997, 1998). Importantly, Kooijman et al. (1997) showed that there are two types of interactions between PINs and membrane lipids: hydrophobic interactions between the Trp residues and lipid tails, and electrostatic interactions between the Arg/Lys residues in the TRD and the phosphate headgroups of lipids. Essentially, the TRD is the most important region of PINs for interaction with lipids, with the size, shape and partitioning properties of the Trp residues influencing their membrane binding (Kooijman et al. 1998; Le Guerneve et al. 1998; Yau et al. 1998; de Planque et al. 1999; Schibli et al. 2002; Jing et al. 2003). The charge and nature of the polar headgroups of lipids as well as chain packing and acyl chain length appear to influence the exact location of PINA in membranes (Kooijman et al. 1997; Le Guerneve et al. 1998; Jing et al. 2003), and the phenylalanine domain flanking the TRD (Marion et al. 1994) may also have a stabilising effect on binding of PINs (Kooijman et al. 1997). PINA appears able to penetrate lipid membranes more deeply than ns-LTPs, which have weaker, lipid interface-only binding (Kooijman et al. 1998) and PINB, which has fewer Trp and basic residues in its TRD. Both PINA and PINB interact more with negatively charged phospholipids than with neutral ones and possibly insert deeper into the former (Dubreil et al. 1997; Le Guerneve et al. 1998), confirmed by studying the interactions between each of the PIN proteins and lipids purified from wheat flour (Biswas and Marion 2006). Consequently, they have more affinity for bacterial membranes than the neutral eukaryotic (including mammalian) ones, thus greater antimicrobial than haemolytic activity (Kooijman et al. 1997; Le Guerneve et al. 1998; Douliez et al. 2000; Jing et al. 2003). The mechanism of antimicrobial activity was proposed to be like that of ns-LTPs: membrane leakiness caused by perturbation of lipid packing (Le Guerneve et al. 1998; Jing et al. 2003; Tassin et al. 1998), rather than by creation of protein channels or pores as in case of purothionins (Hughes et al. 2000). In contradiction, Charnet et al. (2003) showed that PINA could form cation channels in Xenopus oocytes and artificial membranes, confirmed by Llanos et al. (2006) in giant liposomes. However, it was ineffective compared to α1-purothionin in causing responses in murine neuromuscular cells or cytotoxicity to neuroblastoma cells, confirming that its interactions with microbial and mammalian cells differ. PINA forms aggregates (and PINB too, but less so), which may be relevant to their functions, as other Trp-rich proteins including a membranotoxin are reported to form aggregates (Le Bihan et al. 1996) and peptide pores often require oligomers (Jing et al. 2003). From early on, puroindolines have been suggested to be membranotoxins with primary roles in seed or seedling defence against microbial pathogens (Gautier et al. 1994; Le Bihan et al. 1996). The above reports of the membrane-active nature of PINs strongly support this possibility and are also relevant to their effects on kernel texture; both subjects are discussed below.
Individual yet co-operative roles for PINA and PINB in binding to starch granules
As mentioned earlier, the biochemical basis of soft kernel texture of wheat is now understood to require the presence of wild-type PINA and PINB. Soft kernel texture is also associated with the ‘friabilin’ phenomenon (Morris 2002) wherein some kernel PINA and PINB co-isolates during the water isolation of starch from soft wheat but minimally so from hard wheat. The plausible explanation for this relates to the ability of the puroindolines to bind lipids and insert into lipid bilayers making this association possible with the polar lipids on the surface of starch granules. Their sub-cellular in situ location is yet to be fully resolved. Considerable information is now available to analyse the functional relationship between the two PIN proteins and their role in kernel texture. Several reports comparing transcript or protein levels in different wheats show a primary role for PINA in binding to starch granules and a role in mediating the binding of PINB to starch, with a minor but co-operative role for PINB (Corona et al. 2001; Capparelli et al. 2003; Amoroso et al. 2004; Gazza et al. 2005). The PINA-null allele Pina-D1b or the SNP allele Pina-D1m have higher hardness values than all SNP alleles of Pinb-D1 for which such values are known (Giroux et al. 2000; Chen et al. 2006), supporting a greater role for PINA than for PINB in affecting hardness. However, the reduced quantities of PINA in Pinb-D1b mutants also suggest a reciprocal role for PINB in assisting the binding of PINA to starch (Amoroso et al. 2004).
Transgenic work has provided more direct insight into the in vivo roles and interrelationships of PINA and PINB. Rice lacks Pin genes and is hard-textured (Gautier et al. 2000), making it an ideal model system to assess the effects of added puroindolines. Krishnamurthy and Giroux (2001) showed that transgenic rice lines transformed with wheat wild-type Pina and/or Pinb genes and expressing either or both proteins exhibited a decrease in hardness, providing compelling proof for the causative role of puroindolines. Beecher et al. (2002) successfully complemented the hard phenotype of ‘Hi-Line’ Pinb-D1b wheat with an introduced wild-type Pinb-D1a allele, resulting in soft-kernel transgenic plants, confirming that PINB is also a functional determinant of the soft phenotype. Hogg et al. (2004) showed in further work, using lines from these stocks expressing the added soft-type PINA and/or PINB, that both proteins were individually able to reduce hardness but that addition of PINB resulted in softer kernels than addition of PINA, raising the question whether PINA was less active than PINB in this genetic background (which expressed a mutant form of PINB). To address this issue, Martin et al. (2006) transformed PINA-null (Pina-D1b/Pinb-D1a) ‘Bobwhite’ wheat with wild-type Pina-D1a and obtained softer kernels with significantly increased proportions of starch-bound PINA and PINB compared to the PINA-null parent and a soft wheat line used as control, further confirming that both proteins are required in their wild-type for the soft phenotype. Swan et al. (2006) crossed transgenic lines created in cultivar (cv.). Hi-Line (Beecher et al. 2002; Hogg et al. 2004) with the soft wheat cv. Heron. The progeny that expressed added PINB were much softer than those with added PINA and showed an increase in both PINA and PINB bound to starch, while progeny with added PINA only showed an increase in starch-bound PINA, indicating that PINB was limiting in the sense that it assists PINA in binding to starch. However, a limitation in this experiment was the intrinsic PINA. Wanguji et al. (2007) crossed the transgenic wheat lines expressing either added protein (Hogg et al. 2004) with those null for PINA (Pina-D1b) or PINB (Pinb-D1e). The segregation analysis of the progeny showed that each protein was able to associate with starch granules without the other but leads only to intermediate hardness, and that both proteins in their wild-type state are required for the soft phenotype. The causative role of PINA in texture was also confirmed in transgenic plants with added copies of Pina- D1a, created in the soft cultivar Zhongyou 9507–60 background (Xia et al. 2007). However, the biochemical basis for such effects and exactly how the two proteins interact are not yet understood. Le Bihan et al. (1996) showed that PINA can form aggregates and that PINB is less likely to do so, which might be important to their function, especially if they form pores during membrane insertion similar to other antimicrobial proteins (Le Bihan et al. 1996; Hughes et al. 2000; Jing et al. 2003). Giroux and Morris (1998) suggested heterodimer formation, and Ikeda et al. (2005) reported that the amount of GSP-1 was higher in PINA-null wheats (alleles Pina-D1b and Pina-D1k; Morris and Bhave 2007), which suggests some ‘cross-talk’ with GSP-1, adding further intrigue to interactions within this group of proteins. The protein–protein interactions, the individual yet interdependent roles of the wild-type proteins, and any changes to these in the mutant/variant proteins need to be tested rigorously.
Experimental evidence for antimicrobial activities of puroindolines
There is a significant amount of data accumulating on this subject. Purified PINA, PINB and ns-LTPs show antifungal properties (Dubreil et al. 1998), the antimicrobial action of PINA seems to be species-dependent (Mourgues et al. 1998), and Gautier et al. (2000) suggest that the difference in tolerance of common and durum wheat to disease, especially Fusarium head blight, could be related to their Pin genotypes, durum wheat lacking these genes. Synthetic peptides mimicking the TRDs of PINA and PINB exhibit significant antimicrobial activity against both Gram-positive and Gram-negative bacteria (Jing et al. 2003), and their potential as antimicrobial agents is further supported by studies using model membranes and/or microbial and fungal cells (Rezansoff et al. 2005; Vila-Perello et al. 2006). Capparelli et al. (2005) showed that both PINs localised to the aleurone and endosperm and could inhibit the growth of both Gram-positive and Gram-negative bacteria, possibly synergistically. Expression of PINA and PINB, such as that of wheat ns-LTPs in E. coli (Elmorjani et al. 2004) and PINA in yeast cells (Issaly et al. 2001), will help with structural studies as well as testing their antimicrobial properties and technological applications. It will also be important to express PIN and GSP-1 proteins encoded by different alleles in wheat and the related proteins in other cereals to test their antimicrobial potential and any differences therein.
Roles of GSP-1 in influencing kernel texture-the ambiguities
Compared to the relatively limited range of sequence variations noted in the Pina and Pinb genes from thousands of varieties and land races of common wheat tested worldwide, the Gsp-1 genes appear very diverse, even in their limited analyses (Bhave and Morris 2007). This tolerance of diversity makes the biological role(s) of GSP-1 unclear. On one hand, no SNPs in Gsp-1 genes corresponding to the hardness-associated mutations in Pin genes have been reported, and it is questioned whether the only two Trp residues in the TRD would suffice for lipid-binding and starch granule association properties. As mentioned earlier, no differences have been found in the extent of transcription or sequences of Gsp-1 genes between hard and soft wheats, and deletions or allelic variations of Gsp-1 in chromosome substitution and deletion lines had little effect on kernel texture. These observations suggest a role for Gsp-1 in (a)effecting kernel softness is unlikely. On the other hand, reports of Gsp-1 deletions are very few and the two Trp residues of the TRD and the 10 Cys are invariant despite the sequence diversity, suggesting the gene is under selection pressure. Analysis of published data (e.g., Morris et al. 2001; Chen et al. 2006; Lillemo et al. 2006) shows that common wheat cultivars or land races with identical Pina and Pinb alleles often display hardness values varying significantly (by 20–25 SKCS units) within the respective groups, even between plants grown under identical conditions, and true medium-soft types of common wheat have been identified (Lillemo and Ringlund 2002). Significant variation has been noted among progeny from parents of the same textural class, including soft wheats (Baker and Sutherland 1991) or between parents with different hardness alleles, with only a small proportion of the variation being attributed to allelic differences (Giroux et al. 2000). Significant differences are also noted between durum accessions, even those grown at identical sites (Dessalegn et al. 2006; G. Grimes and R. Hare personal communication), all of which lack PINs. While some of the variation could be due to the minor to moderate effects of environmental and biochemical factors on kernel texture of common or durum wheats (Tranquilli et al. 2002; Turnbull and Rahman 2002; Dessalegn et al. 2006; Gazza et al. 2007), it is unlikely that some of the variation may involve GSP-1. A role for GSP-1 has been suggested in kernel softness (Rahman et al. 1994) and cell wall structure (Van den Bulck et al. 2002), and the work on synthetic wheats clearly established that variations in kernel texture could be assigned genetically to the durum parent and proposed a role for Gsp-1 and/or associated QTLs (Gedye et al. 2004). Further, the relative amount of a GSP-1 isoform was found to be higher in two common wheat cultivars lacking PINA and in a durum (Ikeda et al. 2005). In light of the reports of lower amounts of PINB in the absence of PINA (see earlier) suggesting their interaction, the increased amounts of GSP-1 suggests interesting possibilities such as its direct interactions with puroindolines.
It appears that there must be some selection pressure on GSP-1, perhaps through specialised roles for some gene paralogues, as suggested for multigene families (Zhang 2003), and/or antimicrobial roles. It is possible that the membrane-binding ability of PINs leads to binding to the starch granule surface or interactions with the amyloplast membrane, and the incidental effects on kernel texture have been selected by human intervention. GSP-1 may have a minor effect on kernel texture in common wheat due to its limited TRD, but may be more prominent in durum wheats due to their lack of PINs, potentially explaining some of the variations between them. Clearly the peculiarities of Gsp-1 genes confirm the validity of the statement ‘GSP sequence functionality has never been assessed and indeed the whole molecular system encoded by the Ha locus deserves further study’ (Simeone and Lafiandra 2005).
Other factors affecting wheat kernel texture
While the Pin genes are the major genetic determinants of kernel texture, together they do not appear to completely account for the differences in texture noted in studies worldwide or in studies of segregation of different hardness alleles. In addition to the effects of environmental factors mentioned earlier, a number of biochemical factors such as seed moisture content, arabinoxylans (pentosans) and free polar lipids also appear to affect kernel texture directly or indirectly (Lillemo and Rungland 2002; Turnbull and Rahman 2002; Konopka et al. 2005). The unusually wide variations noted among progeny of certain crosses are mentioned above, and numerous loci have been suggested to play relatively minor or major roles in influencing grain texture, directly or indirectly, and alone or in combination; some of these are reviewed in Turnbull and Rahman (2002). Other suggestions include a 66 kDa protein tightly associated with the Ha locus (Partridge et al. 2002), other amphiphilic proteins (Amiour et al. 2003), QTLs on chromosomes 1AS, 1BL and 6B (Breseghello et al. 2005), anonymous loci with major or minor influences (Campbell et al. 1999; Giroux et al. 2000; Lillemo and Ringlund 2002) and loci on 1A and 6D (Perretant et al. 2000) and 1B (Turner et al. 2004). Of particular interest are minor QTLs on the short arm of chromosome 5A in a position homologous to Ha and on its long arm (Turner et al. 2004) and a second Pina copy and a major QTL on the long arm of 5D (Igrejas et al. 2002; Gazza et al. 2003).
Applications of puroindolines
Utilisation for kernel texture manipulation
The effects of some of the mutations in the Pin alleles on kernel texture provide the means to utilise classical plant breeding techniques to modulate the current range of kernel texture. A number of synthetic wheats are also being developed with different durum and Ae. tauschii parents (Gedye et al. 2004; Lillemo et al. 2006), and a critical assessment of the contribution of both parents to kernel texture is essential, the genetic factors contributed by durums being essentially non-Pin-related, while those from Ae. tauschii (DD) parents could include Pin variations unique to this taxon. Any desirable genes from synthetic wheats can be brought into bread wheat conveniently. The presence of functional Pin genes (so far found to be ‘soft’ alleles) on the diploid wheat genomes makes it feasible to transfer these into common wheat to manipulate further the range in texture. Transfer of a segment of chromosome 5A from T. monococcum (AmAm) carrying the Pin genes into T. aestivum v. Chinese Spring was shown to result in significantly softer kernels (Tranquilli et al. 2002), and introduction of the Ha-containing segments of chromosome 5A or 5B from T. monococcum or T. searsii, respectively, into Chinese Spring and further crossing to obtain different Pin copy numbers (depending on whether one or both copies of chromosomes 5A and 5B had been substituted) resulted in a nonlinear but clear correlation between copy number and kernel softness (See et al. 2004). It appears that only the added genes from A genome could be further transferred to a local soft wheat variety (Campbell et al. 2007). Bonafide et al. (2007) have now reduced the Ha-containing region of T. monococcum to a smaller segment to avoid transfer of a detrimental gene to bread wheat. Developing appropriate methods for assessment of any variations in kernel texture in diploid wheats and a critical assessment of the biochemical implications of sequence variations would open up further gateways.
Utilisation for antimicrobial properties
The potential applications of antimicrobial roles of PINs are just beginning to be explored. Krishnamurthy et al. (2001) showed that rice plants transformed with soft alleles Pina-D1a, Pinb-D1a, or both, exhibited increased resistance to fungal pathogens, the double transgenics being most resistant. No synergistic effects between the two proteins were noted, some differences were evident in the extent of resistance, and the mechanism of resistance appeared to involve slowing the spread of disease. Interestingly, in the transgenic plants, the genes were under the control of a constitutive promoter and expressed in the leaf tissue (where they are normally not expressed in wheat; see earlier), showing the potential of this method for creating plants (including wheat) with increased pathogen resistance throughout the plant rather than it being limited to seeds. Expression of PINA in a transgenic durum wheat similarly resulted in enhanced resistance to leaf rust (Luo et al. 2007) and that of PINB in transgenic apples resulted in resistance to a fungal disease (Faize et al. 2004), showing the gene can be expressed in a woody dicot. Further, Capparelli et al. (2006, 2007) tested purified native and recombinant PINA and PINB; all proteins were potent against E. coli, both recombinant proteins inhibited growth of S. epidermidis that causes skin infections such as acne, PINB being more potent than PINA. High concentrations of the proteins were also effective against intracellular infections, indicating their potential for ectopic treatments. Such applications offer excellent future directions.
Conclusions
Accumulating evidence regarding the membrane-active properties of puroindolines indicates that antimicrobial defence in the seed is the most likely primary in-planta role of this group of proteins, the commercially significant aspect of their effects on kernel texture of bread wheat essentially being a side effect of the same phenomenon. It will be essential to conduct protein expression, interaction and mutagenesis studies to test the significance of the various amino acid residues in the secondary and tertiary structures of puroindolines, work out the mechanics of the co-operative and interdependent roles of puroindolines and perhaps GSP-1, and the effects of single and multiple mutations in PIN proteins on their lipid binding ability, in order to understand the effects of some of the naturally occurring alleles. The emerging areas of applications of PINs as ectopic agents for infection control, as well as use of transgenic technology for expressing PINs to impart pathogen resistance in not just cereal species but woody plants, provide strong foci for further work.
Abbreviations
- DAF:
-
Days after flowering (≈days post-anthesis)
- GSP:
-
Grain softness protein
- NILs:
-
Near-isogenic lines
- ns-LTPs:
-
Non-specific lipid-transfer proteins
- QTL:
-
Quantitative trait locus/loci
- SNP:
-
Single nucleotide polymorphism
- TRD:
-
Tryptophan-rich domain
References
Amiour N, Merlino M, Leroy P et al (2003) Chromosome mapping and identification of amphiphilic proteins of hexaploid wheat kernels. Theor Appl Genet 108:62–72
Amoroso MG, Longobardo L, Capparelli R (2004) Real time PCR and flow cytometry to investigate wheat kernel hardness: role of puroindoline genes and proteins. Biotechnol Lett 26:1731–1737
Baker RJ, Sutherland KA (1991) Inheritance of kernel hardness in five spring wheat crosses. Can J Plant Sci 71:179–181
Beecher B, Bettge A, Smidansky E et al (2002) Expression of wild-type pinB sequence in transgenic wheat complements a hard phenotype. Theor Appl Genet 105:870–877
Bhave M, Morris CF (2007) Molecular genetics of puroindolines and related genes: allelic diversity in wheat and other grasses. Plant Mol Biol (in press)
Biswas S, Marion D (2006) Interaction between puroindolines and the major polar lipids of wheat seed endosperm at the air–water interface. Colloids Surf B 53:167–174
Blein J, Coutos-Thevenot P, Marion D et al (2002) From elicitins to lipid-transfer proteins: a new insight I cell signaling involved in plant defence mechanisms. Trends Plant Sci 7:293–296
Bonafede M, Kong L, Tranquilli G et al (2007) Reduction of a Triticum monococcum chromosome segment carrying the softness genes Pina and Pinb translocated to bread wheat. Crop Sci 47:819–826
Bottley A, Xia GM, Koebner RMD (2006) Homeologous gene silencing in hexapolid wheat. Plant J 47:897–906
Boutrot F, Guirao A, Alary R et al (2005) Wheat non-specific lipid transfer protein genes display a complex patterns of expression in developing seeds. Biochim Biophys Acta 1730:114–125
Breseghello F, Finney PL, Gaines C et al (2005) Genetic loci related to kernel quality differences between a hard and soft wheat cultivar. Crop Sci 45:1685–1695
Cameron K, Teece M, Smart L (2006) Increased accumulation of cuticular wax and expression of lipid transfer protein in response to periodic drying in leaves of tree tobacco. Plant Physiol 140:176–183
Campbell KG, Bergman CJ, Gualberto DG et al (1999) Quantitative trait loci associated with kernel traits in a soft × hard wheat cross. Crop Sci 39:1184–1195
Campbell JB, Martin JM, Crutcher F et al (2007) Effects on soft wheat (Triticum aestivum L.) quality of increased Puroindoline dosage. Cereal Chem 84:80–87
Capparelli R, Borriello G, Giroux MJ et al (2003) Puroindoline-a gene expression is involved in association of puroindolines to starch. Theor Appl Genet 107:1463–1468
Capparelli R, Amoroso MG, Palumbo D et al (2005) Two plant puroindolines colocalise in wheat seed and in vitro synergistically fight against pathogens. Plant Mol Biol 58:857–867
Capparelli R, Palumbo D, Iannaccone M et al (2006) Cloning and expression of two plant proteins: similar antimicrobial activity of native and recombinant form. Biotechnol Lett 28:943–949
Capparelli R, Ventimiglia I, Palumbo D et al (2007) Expression of recombinant puroindolines for the treatment of staphylococcal skin infections (Acne vulgaris). J Biotechnol 128:606–614
Charnet P, Molle G, Marion D et al (2003) Puroindolines form ion channels in biological membranes. Biophys J 84:2416–2426
Chen M, Wilkinson M, Tosi P et al (2005) Novel puroindoline and grain softness protein alleles in Aegilops species with the C, D, S, M and U genomes. Theor Appl Genet 111:1159–1166
Chen F, He Z, Xia X et al (2006) Molecular and biochemical characterisation of puroindoline a and b alleles in Chinese landraces and historical cultivars. Theor Appl Genet 112:400–409
Clarke B, Rahman S (2005) A microarray analysis of wheat grain hardness. Theor Appl Genet 110:1259–1267
Clifton LA, Lad MD, Green RJ et al (2007) Single amino acid substitutions in puroindoline-b mutants influence lipid binding properties. Biochem 46:2260–2266
Corona V, Gazza L, Boggini G et al (2001) Variation in friabilin composition as determined by A-PAGE fractionation and PCR amplification, and its relationship to grain hardness in bread wheat. J Cereal Sci 34:243–250
Day L, Bhandari D, Greenwell P et al (2006) Characterisation of wheat puroindoline proteins. FEBS J 273:5358–5373
De Planque MRR, Kruijtzer J, Liskamp R et al (1999) Different membrane anchoring positions of tryptophan and lysine in synthetic transmembrane α-helical peptides. J Biol Chem 274:20839–20846
Dessalegn T, Labuschagne MT, van Deventer CS (2006) Quality of Ethiopean durum wheat lines in two diverse environments. J Agron Crop Sci 192:147–150
Digeon JF, Guiderdoni E, Alary R et al (1999) Cloning of a wheat puroindoline gene promoter by IPCR and analysis of promoter sequences required for tissue-specific expression in transgenic rice seeds. Plant Mol Biol 39:1101–1112
Douliez JP, Michon T, Elmorajani K et al (2000) Structure, biological and technological functions of lipid transfer proteins and indolines, the major lipid binding proteins from cereal kernels. J Cereal Sci 32:1–20
Drea S, Leader DJ, Arnold B et al (2005) Systematic spatial analysis of gene expression during wheat caryopsis development. Plant Cell 17:2172–2185
Dubreil L, Compoint JP, Marion D (1997) Interaction of puroindolines with wheat flour polar lipids determines their foaming properties. J Agric Food Chem 45:108–116
Dubreil L, Gabroit T, Bouchet B et al (1998) Spatial and temporal distribution of the major isoforms of puroindolines (puroindoline-a and puroindoline-b) and non-specific lipid transfer protein (nsLTPe1) of Triticum aestivum seeds. Relationships with their in vitro antifungal properties. Plant Sci 138:121–135
Elmorjani K, Lurquin V, Lelion A et al (2004) A bacterial expression system revisited for the recombinant production of cysteine-rich plant lipid transfer proteins. Bichem Biophys Res Commun 316:1202–1209
Faize M, Sourice S, Dupuis F et al (2004) Expression of wheat puroindoline-b reduces scab susceptibility in transgenic apple (Malus X domestica Borkh.). Plant Sci 167:347–354
Garcia-Olmedo F, Molina A, Alamillo J et al (1998) Plant defence peptides. Biopolymers (Peptide Science) 47:479–491
Gautier MF, Aleman ME, Guirao A et al (1994) Triticum aestivum puroindolines, two basic cysteine-rich seed proteins: cDNA sequence analysis and developmental gene expression. Plant Mol Biol 25:43–57
Gautier MF, Cosson P, Guirao A et al (2000) Puroindoline genes are highly conserved in diploid ancestor wheats and related species but absent in tetraploid Triticum species. Plant Sci 153:81–91
Gazza L, Niglio A, Vaccino P et al (2003) The long arm of chromosome 5D of bread wheat contains a Pina-D1a-like sequence. In: Pogna NE, Romano M, Pogna E, Galterio G (eds) Proc 10th intl wheat genet symp, vol. 3. Paestum, Italy, pp 1330–1332
Gazza L, Nocente F, Ng PKW et al (2005) Genetic and biochemical analysis of common wheat cultivars lacking puroindoline a. Theor Appl Genet 110:470–478
Gazza L, Taddei F, Corbellini M et al (2007) Genetic and environmental factors affecting grain texture in common wheat. J Cereal Sci. doi:10.1016/j.jcs.2007.01.004
Gedye KR, Morris CF, Bettge AD (2004) Determination and evaluation of the sequence and textural effects of the puroindoline a and puroindoline b genes in a population of synthetic hexaploid wheat. Theor Appl Genet 109:1597–1603
Giroux MJ, Morris CF (1997) A glycine to serine change in puroindoline-b is associated with wheat grain hardness and low levels of starch-surface friabilin. Theor Appl Genet 95:857–864
Giroux MJ, Morris CF (1998) Wheat grain hardness results from highly conserved mutations in the friabilin components puroindoline-a and -b. Proc Natl Acad Sci USA 95:6262–6266
Giroux MJ, Talbert L, Habernicht DK et al (2000) Association of puroindoline sequence type and grain hardness in hard red spring wheat. Crop Sci 30:370–374
Hogg AC, Sripo T, Beecher B et al (2004) Wheat puroindolines interact to form friabilin and control wheat grain hardness. Theor Appl Genet 108:1089–1097
Hughes P, Dennis E, Whitecross M et al (2000) The cytotoxic plant protein, β-purothionin, forms ion channels in lipid membranes. J Biol Chem 275:823–827
Igrejas G, Leroy P, Charmet G et al (2002) Mapping QTLs for grain hardness and puroindoline content in wheat (Triticum aestivum L.). Theor Appl Genet 106:19–27
Ikeda TM, Ohnishi N, Nagamine T et al (2005) Identification of new puroindoline genotypes and their relationship to flour texture among wheat cultivars. J Cereal Sci 41:1–6
Issaly N, Soisona O, Joudrier P et al (2001) Optimisation of the wheat puroindoline-a production in Pichia pastoris. J Appl Microbiol 90:397–406
Jing W, Demcoe A, Vogel HJ (2003) Conformation of a bactericidal domain of puroindoline a: structure and mechanism of action of a 13-residue antimicrobial peptide. J Bacteriol 185:4938–4947
Kader J-C (1996) Lipid-transfer proteins in plants. Annu Rev Plant Physiol Plant Mol Biol 47:627–654
Kan Y, Wan Y, Beaudoin F et al (2006) Transcriptome analysis reveals differentially expressed storage protein transcripts in seeds of Aegilops and wheat. J Cereal Sci 44:75–85
Konopka I, Rotkiewicz D, Tanska M (2005) Wheat endosperm hardness. Part II. Relationship to content and composition of flour lipids. Eur Food Res Technol 220:20–24
Kooijman M, Orsel R, Hessing M et al (1997) Spectroscopic characterisation of the lipid-binding properties of wheat puroindolines. J Cereal Sci 26:145–159
Kooijman M, Orsel R, Hamer RJ et al (1998) The insertion behaviour of wheat puroindoline-a into diacylgalactosylglycerol films. J Cereal Sci 28:43–51
Krishnamurthy K, Giroux M (2001) Expression of wheat puroindoline genes in transgenic rice confers grain softness. Nat Biotechnol 19:162–166
Krishnamurthy K, Balconi C, Sherwood JE et al (2001) Wheat puroindolines enhance fungal disease resistance in transgenic rice. Mol Plant Microbe Interact 14:1255–1260
Laudencia-Chingcuanco DL, Stamova BS et al (2007) Transcriptional profiling of wheat caryopsis development using cDNA microarrays. Plant Mol Biol 63:651–668
Le Bihan T, Blochet JE, Desormeaux A et al (1996) Determination of the secondary structure and conformation of puroindolines by infrared and Raman spectroscopy. Biochemistry 35:12712–12722
Le Guerneve C, Seigneuret M, Marion D (1998) Interaction of the wheat endosperm lipid-binding protein puroindoline-a with phospholipids. Arch Biochem Biophys 360:179–186
Lillemo M, Ringlund K (2002) Impact of puroindoline b alleles on the genetic variation for hardness in soft × hard wheat crosses. Plant Breed 121:210–217
Lillemo M, Simeone MC, Morris CF (2002) Analysis of puroindoline a and b sequences from Triticum aestivum cv. ‘Penawawa’ and related taxa. Euphytica 126:321–331
Lillemo M, Chen F, Xia X et al (2006) Puroindoline grain hardness alleles in CIMMYT bread wheat germplasm. J Cereal Sci 44:86–92
Llanos P, Henriquez M, Minic J et al (2006) Puroindoline-a and α1-purothionin form ion channels in giant liposomes but exert different toxic actions on murine cells. FEBS J 273:1710–1722
Luo L, Zhang J, Yang G et al (2007) Expression of puroindoline A enhances leaf rust resistance in transgenic tetraploid wheat. Mol Biol Rep. doi:10.1007/s11033-007-9070-x
Marion D, Gautier MF, Joudrier P et al (1994) Structure and function of wheat lipid binding proteins. In: Wheat kernel proteins: molecular and functional aspects, Proc Int’l Mtg. Universita Degli Sudi della Tuscia, Viterbo, pp 175–180
Martin JM, Meyer FD, Smidansky ED et al (2006) Complementation of the pina (null) allele with the wild type Pina sequence restores a soft phenotype in transgenic wheat. Theor Appl Genet 113:1563–1570
McIntosh S, Watson L, Bundock P et al (2007) SAGE of the developing wheat caryopsis. Plant Biotechnol J 5:69–83
Morris CF (2002) Puroindolines: the molecular genetic basis of wheat grain hardness. Plant Mol Biol 48:633–647
Morris CF, Bhave M (2007) Reconciliation of D-genome puroindoline allele designations with current DNA sequence data. J Cereal Sci (in press)
Morris CF, Lillemo M, Simeone MC et al (2001) Prevalence of puroindoline grain hardness genotypes among historically significant North American spring and winter wheats. Crop Sci 41:218–228
Mourgues F, Brisset M-N, Chevreau E (1998) Activity of different antimicrobial peptides on Erwinia amylovora growth, and evaluation of the phytotoxicity and stability of cecropins. Plant Sci 139:83–91
Partridge MAK, Appels R, Skerritt JH (2002) Simple ELISA detection of a new polymorphic Ha locus encoded protein. J Cereal Sci 35:189–200
Perretant MR, Cadalen T, Charnet G et al (2000) QTL analysis of bread-making quality in wheat using a doubled haploid population. Theor Appl Genet 100:1167–1175
Perrocheau L, Bakan B, Boivin P et al (2006) Stability of barley and malt lipid transfer protein 1 (LTP1) toward heating and reducing agents: relationships with the brewing process. J Agric Food Chem 54:3108–3113
Rahman S, Jolly CJ, Skerritt JH et al (1994) Cloning of a wheat 15 kDa grain softness protein (GSP). GSP is a mixture of different puroindoline-like polypeptides. Eur J Biochem 223:917–925
Rezansoff AJ, Hunter HN, Jing W et al (2005) Interactions of the antimicrobial peptide Ac-FRWWHR-NH2 with model membrane systems and bacterial cells. J Peptide Res 65:491–501
Schibli DJ, Epand RF, Vogel HJ et al (2002) Tryptophan-rich antimicrobial peptides: comparative properties and membrane interactions. Biochem Cell Biol 80:667–677
See DR, Giroux M, Gill BS (2004) Effect of multiple copies of puroindoline genes on grain softness. Crop Sci 44:1248–1253
Shewry PR, Jenkins J, Beaudoin F et al (2004) The classification, functions and evolutionary relationships of plant proteins in relation to food allergens. In: Mills ENC, Shewry PR (eds) Plant food allergens. Blackwell Science, Oxford, pp 24–41
Simeone MC, Lafiandra D (2005) Isolation and characterization of friabilin genes in rye. J Cereal Sci 41:115–122
Simeone MC, Gedye KR, Mason-Gammer R et al (2006) Conserved regulatory elements identified from a comparative puroindoline gene sequence survey of Triticum and Aegilops diploid taxa. J Cereal Sci 44:21–33
Swan CG, Meyer FD, Hogg AC et al (2006) Puroindoline B limits binding of puroindoline A to starch and grain softness. Crop Sci 46:1656–1665
Tassin S, Broekaert WF, Marion D et al (1998) Solution structure of Ace-AMP1, a potent antimicrobial protein extracted from onion seeds. Structural analogies with plant non-specific lipd transfer proteins. Biochemistry 27:3623–3637
Tranquilli G, Heaton J, Chicaiza O et al (2002) Substitutions and deletions of genes related to grain hardness and their effect on grain texture. Crop Sci 42:1812–1817
Turnbull KM, Rahman S (2002) Endosperm texture in wheat. J Cereal Sci 36:327–337
Turnbull KM, Gaborit T, Marion D et al (2000) Variation in puroindoline polypeptides in Australian wheat cultivars in relation to grain hardness. Aus J Plant Physiol 55:89–95
Turnbull KM, Marion D, Gaborit T et al (2003a) Early expression of grain hardness in the developing wheat endosperm. Plants 216:699–706
Turnbull KM, Turner M, Mukai Y et al (2003b) The organization of genes tightly linked to the Ha locus in Aegilops tauschii, the D-genome donor to wheat. Genome 46:330–338
Turner M, Mukai Y, Leroy P et al (1999) The Ha locus of wheat: identification of a polymorphic region for tracing grain hardness in crosses. Genome 42:1242–1250
Turner AS, Bradburne RP, Fish L et al (2004) New quantitative trait loci influencing grain texture and protein content in bread wheat. J Cereal Sci 40:51–60
Van den Bulck K, Loosveld A, Courtin CM et al (2002) Amino acid sequence of wheat flour arabinogalactan peptide, identical to part of grain softness protein GSP-1, leads to improved structural model. Cereal Chem 79:329–331
Vila-Perello M, Tognon S, Sanchez-Vallet A et al (2006) A minimalist design approach to antimicrobial agents based on a thionin template. J Med Chem 49:448–451
Wanguji HW, Hogg AC, Martin JM et al (2007) The role of puroindoline A and B individually and in combination on grain hardness and starch association. Crop Sci 47:67–76
Xia L, Geng H, Chen X et al (2007) Silencing of puroindoline a alters the kernel texture in transgenic bread wheat. J Cereal Sci. doi:10.1016/j.jcs.2007.04.016
Yau WM, Wimley WC, Gawrisch K et al (1998) The preference of tryptophan for membrane interfaces. Biochemistry 37:14713–14718
Zhang J (2003) Evolution by gene duplication: an update. Trends Ecol Evol 18:292–298
Acknowledgements
The authors thank the anonymous reviewer for a through and critical review of the manuscript, which helped greatly with its revision. The assistance of Stacey Sykes in the preparation of this manuscript is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bhave, M., Morris, C.F. Molecular genetics of puroindolines and related genes: regulation of expression, membrane binding properties and applications. Plant Mol Biol 66, 221–231 (2008). https://doi.org/10.1007/s11103-007-9264-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-007-9264-6