
Abstract
This review highlights recent findings of our group on bioactive marine natural products isolated from marine sponges and marine derived fungi. The activated chemical defence of the Mediterranean sponge Aplysina aerophoba is introduced as an example of a dynamic response of marine sponges to wounding. Following tissue disrupture preformed brominated isoxazoline alkaloids are enzymatically cleaved and thereby give rise to aeroplysinin-1 which is believed to protect sponges from invasion of pathogenic bacteria. A preliminary characterization of the membrane bound enzyme(s) involved in the cleavage reaction is presented. Bromotyrosine derived, oxime group bearing peptides, the so called bastadins, obtained from the sponge Ianthella basta and some of their synthetic derivatives were shown to exhibit pronounced antifouling activity against larvae of the barnacle Balanus improvisus. The antifouling activity could be traced to the oxime group as an important pharmacophore that was also found to be present in other sponge derived natural products exhibiting antifouling activity. Marine derived fungi that reside within invertebrates such as sponges or inside Mangrove plants are emerging as a new source of bioactive metabolites as demonstrated for Aspergillus ustus and Alternaria sp. that were isolated from the sponge Suberites domuncula or from the Mangrove plant Sonneratia alba, respectively. The former fungus yielded new moderately cytotoxic sesquiterpenoids of the drimane type whereas the latter was found to produce polyketides such as alternariol that exhibited strong and selective inhibitory activity against several protein kinases, for instance Aurora A and B which are targets for anticancer chemotherapy.
Similar content being viewed by others


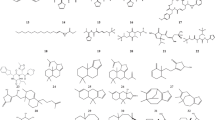
Avoid common mistakes on your manuscript.
Introduction
The rich and unprecedented chemical diversity of marine natural products and their pronounced biological activities continue to thrill natural product chemists, ecologists and pharmacologists alike. It is now clear that the oceans are not only home to a tremendous diversity of species but that their inhabitants produce also a wealth of natural products (Blunt et al. 2009) that are intertwined in intra- and interspecific chemical communication (Paul and Ritson-Williams 2008). Apart from their ecological role, marine natural products are a treasure cove of evolutionary pre-selected lead structures that are indispensable for drug discovery as highlighted by the recent introduction of the marine derived pain killer Prialt® and the anti-tumor drug Yondelis® to the market. Traditionally, marine invertebrates such as sponges, molluscs, tunicates and others as well as algae have been in the focus of marine natural product research. These sessile or slow moving inhabitants of the oceans rely mainly on the accumulation of toxic and/or deterrent chemicals for defence against predators, herbivores, pathogens or during allelopathic competition with neighbours for resources or space. Elucidating the manifold ecological roles of bioactive metabolites continues to be a fascinating element of marine natural product research.
This aspect is covered in our review article tackling the activated chemical defence of the Mediterranean sponge Aplysina aerophoba as an example. This sponge has long been considered as unique in the marine environment since preformed secondary metabolites (brominated isoxazoline alkaloids) are enzymatically cleaved during tissue damage thereby generating the powerful defence metabolite aeroplysinin-1 (Thoms et al. 2006) which shows a multitude of biological activities (Weiss et al. 1996) and is considered to protect the sponge against invading microbial pathogens. Only recently did marine chemical ecologists become aware of similar defence strategies that are present also in other sponges indicating that this might be a more widely occurring phenomenon in the marine habitat (Thoms and Schupp 2008).
The link from basic research to applied science is sometimes short and obvious. In marine natural product research ecological observations can be helpful in identifying organisms that are likely to accumulate bioactive constituents that can be of interest in search of useful compounds for drug discovery, as plant protectants or as anti-fouling constituents. The observation that numerous marine invertebrates are suspiciously free of fouling organisms has repeatedly sparked the search for such constituents from nature that might provide environmentally friendly solutions in replacing broad band toxicants such as tri-n-butyl tin oxide (TBTO) and others. In this review we will provide an example for the successful search of new anti-fouling compounds from the marine sponge Ianthella basta.
Whereas in the last decades the focus in marine natural product research was mainly on macroorganisms such as sponges, sea weeds and others there is now increasing evidence that cyanobacteria, bacteria and fungi from the marine habitat are equally rich sources of novel constituents that are far more accessible through sustainable production by fermentation than many marine macroorganisms that usually have to be collected from limited wild stocks. We will give examples for interesting chemical profiles of marine derived fungi focussing on sponge and Mangrove endophytes.
Overall it is the aim of this review to highlight different facets of marine natural product research that range from basic science to applied aspects and to present some of the major research topics that have been going on in our group in recent years.
Activated chemical defence of the sponge Aplysina aerophoba—preliminary characterization of the enzymatic basis
Most chemical defence strategies reported so far for marine invertebrates or algae are constitutive and rely on preformed toxic or deterrent compounds that are present in tissues and either liberated upon wounding or constantly exuded (Walker et al. 1985). In contrast, activated defence systems represent a highly dynamic defence reaction (Paul and Van Alstyne 1992). Activated defence mechanisms are fast and occur mostly within seconds after cell compartmentalisation is disturbed. Protoxins get into contact with liberating enzymes that catalyse a biotransformation of protoxins yielding the actual defence metabolite. The most prominent example from the terrestrial environment is the generation of the defence metabolite hydrocyanic acid from inactive precursors in cyanogenic plants (Conn 1979; Wajant and Effenberger 1996). However, activated defence mechanisms are not limited to the plant kingdom, but are also known from the marine environment. The first example in the marine habitat was described for tropical green algae of the genus Halimeda that contain halimedatetraacetate as a major metabolite. Upon tissue damage, halimedatetraacetate is (probably enzymatically catalysed) converted into the chemically highly reactive halimedatrial (Paul and Van Alstyne 1992) that causes an increase of feeding deterrency towards herbivorous fishes (Paul and Van Alstyne 1988). For sponges, the wound-induced biotransformation of isoxazoline alkaloids in Aplysina sponges was the first example proposed as an activated chemical defence strategy (Teeyapant and Proksch 1993).
Sponges of the genus Aplysina are known for their structurally diverse brominated isoxazoline alkaloids (Cimino et al. 1983; Ciminiello et al. 1994a, b, 1995, 1996a, b, 1997, 1999, 2000; Thoms et al. 2003a) which act as potent chemical defence against predators and microorganisms (Teeyapant et al. 1993b; Weiss et al. 1996; Encarnación-Dimayuga et al. 2003; Kelly et al. 2003; Thoms et al. 2004). Aplysina species occur in the Mediterranean Sea, the Atlantic Ocean, and in the Caribbean Sea (Pawlik et al. 1995) where they often contribute to the dominant sponges present. The Mediterranean Sea is home to two Aplysina species: Aplysina aerophoba which occurs in water depths as low as 1 m (Riedl 1983; Pansini 1997; Thoms et al. 2003b), and Aplysina cavernicola which prefers shaded caves and deeper habitats (40 m or lower) (Wilkinson and Vacelet 1979; Thoms et al. 2003b). Both A. aerophoba and A. cavernicola contain aerophobin-2 and aplysinamisin-1 as brominated isoxazoline alkaloids (Fig. 1), with the latter compound not always being found in A. aerophoba. The occurrence of aerothionin, isofistularin-3 and the yellowish pigments differs between both species (Teeyapant et al. 1993a; Ciminiello et al. 1997; Thoms et al. 2003a, 2004).

Biotransformation of Aplysina-alkaloids as described by Teeyapant und Proksch (1993) with aerophobin-2 as example
All Aplysina species known are prone to a wound-induced bioconversion of brominated isoxazoline alkaloids giving rise to the nitrile aeroplysinin-1 which in turn is transformed into a dienone (Fig. 1; Teeyapant and Proksch 1993; Ebel et al. 1997; Thoms et al. 2006). The bioconversion products aeroplysinin-1 and dienone show pronounced antibiotic activity against numerous marine and terrestrial bacteria (Teeyapant et al. 1993b; Weiss et al. 1996; Debitus et al. 1998). In contrast, their isoxazoline precursors exhibit no or only weak antibiotic activity, but rather act as feeding deterrents against fishes (Thoms et al. 2004).
The cleavage of Aplysina alkaloids represents an unusual enzymatically catalysed hydrolysis reaction, to which no biochemical equivalent is known up to now. A similar reaction is known in the field of organic chemistry, the so-called abnormal Beckmann fragmentation (McCarty 1970). During the course of this reaction, the C–C bond of an α-oximinoketone is split, yielding a nitrile, an alcohol and, upon addition of water, a carboxylic acid derivative. The brominated isoxazoline alkaloids can be understood as α-oximinoketones, where the keto function is part of an amide partial structure and the imino group is part of the heterocycle. In analogy to the abnormal Beckmann fragmentation, the C–C bond of the sponge alkaloids is split between the carbonyl function and the isoxazoline ring, and a nitrile, aeroplysinin-1, emerges during the course of the biotransformation (Fig. 1). The oxygen substituent of the oximino function is not liberated as water or alcohol, but remains linked to the nitrile aeroplysinin-1 as alcoholic hydroxyl function.
Several earlier studies already confirmed that the cleavage of isoxazoline Aplysina alkaloids is an enzymatically catalysed reaction (Teeyapant 1994; Ebel 1998; Fendert 2000). In experiments employing a crude protein extract of Aplysina sponges the optimal temperature determined for enzyme activity was 50°C, and optimal pH at pH 5.8 (Teeyapant 1994; Ebel 1998; Fendert 2000). The isoxazoline cleaving enzyme appears to be highly specific towards its substrates, the isoxazoline Aplysina alkaloids. By offering different natural or synthetic compounds as substrate analogues, several structural characteristics relevant for enzyme activity could be identified, (a) the presence of an intact spirocyclohexadienisoxazoline ring system, (b) the acid amide side chain, (c) the hydroxyl group at position C-1 and (d) the bromination pattern (Fendert 2000). For the second biotransformation step, which involves the transformation of aeroplysinin-1 into a dienone (Fig. 1), the involvement of a nitrile hydratase was hypothesized due to the structural characteristics of these two metabolites (Ebel 1998; Fendert 2000).
The fact that no biochemical equivalent of the cleavage of isoxazoline alkaloids is known up to now raises questions about the nature of the enzyme(s) catalysing this unusual reaction. We therefore aimed at the purification and characterisation of this exceptional protein. Although the isoxazoline cleaving enzyme could so far not be purified completely, we gained insight into several of its characteristics. Purification of lyophilised sponge cells via differential ultracentrifugation and subsequent ion exchange or affinity chromatography (employing the immobilized substrate aerophobin-2) yielded enzymatically active fractions that catalysed cleavage of the isoxazoline alkaloid aerophobin-2 in vitro giving rise to aeroplysinin-1. Tracking activity in fractions obtained from differential ultracentrifugation and treatment of the membrane fraction with cold sodium carbonate revealed that the isoxazoline cleaving enzyme is an integral membrane protein. The observation that during ion exchange chromatography activity was detected in fractions containing unbound protein, and in fractions that eluted later from the column, indicates that the isoxazoline cleaving enzyme is not a single enzyme, but that different proteins or isoforms of the same protein exist. Calibration by gel filtration column chromatography using a range of standards revealed an approximate size of 540 kDa for the active protein(s). Employment of affinity chromatography as alternative purification step following ultracentrifugation likewise yielded active fractions.
Specific biotin-labelling and subsequent detection via Western Blot led to the identification of five protein bands with affinity for aerophobin-2 in both, fractions obtained from affinity- and from ion exchange chromatography. In-gel trypsin digestion and subsequent mass analysis identified one of these proteins as actin (Fig. 2a). Another peptide sequence was obtained for a protein of approx. 60 kDa in active ion exchange fractions (Fig. 2b), for which no homologous sequences could be found in data banks. Actin is one of the most conserved proteins across eukaryotic species which is attributed to its acquisition of mechanisms for assembly dynamics and interactions with multiple motor and binding proteins, thus contributing to the constraint on actin evolution (Erickson 2007). A key feature of actin is its ability to bind and hydrolyse ATP. The transition from ATP-actin to ADP-bound actin provokes a conformational change which is central to the dynamic turnover of actin filaments. Among a complex set of proteins attached to the surfaces of cell membranes, actin is responsible for stabilising membranes by regulating the topography and mobility of the different transmembrane proteins. This submembranous network, often referred to as membrane skeleton, is composed of filaments of actin, actin-binding proteins such as spectrin, and connecting proteins that link the stabilising infrastructure to the overlaying membrane (Marchesi 1985). Since the isoxzoline cleaving enzyme(s) are integral membrane proteins, the fact that one of the aerophobin-2 binding proteins in A. aerophoba is an actin-type of protein can be explained by its role in the stabilisation of membrane proteins within cell membranes. It therefore seems plausible that an aerophobin-2 binding protein located in the cell membrane is connected with the actin cytoskeleton.

SDS-PAGE (1) and Western blot (2) of a an enzymatically active fraction obtained from affinity chromatography and b a fraction derived from an ion exchange column. In Western blots, aerophobin-2 binding proteins are specifically labelled. Proteins whose sequences were successfully determined after in-gel trypsin digestion and subsequent mass analysis are marked by arrows
No significant homologies were found for the peptide sequences obtained from the 60 kDa protein in active ion exchange fractions. This is probably due to the fact that in protein databases, only a limited amount of protein sequences is available from sponges. For the genus Aplysina, no protein sequences or cDNA databases have been published so far, therefore hampering the identification of peptide sequences if not highly conserved as in the case of actin.
In summary, a combination of differential ultracentrifugation and further purification of the solubilised membrane fraction via affinity chromatography or, alternatively, ion exchange chromatography yielded active fractions of sufficient purity to allow for the identification of aerophobin-2 binding proteins by specific labelling. Comparison with protein databases identified one of these proteins as actin. For the 60 kDa protein, no homologous peptide sequences were found, therefore its identity remains unclear. Based on the findings acquired during purification steps, it became evident that the cleavage of isoxazoline alkaloids is accomplished by several proteins. Whether these are functional on an individual basis or act synergistically remains open. The existence of an enzyme complex appears likely. However, in addition to the proteins identified in this work, further proteins involved in catalysing the cleavage of isoxazoline alkaloids might exist which do not possess affinity for aerophobin-2 or an accessible binding site and were thus not detected in the described experiments. Future approaches involving degenerated primers based on elucidated protein sequences obtained could help to shed more light on the identity of proteins involved in the cleavage of isoxazoline alkaloids.
Anti-fouling constituents from the sponge Ianthella basta
Biofouling is a process that involves natural and man made surfaces and substrates and describes their overgrowth by microorganisms (mainly film forming bacteria and unicellular algae) and macroorganisms such as algae, mussels and barnacles (Clare 1996a). Costs that are caused by biofouling e.g. on ship hulls (that reduce speed which in turn needs to be compensated by higher fuel consumption), in desalination plants or in power stations by overgrowth of turbines reach into billions of dollars annually. Until recently, toxic paint ingredients such as organotin (TBTO) were in use to control or reduce such overgrowth. Other paint ingredients include herbicides or copper oxide. All of these additives are potentially harmful to the environment (Alzieu et al. 1989) and major efforts have been undertaken to stop or reduce their use. The International Maritime Organisation (IMO) has put forward a resolution calling first for a step-wise reduction of the use of organotin compounds and then for a complete prohibition which became effective last year (2008) (IMO 2001). These actions have created the need for the development of new and less harmful anti-fouling strategies that involve chemical, physical or mechanical mechanisms (Bers and Wahl 2004).
It is well known that many marine invertebrates such as sponges are free from overgrowth of fouling organisms. Since sponges are filter-feeders such an overgrowth would likely be detrimental as inhaling pores could easily be clogged. Sponges are also known for their rich chemical diversity which includes numerous compounds with anti-fouling properties (Fusetani 2004). It is therefore tempting to speculate that sponges are chemically defended against fouling organisms much like they are able to fight off or deter predators by accumulation of defensive natural products (Paul and Ritson-Williams 2008). Direct proof for a chemical defence of sponges against fouling organisms such as barnacle larvae, however, is still lacking as compounds would need to be concentrated in cortex cell layers or even be exuded into the surrounding sea water in order to become effective. This has with a few notable exceptions (Thompson et al. 1985) rarely been investigated so far and deserves more attention by marine chemical ecologists. Whereas the ecological importance of sponge derived natural products for defence against fouling organisms is therefore still not entirely clear it has been demonstrated repeatedly that these compounds are able to interfere with the process of biofouling. Hence, they provide new and structurally unprecedented lead structures for the search of alternative anti-fouling agents for future applications.
During a screening study on anti-fouling activities of tropical marine sponges we noticed that a methanolic extract of the sponge Ianthella basta collected in Indonesia strongly suppressed the settlement of larvae of the barnacle Balanus improvisus which are a well established model for anti-fouling studies. Activity guided fractionation of the extract yielded several bastadin derivatives (1–4) as well as hemibastadin-1 (5) as active compounds (Ortlepp et al. 2007). Bastadins are bromotyrosine derived, oxime group bearing peptides which can be cyclic or linear. Tyrosine or tyramine building blocks may be linked not only through peptide bonds but also through ether bonds or by C–C bonds between two aromatic rings. Hemibastadin-1 (5) essentially represents a biogenetic building block of the bastadins which are mostly dimers of hemibastadin units. Earlier studies had already indicated various biological activities of bastadins including cytotoxcity against cancer cells (Pettit et al. 1996) or inhibitory activity towards inosine 5′-phosphate dehydrogenase (Jaspars et al. 1994). Bastadins are also known as modulators of ryanodine sensitive sarcoplasmatic reticulum calcium channels (Mack et al. 1994). Anti-fouling activity of bastadins had, however, not been reported earlier.
Compounds 1–5 inhibited settlement of barnacle larvae in a dose dependent manner at concentrations ranging from 1 to 10 μM (Fig. 3) and thus proved to be powerful anti-fouling compounds. Several of these compounds, however, showed also toxicity against the larvae as demonstrated for example for bastadin-16 (4) or for hemibastadin-1 (5). Toxicity of anti-fouling constituents is of course not desirable when looking for alternatives for synthetic compounds such as TBTO that are likewise toxic. In addition to the bromine substituents which are rarely found in natural products from terrestrial organisms but which occur rather frequently in marine natural products the presence of oxime groups in compounds 1–5 attracted our attention as naturally occurring oxime derivatives are very rare in nature. We chose hemibastadin-1 (5) which is easy to synthesize (Ortlepp et al. 2007) compared to the larger bastadin derivatives as a model compound in order to investigate the effects of bromination and of the oxime group on anti-fouling activity. Several analogues of hemibastadin-1 were synthesized that included 5,5′-dibromohemibastadin-1 (6), debromohemibastadin-1 (7) and L-tyrosinyltyramine (8) the latter featuring an amino instead of an oxime group. Debromohemibastadin-1 (7) which differed from hemibastadin-1 (5) only by the lack of bromine substituents proved to be clearly less active compared to 5 but nevertheless inhibited larval settling almost completely when assayed at a concentration of 100 μM (Fig. 4). This indicates that the presence of bromine atoms increases anti-fouling activity but is not an essential structural requirement. The results were different for L-tyrosinyltyramine (8) that deviates from debromohemibastadin-1 (7) by an exchange of the oxime group versus an amino function. Compound 8 proved to be completely inactive even when tested at a concentration of 100 μM thus providing evidence that the oxime function is essential for anti-fouling activity. Whereas hemibastadin-1 (5) still showed some toxicity towards barnacle larvae the synthetic product dibromohemibastadin-1 (6) was non-toxic but very effective in inhibiting larval settling (Fig. 4). Interestingly, hemibastadin-1 (5) as well as its analogues 6–8 were non-toxic to brine shrimp larvae (Artemia salina) which are an established model for assessment of ecotoxicity (Carballo et al. 2002).

Effects of the active natural products on B. improvisus cyprid settlement. Results are expressed as percentage settlement (±SE) and percentage living swimming cyprids (±SE). C = control, 10 = 10 μMol; 1 = 1 μMol; 0.1 = 0.1 μMol, n = 4. * Results significantly different from the solvent controls

Effects of bioinspired synthetic compounds on B. improvisus cyprid settlement. Results are expressed as percentage settlement (±SE) and percentage living swimming cyprids (±SE). C = control, 100 = 100 μMol; 10 = 10 μMol; 1 = 1 μMol; 0.1 = 0.1 μMol, n = 4
Further sponge-derived compounds featuring oxime groups (9–11) were also investigated in our study and two of them (9 and 10) were highly active against barnacle larvae (Ortlepp et al. 2007). The latter included aplysamine-2 (9) isolated from Pseudoceratina purpurea and psammaplin A (10) from Aplysinella rhax (Ortlepp et al. 2007). Both sponge derivatives showed similar anti-fouling activity when compared to the bastadins or to hemibastadin-1. Psammaplin A (9) was also slightly toxic. Other bominated natural products of sponge origin such as bromopyrrole derivatives like hymenidin (11), sceptrin (12) or ageliferin (13) that were also assayed for anti-fouling activities for comparative reasons proved to be completely inactive. On the other hand, the diterpene alkaloid agelasidine-D N-oxime (14) which had been isolated from the sponge Agelas nakamurai (Hertiani 2007) was active in suppressing larval settlement. Unlike compounds 1–10 this natural product is no bromotyrosine derivative. The sole unifying structural element that compound 14 and the derivatives 1–10 have in common is the oxime group which again underlines the importance of this unusual structural element for anti-fouling activity.
It is possible that the oxime substituent might generate nitric oxide and through this mechanism interfere with the settling behaviour of barnacle larvae. Another plausible explanation for the mode of action at least of the bastadin derivatives involves alteration of the intracellular Ca2+ level of the larvae. It is known that Ca2+ ions are important for settling of barnacle larvae (Rittschof et al. 1986). Reduction of intracellular Ca2+ ions inhibits larval settlement (Clare 1996b). Bastadins on the other hand interact with the ryanodine receptor and affect intracellular Ca2+ concentrations. Clearly, further studies are needed to elucidate the target(s) and mechanism of action of the oxime bearing natural products analyzed in this study. The accessibility of hemibastadin-1 (5) and of its derivatives through organic synthesis and their significant anti-fouling activity make these compounds attractive candidates for further investigations aiming at their possible utilization as constituents of anti-fouling paints in the future. Presently field experiments involving dibromohemibastadin-1 (6) are going on in order to test the anti fouling activity of this synthetic product under more relevant conditions.
Fungal endophytes as sources of bioactive compounds
All higher plants as well as algae and marine invertebrates like sponges are known to harbour a multitude of fungal endophytes (e.g. Proksch et al. 2008; Teuscher et al. 2006; Aly et al. 2008). For filter feeders such as sponges the origin of fungal endophytes and the true nature of their interaction with their hosts remains largely unclear. The fact that many fungi so far isolated from sponges are well known from terrestrial habitats where they ubiquitously occur might suggest that fungal spores which are washed into the sea are sequestered by sponges through filter feeding and are retained in the inhalant canals from which they can be isolated and subsequently cultivated. Furthermore, there has been no evidence reported so far indicating that fungi actively grow inside sponges suggesting that their presence in these invertebrates is in fact limited to spores. On the other hand, compounds obtained from marine isolates of ubiquitously occurring fungal genera such as Aspergillus or Penicillium differ often significantly from those of terrestrial strains (Edrada et al. 2002; Lin et al. 2003) indicating that at least on a biochemical level many sponge derived fungi are distinguished from their terrestrial counterparts. For fungal endophytes isolated from the inner tissues of marine algae or from halotolerant plants such as Mangrove trees there can on the other hand be no doubt that these fungal—host plant communities are more specific in nature as has been demonstrated by numerous investigations on the taxonomy of fungal endophytes that have either been isolated from their hosts or by culture independent molecular approaches (Zuccaro et al. 2003; Xu et al. 2009a). For both groups of fungal endophytes (associated with sponges or with halotolerant higher plants) new bioactive natural products have been isolated underlining the importance of fungal endophytes as sources of novel constituents for drug discovery programs. Our own research on natural products derived from endophytes focuses on fungi associated with sponges (Proksch et al. 2008; Liu et al. 2009) and on Mangrove derived fungi (Xu et al. 2009a, b; Kjer et al. 2009).
The Mediterranean sponge Suberites domuncula serves as a model for our group to study the diversity of culturable associated fungi from a single sponge taxon which had rarely been done by other researchers before (for a notable exception see Höller et al. 2000). A total of 21 different fungal taxa was isolated and identified at least down to the genus level (Proksch et al. 2008). The largest number of isolates from a single taxon was obtained for the genera/taxa Cladosporium, Penicillium, Engyodontium album, Phialophora and Petriella (listed in descending frequency). From these fungal isolates E. album had been previously isolated from the marine sources (Chellappan et al. 2006) whereas all other frequently occurring fungi isolated in this study represent more or less ubiquitous taxa that occur also in soil samples, as plant pathogens or as endophytes of terrestrial plants. All fungi isolated were cultivated and their extracts were submitted to screening for cytotoxicity against murine L5178 lymphoma cells. The extract of the fungus Petriella sp. displayed pronounced cytotoxocity and was chosen for activity-guided isolation. The cyclic tetrapeptide WF-3161 (15) previously reported from P. guttulata that had been isolated from a soil sample was identified as the major secondary product exhibiting cytotoxic activity. The amino acid sequence of the peptide is cyclo(L-Leu-L-Pip-L-Aoe-D-Phe) and the ED50 value against L5178Y cells was <0.1 μg/ml (Proksch et al. 2008). In addition to the peptide further bioactive constituents were isolated that included several pyrone derivatives (16–18). The major pyrone (16) was also strongly active and featured an ED50 value of 0.2 μg/ml against L5178Y cells.
From the various fungal isolates obtained from the sponge S. domuncula only one isolate was identified as Aspergillus ustus. Again, isolates of A. ustus are not restricted to the marine habitat but have also been reported from the terrestrial habitat. Based on the positive initial screening results A. ustus was likewise chosen for activity guided isolation of its secondary compounds. Ten drimane sesquiterpenoids including seven new compounds were isolated and identified (Liu et al. 2009). The compounds proved to be structurally related to the known natural products deoxyvudin (19) or to the lactonized strobilactone B (20). Most of the compounds, however, exhibited only moderate activity against the cell line chosen suggesting that the stronger cytotoxicity of the extract was mainly due to additive or synergistic effects of the individual constituents.
Mangroves, a consortium of higher plants that inhabit the intertidal zones in subtropical and tropical climates, are important ecologically as fish nurseries, pollution sinks, shoreline barriers, bird sanctuaries and wildlife refuges. Mangroves are exposed to extreme conditions of stress, i.e. varying levels of water and concentrations of salt. Mangrove trees and shrubs stand with their roots in salt or brackish waters making them extremely susceptible to clogging, while the aboveground organs are subjected to tidal fluctuations. The germinating seedling roots must elongate fast enough to anchor the new plantlet before they are swept away. These warm, moist biotopes provide ideal habitats for microorganisms, some of which are pathogenic. Thus, survival in this bio-consortium necessitates adapting to both abiotic and biotic stress, presupposing an active secondary metabolism of host plants as a defence mechanism to cope with these conditions of stress. Mangroves are well adapted to cope with biotic stress factors caused by herbivores or plant pathogens through accumulation of numerous biologically active natural products that include polyphenols, limonoids and polyketide derivatives with many of them exhibiting pronounced biological activities (e.g. Yang et al. 2006; Li et al. 2006; Bao et al. 2007; Cui et al. 2007). On the other hand, Mangroves are also hosts to numerous endophytic fungi that are likewise interesting sources of bioactive secondary metabolites.
An example is provided by a hitherto undescribed strain of the genus Alternaria isolated from the Mangrove plant Sonneratia alba from South China (Kjer et al. 2009). From the extract of the endophyte growing on solid rice medium fourteen different natural products were obtained that included several known compounds such as derivatives of the polyketide alternariol (21) as well as two red coloured new natural products named xanalteric acid I and II (22, 23) that featured five highly conjugated ring systems. Compounds like xanalteric acids that exhibit a 10H-phenaleno(1,2,3-de)chromene skeleton have so far not been isolated from fungi. A similar compound, xanosporic acid (24), was previously obtained as a bacterial degradation product of the fungal constituent cercosporin (25) when the latter was added to the fermentation broth of Xanthomonas campestris pv. zinniae (Mitchell et al. 2002, 2003). Thus, it seemed possible that the xanalteric acids isolated in this study could likewise represent bacterial biotransformation products of unknown fungal precursors. It is known that some fungi harbour endosymbiotic bacteria as recently reported for the fungus Rhizopus sp. that is colonized by Burkholderia sp. (Partida-Martinez and Hertweck 2005). However, this hypothesis could be discarded in our case as treatment of the fungal culture with antibiotics had no influence on production of xanalteric acids. Furthermore, PCR experiments using eubacterial primers showed no indication for bacterial DNA in the culture of Alternaria sp. (Kjer et al. 2009). Thus, xanalteric acids I and II can be regarded as true fungal metabolites.
The crude extract of Alternaria sp. exhibited pronounced cytotoxic activity against murine L5178Y cells. Screening of the various isolated constituents revealed that alternariol (21) and some alternariol derivatives were mainly responsible for this activity. The EC50 value of alternariol against the cell line investigated was 1.7 μg/ml. As a potential intracellular target of the compound several protein kinases were identified (Aly et al. 2008). When tested against a panel of more than 20 different human protein kinases alternariol (21) was found to inhibit several kinases with EC50 values lower than 1 μg/ml. Kinases that proved to be particularly sensitive to alternariol included Aurora-A and -B as well as FLT3. However, it has been shown recently that alternariol not only inactivates protein kinases but also inhibits Topoisomerases I and II (Fehr et al. 2009) that are also important targets for tumor chemotherapy making this fungal compound an interesting broad spectrum inactivator of tumor cells. Xanalteric acids I and II exhibited only marginal cytotoxic activity in comparison.









Conclusions
Marine natural product research continues to be a fascinating, multidisciplinary field that draws its strength from mutual influences of basic and applied aspects. The importance of marine derived secondary constituents as leads for new drugs has recently been corroborated by admission of Prialt® and Yondelis® to the market. The development of the pain killer Prialt® that represents a neurotoxic peptide from predatory Conus snails used to paralyze fish impressively underlines the importance of ecological observations for a targeted approach to drug discovery. The search for new anti fouling compounds from the sea adopts a similar research strategy. While research on marine macroorganisms will continue in the years to come marine microorganisms are presently emerging as new sources for bioactive leads due to the relative ease of their accessibility and due to sustainable production of their constituents through fermentation. This new focus will help in bridging the supply problem that in the past has slowed the development of macroorganism derived drug candidates.
References
Aly AH, Edrada-Ebel RA, Indriani ID, Wray V, Müller WEG, Totzke F, Zirrgiebel U, Schächtele C, Kubbutat MHG, Lin WH, Proksch P, Ebel R (2008) Cytotoxic metabolites from the fungal endophytic Alternaria sp. and their subsequent detection in its host plant Polygonum senegalense. J Nat Prod 71:972–980
Alzieu C, Sanjuan J, Michel P, Borel M, Dreno JP (1989) Monitoring and assessment of butyltins in Atlantic coastal waters. Mar Poll Bull 20:22–26
Bao S, Ding Y, Deng Z, Proksch P, Lin WH (2007) Rhynocosides A-F, phenolic constituents from the Chinese mangrove plant Bruguiera sexangula var. rhynchopetal. Chem Pharm Bull 55:1175–1180
Bers AV, Wahl M (2004) The influence of natural surface microtopographies on fouling. Biofouling 20:43–51
Blunt JW, Copp BR, Hu WP, Munro MHG, Northcote PT, Prinsep MR (2009) Marine natural products. Nat Prod Rep 26:170–244
Carballo J, Hernandez-Inda Z, Perez P, Garcia-Gravalos M (2002) A comparison between two brine shrimp assays to detect in vitro cytotoxicity in marine natural products. BMC Biotech 2:17–21
Chellappan S, Jasmin C, Basheer SM, Elyas KK, Bhat SG, Chandrasekaran M (2006) Production, purification and partial characterization of a novel protease from marine Engyodontium album BTMFS10 under solid state fermentation. Process Biochem 41:956–961
Ciminiello P, Costantino V, Fattorusso E, Magno S, Mangoni A (1994a) Chemistry of Verongida sponges, II. Constituents of the Caribbean sponge Aplysina fistularis forma fulva. J Nat Prod 57:705–712
Ciminiello P, Fattorusso E, Magno S (1994b) Chemistry of Verongida sponges, III. Constitutents of a Caribbean Verongula sp. J Nat Prod 57:1564–1569
Ciminiello P, Fattorusso E, Magno S (1995) Chemistry of Verongida sponges, IV. Comparison of the secondary metabolite composition of several specimens of Pseudoceratina crassa. J Nat Prod 58:689–696
Ciminiello P, Dell`Aversano C, Fattorusso E, Magno S (1996a) Chemistry of Verongida sponges—VII Bromocompounds from the Caribbean sponge Aplysina archeri. Tetrahedron 52:9863–9868
Ciminiello P, Fattorusso E, Magno S, Pansini M (1996b) Chemistry of Verongida sponges—VI. Comparison of the secondary metabolic composition of Aplysina insularis and Aplysina fulva. Biochem Syst Ecol 24:105–113
Ciminiello P, Fattorusso E, Forino M, Magno S (1997) Chemistry of Verongida sponges VIII—Bromocompounds from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. Tetrahedron 53:6565–6572
Ciminiello P, Dell`Aversano C, Fattorusso E, Magno S, Pansini M (1999) Chemistry of Verongida sponges. 9. Secondary metabolite composition of the Caribbean sponge Aplysina cauliformis. J Nat Prod 62:590–593
Ciminiello P, Dell`Aversano C, Fattorusso E, Magno S, Pansini M (2000) Chemistry of Verongida sponges 10. Secondary metabilite composition of the Caribbean sponge Verongula gigantea. J Nat Prod 63:263–266
Cimino G, De Rosa S, De Stefano S, Self R, Sodano G (1983) The bromo-compounds of the true sponge Verongia aerophoba. Tetrahedron Let 24:3029–3032
Clare AS (1996a) Natural product antifoulants: status and potential. Biofouling 9:211–229
Clare AS (1996b) Signal transduction in barnacle settlement: calcium re-visited. Biofouling 10:141–159
Conn E (1979) Cyanide and cyanogenic glycosides. In: Rosenthal G, Janzen D (eds) Herbivores: their interaction with secondary plant metabolites. Academic Press, New York, pp 387–412
Cui J, Wu J, Deng Z, Proksch P, Lin WH (2007) Xylocarpins A-I, limonoids from the Chinese mangrove plant Xylocarpus granatum. J Nat Prod 70:772–778
Debitus C, Guella G, Mancini I, Waikedre J, Guemas J, Nicolas J, Pietra F (1998) Quinolones from a bacterium and tyrosine metabolites from its host sponge, Suberea creba from the Coral Sea. J Mar Biotech 6:136–141
Ebel R (1998) Wundinduzierte Biotransformation bromierter Alkaloide in Schwämmen der Gattung Aplysina: Biochemische Charakterisierung und ökologische Bedeutung. Dissertation, University of Würzburg
Ebel R, Brenzinger M, Kunze A, Gross H, Proksch P (1997) Wound activation of protoxins in marine sponge Aplysina aerophoba. J Chem Ecol 23:1451–1462
Edrada RA, Heubes M, Brauers G, Wray V, Berg A, Graefe U, Wohlfarth M, Muehlbacher J, Schaumann K, Sudarsono, Bringmann G, Proksch P (2002) Online analysis of xestodecalactones A-C, novel bioactive metabolites from the fungus Penicillium cf. montanense and their subsequent isolation from the sponge Xestospongia exigua. J Nat Prod 65:1598–1604
Encarnación-Dimayuga E, Ramírez M, Luna-Herrera J (2003) Aerothionin, a bromotyrosine derivative with antimycobacterial activity from the marine sponge Aplysina gerardogreeni (Demospongia). Pharmaceut Biol 41:384–387
Erickson H (2007) Evolution of the cytoskeleton. BioEssays 29(7):668–677
Fehr M, Pahlke G, Fritz J, Christensen MO, Boege F, Altemöller M, Podlech J, Marko D (2009) Alternariol acts as a topoisomerase poison, preferentially affecting the IIalpha isoform. Mol Nutr Food Res 53:441–451
Fendert T (2000) Charakterisierung der enzymatischen Abwehrreaktion in Schwämmen der Gattung Aplysina und Isolierung von Bromotyrosinalkaloiden aus Aplysina insularis. Dissertation, University of Würzburg
Fusetani N (2004) Biofouling and antifouling. Nat Prod Rep 21:94–104
Hertiani (2007) Isolation and structure elucidation of bioactive secondary metabolites from Indonesian marine sponges. Dissertation, University of Duesseldorf
Höller U, Wright AD, Matthee GF, König GM, Draeger S, Aust HJ, Schulz B (2000) Fungi from marine sponges: diversity, biological activity and secondary metabolites. Mycol Research 104:1354–1365
IMO (2001) Resolution on early and effective application of the international convention on the control of harmful antifouling systems on ships. Resolution A928 (22) IMO
Jaspars M, Rali T, Laney M, Schatzmann RC, Diaz MC, Schmitz FJ, Pordesimo EO, Crews P (1994) The search for inosine 5’-phosphate dehydrogenase (IMPDH) inhibitors from marine sponges-evaluation of the bastadin alkaloids. Tetrahedron 50:7367–7374
Kelly S, Jensen P, Henkel T, Fenical W, Pawlik J (2003) Effects of Caribbean sponge extracts on bacterial settlement. Aquat Microb Ecol 31:175–182
Kjer J, Wray V, Edrada-Ebel RA, Ebel R, Pretsch A, Lin WH, Proksch P (2009) Xanalteric Acid and related phenolic compounds from endophytic Alternaria sp. isolated from the mangrove plant Sonneratia alba. J Nat Prod 72:2053–2057
Li L, Li X, Shi C, Deng Z, Fu H, Proksch P, Lin WH (2006) Pongamone A-E, five flavonoids from the stems of a mangrove plant, Pongamia pinnata. Phytochemistry 67:1347–1352
Lin W, Brauers G, Wray V, Berg A, Sudarsono ProkschP (2003) Novel chromone derivatives from the fungus Aspergillus versicolor isolated from the marine Sponge Xestospongia exigua. J Nat Prod 66:57–61
Liu HB, Edrada-Ebel RA, Ebel R, Wang Y, Schulz B, Draeger S, Müller WEG, Wray V, Lin WH, Proksch P (2009) Drimane sesquiterpenoids from the fungus Aspergillus ustus isolated from the marine sponge Suberites domuncula. J Nat Prod 72:1585–1588
Mack MM, Molinski TF, Buck ED, Pessah IN (1994) Novel modulators of skeletal-muscle Fkbp12 calciumchannel complex from Ianthella basta—role of Fkbp12 in channel gating. J Biol Chem 269:23236–23249
Marchesi V (1985) Stabilizing infrastructure of cell membranes. Annu Rev Cell Biol 1:531–561
McCarty C (1970) Syn-anti Isomerizations and rearrangements. Interscience Publishers, London
Mitchell TK, Chilton WS, Daub ME (2002) Biodegradation of the polyketide toxin cercosporin. Appl Environ Microbiol 68:4173–4181
Mitchell TK, Alejos-Gonzales F, Gracz HS, Danehower DA, Daub ME, Chilton WS (2003) Xanosporic acid, an intermediate in bacterial degradation of the fungal phototoxin cercosporin. Phytochemistry 62:723–732
Ortlepp S, Sjögren M, Dahlstöm M, Weber H, Ebel R, Edrada-Ebel RA, Thoms C, Schupp P, Bohlin L, Proksch P (2007) Antifouling activity of bromotyrosine-derived sponge metabolites and synthetic analogues. Mar Biotechnol 9:776–785
Pansini M (1997) Effects of light on the morphology, distribution and ecology of some Mediterranean sponges. Biol Mar Mediterr 4:74–80
Partida-Martinez LP, Hertweck C (2005) Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 437:884–888
Paul VJ, Ritson-Williams R (2008) Marine chemical ecology. Nat Prod Rep 25:662–695
Paul V, Van Alstyne K (1988) Chemical defense and chemical variation in some tropical Pacific species of Halimeda (Halimedaceae; Chlorophyta). Coral Reefs 6:263–269
Paul V, Van Alstyne K (1992) Activation of chemical defenses in the tropical green algae Halimeda spp. J Exp Mar Biol Ecol 160:191–203
Pawlik J, Chanas B, Toonen R, Fenical W (1995) Defenses of Caribbean sponges against predatory reef fish. I. Chemical deterrency. Mar Ecol Prog Ser 127:183–194
Pettit GR, Butler MS, Williams MD, Filiatrault MJ, Pettit RK (1996) Isolation and structure of hemibastadinols 1–3 from the Papua New Guinea marine sponge Ianthella basta. J Nat Prod 59:927–934
Proksch P, Ebel R, Edrada-Ebel RA, Riebe F, Liu H, Diesel A, Bayer M, Li X, Lin WH, Grebenyuk V, Müller WEG, Draeger S, Zuccaro A, Schulz B (2008) Sponge-associated fungi and their bioactive compounds: the Suberites case. Botanica Marina 51:209–218
Riedl R (1983) Fauna und Flora des Mittelmeeres. Verlag Paul Parey, Hamburg
Rittschof D, Maki J, Mitchell R, Costlow JD (1986) Ion and neuropharmacological studies of barnacle settlement. Neth J Sea Res 20:269–275
Teeyapant R (1994) Brominated secondary metabolites of the marine sponge Verongia aerophoba (Schmidt) and the sponge feeding gastropod Tylodina perversa (Gmelin): Identification, biological activities and biotransformation. Dissertation, University of Würzburg
Teeyapant R, Proksch P (1993) Biotransformation of brominated compounds in the marine sponge Verongia aerophoba—evidence for an induced chemical defense? Naturwissenschaften 80:369–370
Teeyapant R, Kreis P, Wray V, Witte L, Proksch P (1993a) Brominated secondary compounds from the marine sponge Verongia aerophoba and the sponge feeding gastropod Tylodina perversa. Z Naturforsch [C] 48:640–644
Teeyapant R, Woerdenbag HJ, Kreis P, Hacker J, Wray V, Proksch P (1993b) Antibiotic and cytotoxic activity of brominated compounds from the marine sponge Verongia aerophoba. Z Naturforsch [C] 48:939–945
Teuscher F, Lin WH, Wray V, Edrada RA, Padmakumar K, Proksch P, Ebel R (2006) Two new cyclopentanoids from the endophytic fungus Aspergillus sydowii associated with the marine alga Acanthophora spicifera. Nat Prod Com 1:927–933
Thompson JE, Walker ND, Faulkner DJ (1985) Screening and bioassays for biologically active substances from forty marine sponge species from San Diego, California, USA. Mar Biol 88:11–21
Thoms C, Schupp PJ (2008) Activated chemical defense in marine sponges—a case study on Aplysinella rhax. J Chem Ecol 34:1242–1252
Thoms C, Ebel R, Hentschel U, Proksch P (2003a) Sequestration of dietary alkaloids by the spongivorous marine mollusc Tylodina perversa. Z Naturforsch [C] 58:426–432
Thoms C, Horn M, Wagner M, Hentschel U, Proksch P (2003b) Monitoring microbial diversity and natural product profiles of the sponge Aplysina cavernicola following transplantation. Mar Biol 142:685–692
Thoms C, Wolff M, Padmakumar K, Ebel R, Proksch P (2004) Chemical defense of Mediterranean sponges Aplysina cavernicola and Aplysina aerophoba. Z Naturforsch [C] 59:113–122
Thoms C, Ebel R, Proksch P (2006) Activated chemical defense in Aplysina sponges revisited. J Chem Ecol 32:97–123
Wajant H, Effenberger F (1996) Hydroxynitrile lyases of higher plants. Biol Chem 377:611–617
Walker R, Thompson J, Faulkner D (1985) Exudation of biologically-active metabolites in the sponge Aplysina fistularis II. Chemical evidence. Mar Biol 88:27–32
Weiss B, Ebel R, Elbrächter M, Kirchner M, Proksch P (1996) Defense metabolites from the marine sponge Verongia aerophoba. Biochem Syst Ecol 24:1–12
Wilkinson C, Vacelet J (1979) Transplantation of marine sponges to different conditions of light and current. J Exp Mar Biol Ecol 37:91–104
Xu J, Kjer J, Sendker J, Wray V, Guan H, Edrada RA, Lin WH, Wu J, Proksch P (2009a) Chromones from the endophytic fungus Pestalotiopsis sp. isolated from the Chinese mangrove plant Rhizophora mucronata. J Nat Prod 72:662–665
Xu J, Kjer J, Sendker J, Wray V, Guan H, Edrada RA, Müller WEG, Bayer M, Lin WH, Wu J, Proksch P (2009b) Cytosporones, coumarins, and an alkaloid from the endophytic fungus Pestalotiopsis sp. isolated from the Chinese mangrove plant Rhizophora mucronata. Bioorg Med Chem 17:7362–7367
Yang Y, Deng Z, Proksch P, Lin WH (2006) Two new 18-en-oleane derivatives from marine mangrove plant, Barringtonia racemosa. Pharmazie 61:365–366
Zuccaro A, Schulz B, Mitchell JI (2003) Molecular detection of ascomycetes associated with Fucus serratus. Mycol Res 107:1451–1466
Acknowledgments
We are indebted for continued financial support to the BMBF and to Drs. L. Schmitt and N. Hanekop (both HHU Düsseldorf) for their continued help regarding enzyme purification.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Proksch, P., Putz, A., Ortlepp, S. et al. Bioactive natural products from marine sponges and fungal endophytes. Phytochem Rev 9, 475–489 (2010). https://doi.org/10.1007/s11101-010-9178-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11101-010-9178-9