
ABSTRACT
Purpose
We have demonstrated that (-)-gossypol-enriched cottonseed oil [(-)-GPCSO] can down-regulate Bcl-2 expression in MCF-7 and primary cultured human breast cancer epithelial cells (PCHBCECs). However, this agent has not been evaluated in vivo due to its limited solubility. We aimed to develop liposomes containing (-)-GPCSO to suppress Bcl-2/Bcl-xL expression.
Methods
(-)-GPCSO liposomes were prepared and evaluated for effects on breast cancer cell viability, MDA-MB-231 xenograft tumor growth, cellular Bcl-2 and Bcl-xL mRNA levels, and chemosensitivity to paclitaxel.
Results
(-)-GPCSO liposomes prepared had excellent stability. Cytotoxicity of (-)-GPCSO liposomes was significantly reduced compared to (-)-GPCSO in culture medium. Bcl-2 and Bcl-xL mRNA expression was down-regulated by (-)-GPCSO in culture medium or (-)-GPCSO liposomes in MDA-MB-231 cells. In PCHBCECs, Bcl-2 and Bcl-xL expression were down-regulated by (-)-GPCSO liposomes. (-)-GPCSO in culture medium induced only a mild reduction in Bcl-xL. In the MDA-MB-231 xenograft tumor model, (-)-GPCSO liposomes exhibited tumor-suppressive activity and significantly reduced intratumoral Bcl-2 and Bcl-xL expression. Cytotoxicity of paclitaxel was increased by pretreatment with (-)-GPCSO liposomes in MDA-MB-231 and PCHBCECs.
Conclusions
Findings suggest that (-)-GPCSO liposomes warrant continued investigation as a chemosensitizer for breast cancers exhibiting Bcl-2-/Bcl-xL-mediated drug resistance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Breast cancer is the most commonly diagnosed malignancy in women in the United States. About one in every eight women is affected during their lifetime (1). It is the second leading cause of cancer death for women (2). Chemotherapy is the mainstay of treatment for breast cancer patients who are resistant to hormone or human epidermal growth factor receptor 2 (HER2)-targeted therapy (3,4). However, despite the fact that tumors initially respond to chemotherapy, tumor cells can subsequently survive and gain resistance to these treatments. Anti-apoptotic proteins Bcl-2 and Bcl-xL play an important role in the resistance of breast cancer cells to anticancer therapies. Since Bcl-2 and Bcl-xL are frequently expressed in breast cancer cells, down-regulation of Bcl-2 and Bcl-xL can potentially restore chemosensitivity in breast cancer cells. Thus, inhibiting the anti-apoptotic Bcl-2 and Bcl-xL proteins is considered a promising strategy for the treatment of breast cancer. This is supported by the preclinical findings on an antisense oligonucleotide against Bcl-2, G3139 (oblimersen) (5). However, phase I/II clinical studies on G3139 (Bcl-2 antisense oligonucleotide) in combination with doxorubicin and docetaxel in breast cancer showed very little down-regulation of Bcl-2 in primary tumors, perhaps related to issues with insufficient drug delivery (6). Therefore, targeting Bcl-2 and bcl-xL with small-molecule antagonists is a promising strategy for breast cancer therapy.
Cottonseed oil is extracted from the glanded seeds of cotton family (gossypium) and contains gossypol (GP) in small quantities. Naturally occurring GP is a racemic mixture of two optical isomers, (+)-enantiomer and (-)-enantiomer, and has been shown to possess anti-neoplastic activity against a variety of malignant cell types both in vitro and in vivo (7–11). Interestingly, the (-)-enantiomer of GP, (-)-GP, has greater in vitro anticancer activity than the (+)-enantiomer or racemic GP (12–14). Our preliminary study has shown that (-)-GP-enriched cottonseed oil [(-)-GPCSO], which is specifically developed by the USDA, containing 65% (-)-GP and 35% (+)-GP (equivalent to 10.6 and 5.6 μM, respectively), possessed more potent anticancer activity in human breast cancer cells than conventional cottonseed oil, which contains 35% (-)-GP and 65% (+)-GP. Moreover, we have also reported the anticancer activity of (-)-GPCSO against human breast cancer cells and demonstrated that these effects may be attributed to the suppression of anti-apoptotic protein Bcl-2 expression (15). Furthermore, (-)-GPCSO showed more potent Bcl-2 down-regulation activity than (-)-GP at the same concentration of (-)-GP (15). Therefore, (-)-GPCSO may be more practicable for development as a chemosensitizing agent to overcome chemoresistance. However, the activity of Bcl-2 down-regulation has not been evaluated in vivo due to the limited solubility. Even in vitro Bcl-2 down-regulation was also carried out in the presence of dimethyl sulfoxide (DMSO) (15). According to the United States Pharmacopoeia and National Formulary (USP-NF), a drug is insoluble or practically insoluble if its solubility is less than 0.1 mg/ml. Our preliminary data showed that the solubility of (-)-GPCSO in DMSO and ethanol was about 10 and 6 mg/ml, respectively. In water, the solubility of (-)-GPCSO was less than 0.1 mg/ml. Moreover, our unpublished data showed that (-)-GP, a major active component of (-)-GPCSO, was slightly inhibited MDA-MB-231 xenograft tumor growth in vivo at dose of 5 mg/kg. Considered with animal dose and solubility, a delivery vehicle will be needed for efficient delivery to tumor cells and continued development as an anticancer therapeutic agent.
Liposomes are biodegradable and essentially non-toxic and have been used effectively as a carrier of anticancer drugs (16–22). Previous studies have shown that liposome formulation can improve the therapeutic efficacy of anticancer drugs and reduce or modulate their toxicity profiles (16). Liposomes can also improve solubility, prolong blood circulation time and enhance therapeutic activity (20, 21). In addition, we have described liposomal formulation for the delivery of GP, which has low aqueous solubility (23).
In the present study, liposomes containing (-)-GPCSO were prepared and evaluated for Bcl-2 and Bcl-xL downregulation activities in in vitro and in vivo models of breast cancer. Anti-tumor activity of (-)-GPCSO liposomes was evaluated in MDA-MB-231 xenograft tumor model. Moreover, chemosensitization activity to paclitaxel was evaluated by pretreatment of the cancer cells by (-)-GPCSO liposomes.
MATERIALS AND METHODS
Reagents
(-)-GPCSO was provided by Dr. Michael Dowd of the USDA Southern Regional Research Center (New Orleans, LA, USA). Egg phosphatidylcholine (EPC), cholesterol, methoxy-polyethylene glycol (MW = 2000) distearoyl phosphatidylethanolamine (mPEG-DSPE) were purchased from Avanti Polar Lipids (Alabaster, AL, USA). All other chemicals were of analytical grade.
Cell Culture
Human breast cancer tissue was procured from the Tissue Procurement Program of The Ohio State University Comprehensive Cancer Center. PCHBCECs were isolated from these tissues according to the method described previously (14) and cultured in 25 cm2 flasks in low calcium (0.04 mM CaCl2) Dulbecco’s modified Eagle’s medium and Ham’s F12 medium (1:1) mixture (Atlanta Biologicals, Norcross, GA, USA) supplemented with 10% Chelex-100 (Bio-Rad Laboratories, Richmond, CA, USA)-treated fetal bovine serum (GIBCO, Grand Island, NY, USA), 10 μg/ml insulin, 5 μg/ml hydrocortisone and 20 μg/ml epidermal growth factor (Calbiochem, San Diego, CA, USA), plus 100 ng/ml cholera toxin (Sigma-Aldrich, St Louis, MO, USA). MDA-MB-231 cells were purchased from the American Type Culture Collection (ATCC, Rockville, MD, USA) and cultured in DMEM containing 10% FBS.
Preparation of Liposomes Containing (-)-GPCSO
Liposomes containing (-)-GPCSO were prepared by polycarbonate membrane extrusion methods. The lipid composition was EPC/Cholesterol/mPEG-DSPE at molar ratios of 65:30:5. Briefly, lipids and (-)-GPCSO were dissolved in chloroform and dried in a round-bottom flask on a rotary evaporator under vacuum. The resulting lipid film was hydrated with PBS (pH 7.4). The lipid suspension was then serially passed through 0.2, 0.1 and 0.05 μm pore size polycarbonate membranes on a nitrogen-driven Lipex™ extruder (Northern Lipid Inc., Canada). To determine (-)-GPCSO concentration, liposomes were lysed with ethanol, and the concentration in the lysate was determined by measuring absorption at 371 nm, on a Shimadzu UV–vis spectrophotometer. Size distribution of (-)-GPCSO liposomes was determined by dynamic light scattering on a NICOMP 370 Submicron Particle Sizer (NICOMP, Santa Barbara, CA). The final product was stored at 4°C and evaluated for colloidal stability by monitoring changes in the mean particle diameter.
Cryogenic Transmission Electron Microscopy (Cryo-TEM)
The morphology of (-)-GPCSO liposomes was examined by cryo-TEM. Samples for cryo-TEM were prepared as described previously (24). Briefly, a drop of (-)-GPCSO liposomes was applied to a perforated carbon film, supported by a copper grid and held by the controlled environment vitrification system. Then, the grid was immediately plunged into liquid ethane at its melting point (−183°C) and stored in liquid nitrogen (−196°C). The vitrified sample was examined in a Tecnai G2 Transmission Electron Microscope (FEI Company, Oregon, USA) operated at 120 kV using a Gatan HC3500 Tilt heating/Nitrogen cooling holder (Pleasanton, CA, USA). Digital images were captured by a Gatan 791 MultiScan CCD camera and processed using the Digital Micrograph 3.1 software package.
Cytotoxicity of Liposomes Containing (-)-GPCSO Against Breast Cancer Cells
Cytotoxicity of liposomes containing (-)-GPCSO was determined in MDA-MB-231 and PCHBCECs using the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide] assay. Cells were seeded into 96-well plates at 3,000 and 5,000 cells/well for MDA-MB-231 and PCHBCECs, respectively. After overnight incubation, cells were treated for 48 h at 37°C with (-)-GPCSO in culture medium or (-)-GPCSO-containing liposomes prepared by 4-fold serial dilutions in culture medium to generate final concentrations of (-)-GPCSO ranging from 0.2 to 1,000 μg/ml. For the chemosensitization experiments, cells were cultured as described above and then treated with 200 μg/ml (-)-GPCSO liposomes for 24 h, followed by exposure to various concentrations of paclitaxel (0.0001–10 μM) for an additional 24 h at 37°C. After treatments, 20 μL of 5 mg/ml MTT was added to each well and incubated for 4 h at of 37°C. The formazan crystals were then dissolved in 200 μL of DMSO, and the absorbances were determined at 570 nm on a Dynatech MR-600 microplate reader (Dynatech Laboratories, Chantilly, VA, USA).
Effect of Liposomes Containing (-)-GPCSO on mRNA Expression Levels of Bcl-2 Family Members in Breast Cancer Cells
MDA-MB-231 and PHBCECs were seeded into 6-well plates at 5 × 105/well and cultured overnight. Cells were treated with different concentrations of (-)-GPCSO in culture medium or (-)-GPCSO liposomes for 48 h. (-)-GPCSO in culture medium was prepared for treatment by dissolving in DMSO and then diluting with PBS. The final DMSO concentration was less than 2%. After incubation, cells were collected, and RNA was isolated for analysis of mRNA levels by real-time quantitative RT–PCR.
RNA Isolation and Real-Time Quantitative RT-PCR
The Bcl-2 and Bcl-xL mRNA levels in (-)-GPCSO-treated MDA-MB-231 cells and PCHBCECs were evaluated using real-time RT-PCR. Briefly, total RNA was extracted using Trizol reagent (Invitrogen, Carlsbad, CA, USA), and cDNA was synthesized by incubating RNA with random hexamer primer and dNTPs, and then with M-MLV reverse transcriptase (Invitrogen, Carlsbad, CA, USA) in the presence of first-strand buffer, dithiothreitol, and RNAase inhibitor, followed by incubation at 37°C for 50 min and 75°C for 15 min in a gradient mastercycler (Eppendorf, Westbury, NY, USA). The resulting cDNA was amplified by real-time PCR using SYBR green PCR Mastermix (Applied Biosystems, Foster City, CA, USA) and specific primers. For Bcl-2, forward primer (CACACCTGGATCCAGGATAAC) and reverse primer (AGACAGCCAGGAGAAATCAAAC) were used. For Bcl-xL, forward primer (TTACCTGAATGACCACCTA) and reverse primer (ATTTCCGACTGAAGAGTGA) were used. The mRNA level of the housekeeping gene 36B4 was also analyzed concurrently, and all data were normalized to the 36B4 value.
In Vivo Anticancer Activity of Liposomes Containing (-)-GPCSO in MDA-MB-231 Xenograft Tumor Model
In vivo anticancer activity of liposomes containing (-)-GPCSO were evaluated in MDA-MB-231 tumor-bearing athymic nude mice (Charles River, Wilmington, MA, USA). Tumors were established in the right flank of each mouse (18–22 g) by subcutaneous injection of 4 × 106 MDA-MB-231 cells suspended in 0.1 ml of vehicle containing equal volumes of culture medium and Matrigel (Becton Dickinson Labware, Bedford, MA, USA). When tumors reached a volume of 50–100 mm3, mice were randomly assigned to two groups (5 mice/group). The treatment group received liposomes containing (-)-GPCSO at 20 mg/kg on days 1, 5, 9 and 13 via intravenous injection into the tail vein. The control group was treated with blank liposomes. Tumor dimensions were measured with calipers every 4 days after the initiation of treatments, and tumor volume (mm3) was calculated using the formula: \( 0.{5} \times {\text{length}} \times {\text{width}} \times {\text{height}} \). Twenty-four hours after the last treatment, the mice were euthanized by CO2 inhalation, tumors were collected, and portions were snap-frozen for subsequent analysis of mRNA expression levels or fixed in 10% formalin for histopathological evaluation of hematoxylin & eosin-stained sections.
Statistical Analysis
Data are presented as means ± standard deviations and were analyzed by two-tailed Student’s t-test using MiniTAB Program (Minitab Inc., State College, PA). Differences between group means were considered statistically significant at P < 0.05.
RESULTS
Preparation and Characterization of Liposomes Containing (-)-GPCSO
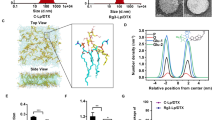
Liposomes containing (-)-GPCSO were prepared by the polycarbonate membrane extrusion method. The resulting liposomes had an average particle size of 73.2 ± 2.4 nm and a zeta potential of −12.6 ± 1.10 mV. The (-)-GPCSO encapsulation efficiency was found to be over 95% at the (-)-GPCSO-to-lipid ratio of 0.1:1 and 0.2:1 (wt/wt). However, the (-)-GPCSO encapsulation efficiency was decreased to 70% with increasing the (-)-GPCSO-to-lipid ratio to 0.3:1. Therefore, the (-)-GPCSO-to-lipid ratio of 0.2:1 was selected. The final (-)-GPCSO concentration in the liposomes was adjusted to 5 mg/ml. The cryo-TEM results showed the (-)-GPCSO-containing liposomes to have a uni- or bilamellar structure (Fig. 1a). The stability of liposomes containing (-)-GPCSO was determined by examining changes in the particle size over time. As shown in Fig. 1b, no change in particle size was observed over the course of 3 months of storage at 4°C.

Characterization of liposomes containing (-)-GPCSO. (a) Cryogenic transmission electron micrographs of liposomes containing (-)-GPCSO. Black and white arrows indicate uni- and bi-lamellar structures, respectively. (b) Stability of liposomes containing (-)-GPCSO. Liposomes containing (-)-GPCSO were stored at 4°C and evaluated for colloidal stability by monitoring changes in the mean particle diameter over 90 days. Data represents the mean ± SD (n = 4).
In Vitro Cytotoxicity of Liposomes Containing (-)-GPCSO in MDA-MB-231 and PCHBCECs
The cytotoxic effects of liposomes containing (-)-GPCSO on MDA-MB-231 and PCHBCECs were determined by MTT assay. As shown in Fig. 2, (-)-GPCSO liposomes were less cytotoxic to these cancer cells compared with (-)-GPCSO in culture medium. The IC50 values of (-)-GPCSO in MDA-MB-231 cells and PCHBCECs from two different patients were 217.2 ± 30.5, 137.8 ± 29.8 and 247.3 ± 24.5 μg/ml, respectively. In contrast, the IC50 value of liposomes containing (-)-GPCSO was >1,000 μg/ml in all three breast cancer cell types. At 1,000 μg/ml concentrations, cell viabilities after treatment with (-)-GPCSO liposomes were 73.0, 80.9 and 70.1% in MDA-MB-231 cells and the two patient-derived PCHBCECs, respectively. These results demonstrate that the cytotoxicity of (-)-GPCSO was decreased by liposomes containing (-)-GPCSO.

In vitro cytotoxicity of liposomes containing (-)-GPCSO in MDA-MB-231 cells (a) and PCHBCECs from Patient 1 (b) and Patient 2 (c). 3000 (MDA-MB-231) and 5000 (PCHBCEC) cells were seeded into each well of 96-well plates and exposed to different concentrations of (-)-GPCSO or liposomes containing (-)-GPCSO for 48 h at 37°C. Cell viability was assessed by MTT assay. Data represent the mean ± SD (n = 4).
Bcl-2 and Bcl-xL Expression in PCHBCECs
Over-expression of Bcl-2 or Bcl-xL is known to inhibit chemotherapy-induced apoptosis (25–27). Therefore, Bcl-2 and Bcl-xL mRNA expression levels in PCHBCECs from five different patient tumor samples and two different breast cancer cell lines were determined by RT-PCR (Fig. 3a). As shown, both Bcl-2 and Bcl-xL mRNA were expressed in MDA-MB-231 cells, while MCF-7 cells expressed only Bcl-2. All five of the patient-derived PCHBCECs expressed both Bcl-2 and Bcl-xL mRNA; however, the expression levels of Bcl-2 mRNA varied among these samples and were markedly lower than in the cell lines, while the Bcl-xL expression levels were relatively consistent among the samples and were only slightly lower than in the cell lines. In contrast to our findings, Emi et al. reported that Bcl-2 is more frequently expressed than Bcl-xL (70% versus 40%) in breast cancer tissue (5). This discrepancy might reflect differences in gene expression among the epithelial cells, stroma cells and pre-adipocytes that are present in breast tissue. Considering that the great majority of human breast cancers arise from epithelial cells (28), Bcl-2, as well as Bcl-xL, might play important roles in conferring drug resistance.

Effect of (-)-GPCSO and (-)-GPCSO liposomes on Bcl-2 and Bcl-xL mRNA expression levels in breast cancer cells. (a) Expression levels of Bcl-2 and Bcl-xL mRNA in the MCF-7 and MDA-MB-231 breast cancer cell lines and in PCHBCECs isolated from five different patient tumor samples as determined by RT-PCR. (b) Down-regulation of Bcl-2 and Bcl-xL mRNA expression by (-)-GPCSO in culture medium and (-)-GPCSO liposomes in MDA-MB-231 cells and PCHBCECs from two patient tumor samples. Cells were treated with different concentrations of (-)-GPCSO and liposomes containing (-)-GPCSO for 48 h at 37°C and Bcl-2 and Bcl-xL mRNA expression was determined by real-time qRT-PCR. Data represent the mean ± SD (n = 4). *, P < 0.05 compared with control.
Liposomes Containing (-)-GPCSO Down-Regulated Bcl-2 and Bcl-xL mRNA Expression
Previously, we demonstrated that (-)-GPCSO significantly down-regulated Bcl-2 mRNA and protein levels in MCF-7 cells (15). Here, the effect of (-)-GPCSO liposomes versus (-)-GPCSO in culture medium on Bcl-2 and Bcl-xL mRNA expression was investigated in MDA-MB-231 cells and two patient-derived PCHBCECs (Fig. 3b). In MDA-MB-231 cells, the mRNA levels of both Bcl-2 and Bcl-xL were down-regulated in a concentration-dependent manner by treatment with either (-)-GPCSO in culture medium or (-)-GPCSO liposomes. In PCHBCECs, however, only (-)-GPCSO liposomes were able to dose-dependently suppress both Bcl-2 and Bcl-xL mRNA levels. (-)-GPCSO in culture medium did not alter Bcl-2 mRNA levels and caused only mild reductions in Bcl-xL mRNA expression in these patient-derived cells. The higher concentration of (-)-GPCSO (500 μg/ml) could not be used because of cytotoxicity. The reason for this discrepancy between breast cancer cell lines and PCHBCECs in their sensitivity to (-)-GPCSO-induced down-regulation of Bcl-2 is unclear, but may be related to the low basal levels of Bcl-2 expression in the primary cultured cells used in this study. Nonetheless, these results suggest that (-)-GPCSO liposomes possesses stronger anti-Bcl-2 and Bcl-xL activity than (-)-GPCSO.
Liposomes Containing (-)-GPCSO Suppressed MDA-MB-231 Xenograft Tumor Growth
To examine the anti-cancer activity of liposomes containing (-)-GPCSO, MDA-MB-231 tumor-bearing athymic nude mice were treated four times with (-)-GPCSO liposomes at 20 mg/kg every 4 days by intravenous injection. The tumor volume was significantly inhibited by approximately 27% at 12 days after the start of treatment compared to the empty liposome-treated control group (Fig. 4a). All animals tolerated the treatments well with no observable signs of toxicity and no change in body weight over the course of the study (data not shown). Consistent with our in vitro data, the intratumoral expression of anti-apoptotic Bcl-2 and Bcl-xL was significantly downregulated by treatment with (-)-GPCSO liposomes (Fig. 4b). Histopathological examination of MDA-MB-231 tumors from liposomes containing (-)-GPCSO-treated mice showed significant necrosis compared with those from control mice (Fig. 4b), suggesting that necrosis may account for part of the antitumor activity of liposomes containing (-)-GPCSO.

In vivo activity of liposomes containing (-)-GPCSO in the MDA-MB-231 breast cancer xenograft tumor model. Mice with established subcutaneous MDA-MB-231 tumors were treated intravenously every four days for a total of four treatments with liposomes containing (-)-GPCSO at a dose of 20 mg/kg. Control mice received empty liposomes. (a) Tumor growth curves for treated and control groups. Data are presented as means ± SD (n = 5). (b) Intratumoral mRNA levels of Bcl-2 and Bcl-xL were determined by real-time qRT-PCR. Expression levels of the targets were normalized to that of the housekeeping gene, 36B4. (c) Photomicrographs of H&E-stained sections of MDA-MB-231 xenograft tumors from control (left panel) and liposomes containing (-)-GPCSO-treated (right panel) mice (magnification, x 400; inserted magnification, x 40). *, P < 0.05 compared with control.
Liposomes Containing (-)-GPCSO Improved Chemosensitivity of Breast Cancer Cells to Paclitaxel
The expression of the Bcl-2 and/or Bcl-xL protein confers resistance to chemotherapy-mediated apoptotic signals in patients with breast cancer (25–27). A preclinical study in breast cancer has demonstrated that combining anti-Bcl-2 therapy with cytotoxic agents, such as paclitaxel, leads to enhanced antitumor effects (5). Therefore, we sought to demonstrate that liposomal delivery of (-)-GPCSO results in an increased sensitivity to paclitaxel in MDA-MB-231 and PCHBCECs. Cells were treated with liposomes containing (-)-GPCSO or (-)-GPCSO in culture medium, followed by exposure to various concentrations of paclitaxel. As shown in Fig. 5a, liposomes containing (-)-GPCSO at 200 μg/ml could sensitize MDA-MB-231 cells to paclitaxel, as evidenced by a 3.3-fold decrease in paclitaxel’s IC50 from 8.70 μM to 2.65 μM. In contrast, (-)-GPCSO in culture medium could not sensitize MDA-MB-231 cells to paclitaxel at 200 μg/ml concentrations. Similar results were also observed in PCHBCECs. Liposomes containing (-)-GPCSO at 200 μg/ml decreased paclitaxel’s IC50 5-fold, from 1.34 μM to 0.24 μM (Fig. 5b).

Chemosensitization of MDA-MB-231(a) and PCHBCECs (b) to paclitaxel by pre-treatment with liposomes containing (-)-GPCSO. Cells were pre-treated with buffer (control), (-)-GPCSO in culture medium or (-)-GPCSO liposomes at 200 μg/ml for 24 h, after which the cells were treated with paclitaxel at various concentrations (0.0006–10 μM) for 24 h. Viability was then determined by MTT assays. Data represent the mean ± SD (n = 4).
DISCUSSION
The aim of our work is to develop liposomal formulation of (-)-GPCSO to suppress antiapoptotic Bcl-2 and Bcl-xL expression. Therefore, liposomes containintg (-)-GPCSO were prepared and assessed for its ability to inhibit the mRNA expression levels of Bcl-2 and Bcl-xL in MDA-MB-231 and PCHBCECs and to suppress xenograft tumor growth in vivo. The results showed that the solubility of (-)-GPCSO was significantly increased by liposome encapsulation. The particle size of liposomal formulation was decreased by encapsulation (-)-GPCSO. The stability of liposomes containing (-)-GPCSO was confirmed for up to 3 months of storage at 4°C, (Fig. 1b) without evidence of aggregation and changes in size.
The inhibition of both Bcl-2 and Bcl-xL mRNA expression was observed in MDA-MB-231 cells and PCHBCECs from two patients by treatment of (-)-GPCSO liposomes. Moreover, liposomes containing (-)-GPCSO showed greater anti-Bcl-2 and Bcl-xL activity than (-)-GPCSO in culture medium. Decreased Bcl-2 and Bcl-xL gene expression was also founded in vivo in MDA-MB-231 xenograft tumors from mice treated with (-)-GPCSO liposomes in which Bcl-xL was completely down-regulated compared to Bcl-2 which was reduced by approximately 50%. In light of the numerous studies that have demonstrated the relationship between Bcl-2 and Bcl-xL expression and chemosensitivity in breast cancer cells and, specifically, that inhibition of Bcl-2 and Bcl-xL expression increases sensitivity to doxorubicin, mitomycin C and paclitaxel (5), we assessed the effect of (-)-GPCSO on the sensitivity of MDA-MB-231 and PCHBCECs to paclitaxel. Our findings show that pretreatment of breast cancer cells with (-)-GPCSO liposomes increased chemosensitivity to paclitaxel (Fig. 5). In contrast, (-)-GPCSO in culture medium could not sensitize MDA-MB-231 cells to paclitaxel at 200 μg/ml concentrations. These results suggest that liposomes containing (-)-GPCSO may have value as chemosensitizer in breast cancer cells exhibiting Bcl-2-/Bcl-xL-mediated drug resistance.
As mentioned above, Bcl-2 and Bcl-xL mRNA expression levels were reduced in MDA-MB-231 xenograft tumors from (-)-GPCSO-treated mice, which occurred in association with the inhibition of tumor growth (Fig. 4). Consistent with the inhibition of tumor growth, histopathology of the (-)-GPCSO-treated MDA-MB-231 tumors showed significant necrosis compared with those from the control mice. Intratumoural hypoxia is reflected by the presence of necrosis, which has also been reported to be a prognostic factor in patients with breast and bladder cancers (29). The hypoxia-inducible factor-1α (HIF-1α) plays a key role in cellular response to hypoxia, including the regulation of genes involved in energy metabolism, angiogenesis, and apoptosis. In light of reported HIF-1α dependent Bcl-xL overexpression (30), HIF-1α-mediated Bcl-xL down-regulation might contributed to the anticancer activity of liposomes containing (-)-GPCSO in MDA-MB-231 xenograft tumors.
In summary, liposomes containing (-)-GPCSO significantly down-regulated Bcl-2 and Bcl-xL expression in association with the inhibition of breast cancer cell survival in vitro and tumor growth in vivo. Moreover, (-)-GPCSO liposomes increased the chemosensitivity of breast cancer cells to paclitaxel. Together, these findings indicate that the (-)-GPCSO liposomes may have particular value in breast cancer therapy for overcoming drug resistance mediated by Bcl-2 and Bcl-xL. Also, liposomes containing (−)-GPCSO as delivery vehicle have the potential to be used in the anticancer drug delivery.
REFERENCES
Jemal A, Siegel R, Ward E, Murry T, Xu J, Smigal C, et al. Cancer statistics. CA Cancer J Clin. 2006;56:106–30.
Smigal C, Jemal A, Ward E, Collinides V, Smith R. Trends in breast cancer by race and ethnicity. CA Cancer J Clin. 2006;56:168–83.
Coley HM. Mechanisms and strategies to overcome chemotherapy resistance in metastatic breast cancer. Cancer Treat Rev. 2008;34:378–90.
LaPensee EW, Ben-Jonathan N. Novel roles of prolactin and estrogens in breast cancer: resistance to chemotherapy. Endocr Relat Cancer. 2010;17:R91–107.
Emi M, Kim R, Tanabe K, Uchida Y, Toge T. Targeted therapy against Bcl-2-related proteins in breast cancer cells. Breast Cancer Res. 2005;7:R940–52.
Moulder SL, Symmans WF, Booser DJ, Madden TL, Lipsanen C, Yuan L, et al. Phase I/II study of G3139 (Bcl-2 antisense oligonucleotide) in combination with doxorubicin and docetaxel in breast cancer. Clin Cancer Res. 2008;14:7909–16.
Tuszynski GP, Cossu G. Differential cytotoxic effect of gossypol on human melanoma, colon carcinoma, and other tissue culture cell lines. Cancer Res. 1984;44:768–71.
Rao PN, Wang YC, Lotzova E, Khan AA, Rao SP, Stephens LC. Antitumor effects of gossypol on murine tumors. Cancer Chemother Pharmacol. 1985;15:20–5.
Majumdar SK, Daly EP, Kleemeyer KM, Daehler CC, Baker MA. Genotoxic effects of gossypol acetic acid on cultured murine erythroleukemia cells. Environ Mol Mutagen. 1991;18:212–9.
Gilbert NE, O’Reilly JE, Chang CJ, Lin YC, Brueggemeier RW. Antiproliferative activity of gossypol and gossypolone on human breast cancer cells. Life Sci. 1995;57:61–7.
Wang X, Wang J, Wong SC, Chow LS, Nicholls JM, Wong YC, et al. Cytotoxic effect of gossypol on colon carcinoma cells. Life Sci. 2000;67:2663–71.
Benz CC, Keniry MA, Ford JM, Townsend AJ, Cox FW, Palayoor S, et al. Biochemical correlates of the antitumor and antimitochondrial properties of gossypol enantiomers. Mol Pharmacol. 1990;37:840–7.
Shelley MD, Hartley L, Fish RG, Groundwater P, Morgan JJ, Mort D, et al. Stereo-specific cytotoxic effects of gossypol enantiomers and gossypolone in tumour cell lines. Cancer Lett. 1999;135:171–80.
Liu S, Kulp SK, Sugimoto Y, Jiang J, Chang HL, Dowd MK, et al. The (-)-enantiomer of gossypol possesses higher anticancer potency than racemic gossypol in human breast cancer. Anticancer Res. 2002;22:33–8.
Ye W, Chang HL, Wang LS, Huang YW, Shu S, Sugimoto Y, et al. Induction of apoptosis by (-)-gossypol-enriched cottonseed oil in human breast cancer cells. Int J Mol Med. 2010;26:113–9.
Safra T, Muggia F, Jeffers S, Tsao-Wei DD, Groshen S, Lyass O, et al. Pegylated liposomal doxorubicin (Doxil): reduced clinical cardiotoxicity in patients reaching or exceeding cumulative doses of 500 mg/m2. Ann Oncol. 2000;11:1029–33.
Gibbs WJ, Drew RH, Perfect JR. Liposomal amphotericin B: clinical experience and perspectives. Expert Rev Anti-infective Ther. 2005;3:167–81.
Sharma A, Mayhew E, Straubinger RMA. Antitumor effect of paclitaxel-containing liposomes in a paclitaxel-resistant murine tumor model. Cancer Res. 1993;53:5877–81.
Boehlke L, Winter JN. Sphingomyelin/cholesterol liposomal vincristine: a new formulation for an old drug. Expert Opin Biol Ther. 2006;6:409–15.
Medina OP, Zhu Y, Kairemo K. Targeted liposomal drug delivery in cancer. Curr Pharm Des. 2004;10:2981–9.
Cui Y, Wu Z, Liu X, Ni R, Zhu X, Ma L, et al. Preparation, safety, pharmacokinetics, and pharmacodynamics of liposomes containing Brucea javanica oil. AAPS PharmSciTech. 2010;11:878–84.
Pan XQ, Zheng X, Shi G, Wang H, Ratnam M, Lee RJ. Strategy for the treatment of acute myelogenous leukemia based on folate receptor beta-targeted liposomal doxorubicin combined with receptor induction using all-trans retinoic acid. Blood. 2002;100:594–602.
Zhai G, Wu J, Zhao X, Yu B, Li H, Lu Y, et al. A liposomal delivery vehicle for the anticancer agent gossypol. Anticancer Res. 2008;28:2801–5.
Yang X, Koh CG, Liu S, Pan X, Santhanam R, Yu B, et al. Transferrin receptor-targeted lipid nanoparticles for delivery of an antisense oligodeoxyribonucleotide against Bcl-2. Mol Pharm. 2009;6:221–30.
Johnstone RW, Ruefli AA, Lowe SW. Apoptosis: a link between cancer genetics and chemotherapy. Cell. 2002;108:153–64.
Shangary S, Johnson DE. Recent advances in the development of anticancer agents targeting cell death inhibitors in the Bcl-2 protein family. Leukemia. 2003;17:1470–81.
Kirkin V, Joos S, Zornig M. The role of Bcl-2 family members in tumorigenesis. Biochim Biophys Acta. 2004;1644:229–49.
Elenbaas B, Spirio L, Koerner F, Fleming MD, Zimonjic DB, Donaher JL, et al. Human breast cancer cells generated by oncogenic transformation of primary mammary epithelial cells. Genes Dev. 2001;15:50–65.
Hiraoka N, Ino Y, Sekine S, Tsuda H, Shimada K, Kosuge T, et al. Tumour necrosis is a postoperative prognostic marker for pancreatic cancer patients with a high interobserver reproducibility in histological evaluation. Br J Cancer. 2010;103:1057–65.
Chen N, Chen X, Huang R, Zeng H, Gong J, Meng W, et al. BCL-xL is a target gene regulated by hypoxia-inducible factor-1{alpha}. J Biol Chem. 2009;284:10004–12.
ACKNOWLEDGMENTS & DISCLOSURES
This study is supported by NIH Grant R01 ES-015212 and Cotton Inc. Grant 06–864.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Li, H., Piao, L., Xu, P. et al. Liposomes Containing (-)-Gossypol-Enriched Cottonseed Oil Suppress Bcl-2 and Bcl-xL Expression in Breast Cancer Cells. Pharm Res 28, 3256–3264 (2011). https://doi.org/10.1007/s11095-011-0498-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11095-011-0498-2