
Abstract
A significant challenge for understanding alcoholism lies in discovering why some, but not other individuals, become dependent on alcohol. Genetic, environmental, cultural, developmental, and neurobiological influences are recognized as essential factors underlying a person's risk for becoming alcohol dependent (AD); however, the neurobiological processes that trigger this vulnerability are still poorly understood. Hormones are important in the regulation of many functions and several hormones are strongly associated with alcohol use. While medical consequences are important, the primary focus of this review is on the underlying confluence of appetitive/feeding, reproductive and posterior pituitary hormones associated with distinct phases of alcoholism or assessed by alcohol craving in humans. While these hormones are of diverse origin, the involvement with alcoholism by these hormone systems is unmistakable, and demonstrates the complexity of interactions with alcohol and the difficulty of successfully pursuing effective treatments. Whether alcohol associated changes in the activity of certain hormones are the result of alcohol use or are the result of an underlying predisposition for alcoholism, or a combination of both, is currently of great scientific interest. The evidence we present in this review suggests that appetitive hormones may be markers as they appear involved in alcohol dependence and craving, that reproductive hormones provide an example of the consequences of drinking and are affected by alcohol, and that posterior pituitary hormones have potential for being targets for treatment. A better understanding of the nature of these associations may contribute to diagnosing and more comprehensively treating alcoholism. Pharmacotherapies that take advantage of our new understanding of hormones, their receptors, or their potential relationship to craving may shed light on the treatment of this disorder.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Though interest to improve treatments for alcohol dependence (AD) continues to grow, we lack consistently effective treatments to prevent or decrease alcohol use in the alcohol dependent individual (Edwards et al. 2011; Heilig et al. 2011). Advances in neurobehavior have identified neural systems that initiate and sustain alcohol drinking and targeted pharmacologic modification of the neurotransmitters or receptors of these neural systems may alter AD (Kranzler and Edenberg 2010). Animal models often identify pharmacologic agents that reduce alcohol consumption in animals, suggesting similar agents could reduce alcohol use in humans. However, successful translation into safe and effective drugs for human use still seems distant (Kenna et al. 2004; Leggio et al. 2009a). Many factors are widely recognized as essential to underlying a person's risk of becoming AD, however the neurobiological processes that trigger this vulnerability are still poorly understood (Volkow et al. 2012).
Individuals at risk for developing an alcohol use disorder consistently display deficits in response inhibition, cognitive control, emotional regulation, and are often diagnosed with a comorbid psychopathology. Children of alcoholics are also reported to suffer from neuropsychological abnormalities (Tessner and Hill 2010). Animal and human research suggests that alcohol targets cognitive processes by selectively interacting with brain circuits and neurotransmitter systems (Sullivan 2007). Alcohol affects neurobehavioral functions differently during different phases of life and these functions can also vary with age, gender, nutritional status and family history of alcoholism (Oscar-Berman and Marinković 2007). While connections within the limbic forebrain and distant parts of cognitive processing are “hard-wired,” communication also occurs through hormones.
The science of neuroendocrinology has grown since the early 20th century and more recent advances have led to the discovery and understanding of a greater variety and function of hormones. Additionally, methods for detection of hormones have made endocrine research in humans more accessible (Bellem et al. 2011; Salvante et al. 2012). Changes in hormone levels have long been associated with alcohol use (Vassaf and Hall 1946) as well as potential treatments for alcoholism (Smith 1950). For hormones to function properly, the timing and amount of their release must be synchronized, and the target tissues must be able to respond to them accurately. Alcohol can impair the functions of the hormone-releasing glands and of the target tissues, thereby causing serious medical consequences. Chronic drinking may promote changes in hormone levels that may in turn significantly affect nutritional status leading to malnutrition (Nicolás et al. 2001). Yet, while genetics research is often at the forefront of the discussion of predisposition to alcoholism and other psychopathologies (Heilig et al. 2011; Kenna et al. 2012), hormones are also essential to this discussion as fundamental causative mechanisms associated with craving, drinking, treatment and links to other medical conditions caused by alcoholism. While the evidence is strong that alcohol use causes significant medical problems (Gordon and Lieber 1992), the direction of causation between neuroendocrine changes that occur as a result of alcoholism or predispose one to becoming AD are not yet fully known and are the subject of intense research.
This review will focus primarily on human studies and on three classes of hormones: feeding/appetitive hormones (leptin, ghrelin, growth-hormone: GH, insulin, insulin-like growth factor 1 (IGF-1), adiponectin, thyroid hormones); reproductive hormones (prolactin, luteinizing hormone: LH, follicle stimulating hormone: FSH, testosterone and estrogens); atrial natriuretic peptide (ANP), and the posterior pituitary hormones (vasopressin and oxytocin). This review will not discuss research that has focused on other central hormones (e.g. the proopioidmelanocortin (POMC) system, including adrenal hormones). While undoubtedly these hormones are essential contributors to underlying mechanisms for craving and alcohol consumption, they have been covered elsewhere (e.g. Ciccocioppo et al. 2009; Koob 2009). Rarely though, have reviews highlighted research on the lesser known hormonal mechanisms contributing to alcoholism in human studies. The evidence we present suggests in general that appetitive hormones are potentially markers that are involved with alcohol dependence and craving; that reproductive hormones seem affected by alcohol dependence, providing an example of the consequences of alcohol; and posterior hormones have the potential for being targeted for treatment. While the hormones are of diverse origin, the involvement with alcoholism by these hormone systems is unmistakable, and truly demonstrates how complex the interactions with alcohol are and the difficulty of successfully pursuing effective treatments is. The following review demonstrates how far research has come in this quest, yet how far we also have to go.
Appetitive Hormones
There is converging evidence that alcohol and food-seeking behaviors share common neural pathways (Volkow and Wise 2005). For example, individuals with eating disorders such as anorexia (23 % lifetime prevalence) and nervosa bulimia nervosa (55 % lifetime prevalence) have a higher prevalence of alcohol (Bulik et al. 2004) and/or other drug problems (Franko et al. 2008). Moreover in a study of 672 women, alcohol use disorders were significantly more prevalent in women with anorexia nervosa with binge eating and bulimia nervosa than in women with just anorexia nervosa (Bulik et al. 2004). Many of the neurotransmitters and hormones that are reported to regulate appetite also have an influence on alcohol consumption (Leggio 2009; Hillemacher et al. 2010). Consistent with these findings, as detailed next, clinical investigations have been conducted testing feeding-related hormones in alcoholism.
Leptin. Leptin is a neuropeptide secreted by white adipocytes and acts as a key regulator of food intake and energy expenditure, thus playing an important role in hypothalamic appetite regulation (Inui 1999). Leptin also regulates the hypothalamic–pituitary–adrenal (HPA) axis, which is involved in the neurobiology of alcohol craving (Adinoff et al. 2005), and several studies suggest a role of leptin in the neurobiology of alcohol craving. For example, leptin may enhance motivation for alcohol consumption in habituated mice after alcohol withdrawal (Kiefer et al. 2001a). Similarly, in AD subjects, plasma leptin is significantly higher than in healthy controls, and correlates with self-rated craving and decreases during withdrawal (Kiefer et al. 2001b). Furthermore, Kiefer et al. (2005) found an association between leptin plasma levels and alcohol relapse. The difference in leptin plasma concentrations between week 4 and baseline was inversely correlated with abstinence duration. Increasing leptin was associated with early relapse whereas decreasing leptin was associated with longer abstinence duration. In the same study, leptin levels decreased significantly in patients undergoing pharmacological treatment with naltrexone and acamprosate. It should also be noted that other studies focused on the relationship between leptin and nutritional parameters did not find short-term changes of leptin levels in AD subjects (Santolaria et al. 2003). Other research demonstrates an association of leptin and alcohol craving during withdrawal in women (Kraus et al. 2004), and a larger study of 189 men and women AD patients found a positive correlation between leptin serum levels and alcohol craving as measured by the Obsessive Compulsive Drinking Scale (OCDS) in both genders (Hillemacher et al. 2007a). There are several possible explanations why findings were not consistent between studies, including assay variability, gender, differences in the study length and - perhaps the most important - differences in the nutritional status of the samples. In fact, leptin is secreted by white adipocytes, therefore differences in body composition may provide an explanation of different findings among studies. It should be noted, for example, that while most of the studies have corrected leptin values for body mass index (BMI)) levels, patients with similar BMI values, may have differences in the ratio between fat mass and fat-free mass, which might explain differences in leptin levels (Leggio 2009).
Consistent with the increasing knowledge on the importance of subtypes in AD (Leggio et al. 2009a), Hillemacher et al. (2007b) performed a re-analysis of their previous data (Hillemacher et al. 2007a) classifying the subjects according to the Lesch typologies (Lesch et al. 1990). Results showed that leptin levels were highest in Type 4 patients (patients with pre-morbid cerebral defects, behavioral disorders, psychiatric, somatic, and social abnormalities, enuresis nocturna without severe psychiatric comorbidity before the age of 14 years) and lowest in Type 1 patients (suffer from a severe alcohol withdrawal syndrome, and tend to use alcohol to prevent or weaken withdrawal symptoms). Furthermore, leptin serum levels were positively associated with craving particularly in patients of Lesch´s Type 1 and 2 (alcohol used as a means of self-medication due to its sedative effect), and not in patients of Type 3 (alcohol is used as an anti-depressant) and Type 4 (Hillemacher et al. 2007b). In summary, the available studies suggest a pathophysiologic role of leptin in alcohol craving, possibly mediated via its impact on the HPA axis (Hillemacher et al. 2007a), at least in some alcoholic subtypes.
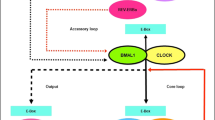
Ghrelin and Growth Hormone. Ghrelin is a 28-amino acid gut peptide acting as the endogenous ligand for the growth hormone secretagogue (GHS) receptor (Kojima et al. 1999). A growing literature has demonstrated a key role of ghrelin in controlling appetite and food intake (reviewed in: Suzuki et al. 2011, see Fig. 1.). Subsequently, research has moved forward to investigating the potential role of ghrelin in alcoholism. Preclinical studies demonstrate that ghrelin centrally administered into the ventral tegmental area (VTA) increases extracellular concentrations of accumbal dopamine (Jerlhag et al. 2007), and both ghrelin and ethanol activate the cholinergic-dopaminergic reward link (Jerlhag et al. 2007, 2008). Similar results were found when ghrelin was injected peripherally (Jerlhag 2008). Moreover, ghrelin administration into the VTA increases alcohol consumption under a 2-bottle alcohol/water free choice paradigm, while alcohol consumption is significantly reduced using genetic and pharmacological models of suppressed ghrelin signaling, i.e. knockout mice for the ghrelin receptor and use of antagonists of the ghrelin receptor, respectively (Jerlhag et al. 2009).

Main Positive and Negative hormonal feed-backs involved in the control of Growth Hormone (GH) release. (+: releasing action; -: inhibiting action, GHIH: GH-inhibiting hormone, (a.k.a. somatostatin), GHRH: GH-releasing hormone, IGF-1: Insulin Growth Factor-1)
While studies with healthy controls consistently report that alcohol consumption acutely reduces plasma ghrelin levels (Calissendorff et al. 2005, 2006; Zimmermann et al. 2007), when studied in AD patients, the findings on ghrelin levels have been conflicting. Differences in subtypes of alcoholic individuals have been proposed as a possible reason to explain these results (Hillemacher et al. 2007b). Furthermore, it seems reasonable that this was due to plasma ghrelin levels investigated in patients at different stages of their alcohol drinking, i.e. active drinkers vs. abstinent. Moreover, plasma ghrelin levels were lower in those studies with actively drinking AD patients (Addolorato et al. 2006; Badaoui et al. 2008), while ghrelin levels were higher in those with abstinent patients (Kim et al. 2005; Kraus et al. 2005). Other factors that could explain the discrepancy between these studies include differences related to patients’ gender and genotype(s) (Leggio 2009).
In order to understand better the link between plasma ghrelin levels and alcohol drinking in AD individuals, a recent longitudinal study assessed repeated plasma ghrelin determinations in a sample of AD patients. There was a statistical difference for ghrelin between non-abstinent and abstinent subjects, with a statistically significant difference in baseline ghrelin levels, i.e. ghrelin levels were higher in those individuals who drank during the 12-week period of the study, while baseline ghrelin levels were lower in those who did not drink (Leggio et al. 2011a). Furthermore, baseline ghrelin levels were significantly and positively correlated with most of the alcohol craving scores assessed during the study (Leggio et al. 2011a). Consistent with Leggio et al. (2011a), a more recent study reported a significant positive correlation between the plasma concentration of active ghrelin and alcohol craving (Koopmann et al. 2011), further supporting the overall hypothesis that ghrelin is potentially able to affect alcohol-seeking behaviors, such as alcohol drinking and craving, representing a new potential neuropharmacological target for AD (Leggio 2010, 2011b).
The physiological role for which ghrelin was discovered is the release of growth hormone (GH; Kojima et al. 1999). Growth hormone release is under the control of several hormones, as well as under the control of neurotransmitter, in particular gamma-aminobutyric acid (GABA)—which is a key neurotransmitter involved in the neurobiology of alcoholism—given the high expression of GABA receptors in the pituitary GH cells (Gamel-Didelon et al. 2002). Human studies comparing normal controls vs. AD individuals demonstrated that the administration of the GABAB receptor agonist baclofen 10 mg (Vescovi et al. 1998; Vescovi and Coiro 2001) or 20 mg (Ozsoy et al. 2007), compared to placebo, induced a significant increase in serum GH levels in healthy controls, while GH levels did not change in AD individuals. This data suggests that unlike healthy controls, AD individuals lose normal physiological GH release after a challenge with baclofen. Furthermore, a recent pilot human lab study found a trend toward a significant reduction in GH levels in AD individuals who received baclofen 30 mg a day for 1 week, and that GH levels correlated positively with the craving score (Leggio et al. 2011c). Though a small sample, baclofen reduced GH levels that were associated with lower craving scores. This leads us to hypothesize a potential dose and time-dependent role of GH in baclofen’s central mechanism of action for alcohol craving.
Insulin and Insulin Growth Factor-1 (IGF-1). Insulin, together with the C-peptide, is secreted by the pancreatic (β)-cells and involved in the regulation of glucose homeostasis. Insulin also plays an important central role in appetitive behaviors (Pristupa et al. 1994; Figlewicz 2003) and has also been suggested to have a role in alcoholism. For example, in the fruit fly Drosophila melanogaster, a role of the Insulin/Insulin-Receptor pathway in regulating behavioral responses to alcohol has been demonstrated (Corl et al. 2005). In humans, a set of three studies investigated the relationship between blood insulin and alcohol craving in AD individuals. The results showed a significant positive correlation between blood insulin level and alcohol craving (Leggio et al. 2008a). Moreover, the insulin level was strongly correlated with the compulsive subscale of the alcohol OCDS craving score, while a correlation trend was found with the obsessive subscore. Interestingly, this positive correlation between insulin and craving was present in actively drinking AD patients, but disappeared in the same patients after 12-weeks of abstinence, suggesting that the link between insulin and craving was specific to drinking (Leggio et al. 2008a). The link between insulin and the dopaminergic system (in particular with the dopamine transporter; Pristupa et al. 1994) could represent a possible neurobiological explanation of these findings. Additionally, the correlation between alcohol craving and insulin was somewhat unique for insulin. In fact, in these experiments, blood C-peptide and IGF-1 levels were also tested. C-peptide, cleaved-off during the synthesis of insulin, showed significant correlations with alcohol craving, similar to insulin (Leggio et al. 2008a). However, no correlation between alcohol craving and IGF-1, an insulin-like hormone produced by the liver, was found during either the active drinking phase or during abstinence. Interestingly, a link between glucose homeostasis, which is under the direct control of insulin levels, and alcohol-seeking behavior has been suggested. For example, C57BL mice which are hyperglycemic (Connelly et al. 1983) as well as Wistar rats with glucose intolerance (Zito et al. 1984) have shown a preference for ethanol. In humans, Blum et al. (2007) developed a ‘reward deficiency syndrome’ model, suggesting that subjects with an addiction disorder (i.e., alcohol dependence, binge eating) have genetic alterations of the dopamine brain system and that the dopamine-glucose link plays a key role in this syndrome. More recently, an analysis of the Combined Pharmacotherapies and Behavioral Interventions for Alcohol Dependence sample (Anton et al. 2006), suggested that in AD individuals, higher baseline blood glucose level predicted greater subsequent alcohol use, and these observations were found even after controlling for baseline drinking, gender and body mass index levels (Leggio et al. 2009b). Together, these data (Leggio et al. 2008a, 2009b) are consistent with the knowledge that chronic alcohol use may reduce insulin sensitivity, leading to elevated insulin and glucose levels. This suggests that manipulations of the insulin signaling might represent a novel pharmacological approach to treat alcoholism.
Adiponectin. Adiponectin, a peptide secreted by white adipocytes, is also involved in hypothalamic appetite regulation (Ahima 2004; Qi et al. 2004). A recent study of blood adiponectin level in AD patients found a significant decrease of adiponectin during the course of withdrawal (Hillemacher et al. 2009a). Furthermore, adiponectin level showed a significant negative correlation with self-rated craving, but only in men. In summary, adiponectin could be involved in the neurobiology of alcohol craving, possibly via its effects on the hypothalamic circuits (Hillemacher et al. 2009a). However, further studies are needed in order to clarify the role of adiponectin in alcoholism, as well as potential gender differences.
Thyroid Hormones. Thyroid hormones help regulate appetite; patients with thyroid diseases often report significant changes in appetite and weight, with higher levels of thyroid hormones inducing a higher appetite (Pijl et al. 2001). The hypothalamic-pituitary-thyroid (HPT) axis also may play an important role in AD patients, including its ability to affect anxious and/or depressed mood (Hermann et al. 2002). Interestingly, the HPT axis may affect key neurotransmitters involved in alcohol-seeking behaviors, e.g. thyroid-stimulating hormone (TSH) is modulated by dopamine (Cooper et al. 1983) in the hypothalamus and/or pituitary gland (Delitala et al. 1981). Furthermore, some studies have reported a reduction in peripheral thyroid hormones and/or blunted TSH response to thyrotropin releasing hormone (TRH) in AD individuals (Välimäki et al. 1984; Baumgartner et al. 1994; Garbutt et al. 1996). A more recent study specifically analyzed the potential relationship between thyroid hormones and craving in AD subjects. This study reported a significant negative correlation between TSH and alcohol craving, and a significant positive correlation between the free-T3 and some of the alcohol craving scores (Leggio et al. 2008b). Furthermore, TSH showed a negative significant correlation with assessments measuring state anxiety, trait anxiety, current depression and aggressiveness. All of these significant correlations, but the one between free-T3 and alcohol craving, disappeared after 12 weeks of abstinence (Leggio et al. 2008b). Together, these findings suggest a possible explanation of the role of the HPT-axis in alcohol craving, especially during the active drinking phase. Interestingly, TSH is modulated by dopamine (Delitala et al. 1981; Cooper et al. 1983) and alcohol craving is inversely correlated with dopamine D2 receptors availability (Heinz et al. 2004), therefore the link between TSH and dopamine might explain the findings on the relationship between thyroid hormones and alcohol craving. Furthermore, this relationship might also be mediated by the potential link between TSH and depressive or anxious mood, which in turn may influence alcohol craving levels.
In summary, there is an increasing interest in the role of appetitive pathways and their response in alcohol craving and dependence. These circuits involve peptides that are either orexigenic (e.g. ghrelin), anorexigenic (e.g. leptin, insulin, adiponectin) or both (e.g. thyroid hormones). Research demonstrates that alcohol and food-seeking behaviors share some common neural pathways as overlapping neuronal circuits exist between addiction and obesity. Neurobiological circuits related to food-seeking behavior have been suggested as possible new targets in the treatment of alcohol dependence. Consistent with these observations, there is converging research suggesting that feeding-regulating circuits may provide important information in understanding alcohol craving and dependence.
Reproductive Hormones
Reproductive, or “sex” hormones, appear to promote permanent and transient effects on the brain. A combination of exposure to the wrong reproductive hormone at the wrong time or in the wrong concentration, in utero or during development, may contribute to developing an increased risk of AD. Moreover, reproductive hormone levels promote alcohol consumption which can also bidirectionally alter reproductive hormone levels. As a result, the change levels of these hormones vary throughout the addiction process (Lenz et al. 2012).
Prolactin. Prolactin, a peptide hormone produced by lactotropes of the anterior pituitary (Wurst et al. 2007), is essential for the initiation and maintenance of lactation (Heil and Subramanian 1998). Lactotrope function is primarily regulated by thyrotropin-releasing hormone (TRH) which facilitates, and dopamine which inhibits, the synthesis and secretion of prolactin. There is also preliminary evidence that ghrelin stimulates prolactin secretion in healthy women, but the effect is less than TRH and is not additive to TRH (Messini et al. 2010a). Prolactin also decreases lutienizing (LH) and follicle stimulating hormone (FSH) secretion that accounts for the lack of fertility and changes in the menstrual cycle associated with hyperprolactinemia (Murphy et al. 2006). Alcohol consumption has long been known to stimulate dopamine receptors (Imperato and Di Chiara 1986), and given that prolactin release is governed by dopamine and D2 receptors, an association with the mesolimbic dopamine system also regulated by D2 receptors, would be expected (Wilhelm et al. 2011). However, while the extent to which the mesolimbic DA pathway is involved in control of prolactin release in humans is unknown, potentially this pathway may be mediated by the HPA axis and D2 receptors involved with both the mesolimbic and tuberoinfundibular systems (Hillemacher et al. 2005).
The most prominent function of prolactin in concert with oxytocin, progesterone, estradiol, GH and glucocorticoids, is to initiate and support lactation (Murphy et al. 2006). Alcohol’s effects on prolactin release have not been studied in lactating women, however Volpi et al. (1994) reported that a blood alcohol level of about 0.08 % diminished prolactin secretion in response to breast stimulation in a study of non-lactating non-alcoholic women controls.
Differences in prolactin response to alcohol appear to differ based on level of active drinking, withdrawal (with and without seizures), as well as having a positive family history of alcoholism, Lesch typology and gender. Mendelson and Mello (1988) reported that elevated plasma prolactin levels were found in premenopausal women heavy drinkers. In 26 healthy, adult women heavy, social and occasional drinkers, no evidence of menstrual cycle dysfunction or abnormality in reproductive hormone levels was found in the occasional drinkers or in two of the social drinkers who consumed less than an average of 3 drinks a day. By contrast, half of the social drinkers who consumed more than 3 drinks a day and 60 % of the heavy drinkers had significant alterations of their menstrual cycle and reproductive hormone function. The major abnormality found in social drinkers was anovulatory cycles, and 3 of the 5 women who were heavy drinkers had persistent hyperprolactinemia. These findings suggest that alcohol-related menstrual cycle and reproductive hormone dysfunction may be more prevalent among women who are social and heavy drinkers (Mendelson and Mello 1988). Likewise, Frias et al. (2002) demonstrated in 12 men and 9 women that acute alcohol intoxication produces a high increase in plasma prolactin, corticotropin (adrenocorticotropic hormone: ACTH), and cortisol in adults of both sexes. Additionally they noted a decrease in LH levels only in men, and reported changes to testosterone according to gender, with increased plasma testosterone in women and a decrease in men. The authors also reported that ACTH and prolactin correlated positively with cortisol, and testosterone in women, which suggests that prolactin and ACTH could contribute to stimulated adrenal androgen production.
On the other hand, differences in response to alcohol have also been demonstrated to be related to a positive family history of alcoholism. Controlling for baseline prolactin measures and hormonal changes after placebo, Schuckit and colleagues (1987) reported that 30 sons of alcoholic individuals compared to matched controls had significantly lower prolactin levels in response to a high-dose alcohol challenge.
During the first 14 days of withdrawal and early abstinence, Wilhelm and colleagues (2011) assessed prolactin levels in 99 AD men compared to 43 controls and reported that prolactin levels were elevated during the entire period in the AD subjects but not the healthy controls. Prolactin levels on day 1 of the study were significantly associated with the severity of alcohol withdrawal and AD but not craving, depressive symptoms, or anxiety (Wilhelm et al. 2011). The lack of association with craving, depression symptoms and anxiety, contrasts with previous data suggesting a significant positive relationship between depressed mood and prolactin level during withdrawal (Miller et al. 1986). Alternatively, this primary outcome is consistent with the finding that prolactin may be a clinical marker for severity of alcohol withdrawal in men, as increased prolactin levels were associated with an elevated risk for alcohol withdrawal seizures in men admitted to alcohol detoxification (Hillemacher et al. 2007b). The authors hypothesize that the significant association of prolactin levels at admission and the severity of AD, support the notion that alterations in prolactin serum levels result from long-term alcohol induced disturbances in dopamine circuits (Wilhelm et al. 2011).
Researching the dopamine-prolactin connection between the mesolimbic and tuberoinfundibular systems, Hillemacher and colleagues assessed alcohol craving reported by 145 men and women during early alcohol withdrawal. Increased levels of prolactin were demonstrated to be significantly related to lower craving in women alone, and prolactin levels declined significantly in women during withdrawal as well (Hillemacher et al. 2005). The authors suggest that the increase in prolactin levels in women AD patients associated with lower craving may denote normalization of the chronically stimulated DA pathway during active drinking (Gessa et al. 1985) and the HPA axis during alcohol withdrawal (Hillemacher et al. 2005). Certainly, research performed on healthy women where the dopamine agonist drug bromocriptine was shown to block intravenous ghrelin stimulation of prolactin secretion in women should, be considered in AD patients to tease apart the importance of prolactin to craving in men and women (Messini et al. 2010b).
Luteinizing Hormone (LH), Follicle Stimulating Hormone (FSH) and Testosterone. For roughly 40 years, evidence suggests that alcohol affects sex hormones (Galvão-Teles et al. 1973). Sexual disturbances are a common medical consequence of alcohol abuse and are associated with dysregulation of sexual hormones including testosterone (Castilla-García et al. 1987). As shown in Fig. 2, the hypothalamus produces gonadotropin-releasing hormone (GnRH), also known as Luteinizing-hormone-releasing hormone (LHRH) and luliberin. LHRH is a tropic peptide hormone released into blood vessels that connect the hypothalamus and the anterior pituitary. In response to the release of LHRH, the anterior pituitary produces and releases two protein hormones, follicle-stimulating hormone (FSH) and luteinizing hormone (LH) that are then released into the body's general circulation and act primarily on the gonads. In men, LH stimulates testosterone production from Leydig cells in the testes, while in the epididymis, FSH is important for sperm maturation. Testosterone feeds back to the hypothalamic-pituitary circuit and regulates the further production and secretion of LHRH and LH (Goodman 2003).

Reproductive function in men and women. To the left, negative feedback of testicular function; (+) stimulation and (-) inhibition. To the right ovarian pituitary interactions during the 3 phases of the menstrual cycle. Solid arrows indicate stimulation while dashes indicate inhibition. [permission to reprint granted by Elsevier, Basic Medical Endocrinology, 2nd Edition H. Maurice Goodman 2003]
Androgen is the general term for any natural or synthetic compound that controls the development and maintenance of characteristics in male vertebrates by binding to androgen receptors. This includes the activity of the sex organs and development of secondary sex characteristics in men. In women, androgens are also the original anabolic steroids and the precursor of all estrogens. In addition, the principal sex hormone in men is testosterone (Goodman 2003). Testosterone plays a central role in the development of the testis and prostate as well as promoting secondary sexual characteristics such as increased muscle, bone mass, and the growth of body hair. Moreover, testosterone is essential for health and well-being as well as the prevention of osteoporosis (Goodman 2003).
Under normal functioning, a low testosterone level results in a rise in LH and FSH, however though alcohol use is associated with even lower testosterone, LH levels and to a lesser degree FSH levels, do not rise as expected (Emanuele and Emanuele 2001). Ruusa and colleagues (1997) conducted a study with 29 AD men without severe liver disease during inpatient alcohol detoxification and then 3 weeks later. Hormone levels were assessed the day after admission (day 2), at discharge (day 5) and at the 3-week follow-up. Levels of testosterone and sex hormone-binding globulin (SHBG is a glycoprotein that binds to the sex hormones testosterone and estradiol) were low during the entire withdrawal period but were within normal expected levels; though at the 3-week follow-up these hormone levels rose. On the other hand, while LH and FSH were both high at intake, during the detoxification period, both were depressed. At the 3-week follow-up however, while FSH levels recovered to those at admission, LH levels remained low (Ruusa et al. 1997). One hypothesis is that due to chronic drinking, the feed-back loop becomes dysregulated where low levels of testosterone should activate LH production, to in turn, stimulate testosterone production (Ruusa et al. 1997).
La Grange et al. (1995) on the other hand demonstrated in a non-dependent sample of college student men, increased alcohol use was associated with a higher testosterone level. Moreover, men who are more likely to binge drink, be more highly intoxicated, and more frequently diagnosed with alcohol dependence have higher testosterone levels (Eriksson et al. 2005; Suzuki et al. 2009).
Research on endophenotypes involving hormones represents the nexus of genetic and neuroendocrinology research on how polymorphisms may influence alcoholism and craving (Sturgess et al. 2011). For example, the polymorphic polyglutamine locale on chromosome Xq11-12 in the amino-terminal domain of the androgen receptor, is encoded by the CAG repeat that generally regulates the androgen receptor’s regulation as a transcription factor (Chamberlain et al. 1994). Long repeats of the CAG trinucleotide have been shown to inhibit interactions between androgen receptors and various co-activators (Irvine et al. 2000), and short repeats are associated with a higher risk of depression (Seidman et al. 2001) and other medical conditions (Tut et al. 1997; Westberg et al. 2001). In an effort to examine the influence of this androgen polymorphism on craving Lenz and colleagues (2009) studied 112 men admitted for alcohol detoxification treatment. Craving was assessed by the OCDS on the day of admission and all were genotyped for the androgen CAG polymorphism and were grouped into short (7-18), intermediate (19-23) and long (24-32) repeats. A significant inverse relation was reported between total and obsessive craving and androgen receptor repeat length such that a greater number of length of repeats was significantly associated with attenuated severe craving in men (Lenz et al. 2009). A subsequent study on this group of men in alcohol withdrawal also reported that this polymorphism mediates its effect on craving primarily through regulation of leptin (Lenz et al. 2010).
Gonandal hormone changes may also vary by gender. In a study of intravenous alcohol infusion in 24 men and 24 women, acute alcohol administration resulted in hormonal changes that varied by sex. Predominantly in men, alcohol administration significantly induced suppression of testosterone levels, whereas estradiol was significantly increased in women (Vatsalya et al. 2012). Alcohol has long been suspected to increase the conversion of testosterone into estradiol (Gordon et al. 1979). Accordingly, postmenopausal women who drank (Gavaler and Van Thiel 1992a, b) were found to have higher estradiol levels than abstaining women. Because in postmenopausal women the major source of endogenous estrogen is the aromatization of androgens to estrogens is alcohol, the relationship between moderate alcohol use and serum estradiol levels were assessed in 128 normal postmenopausal women. Among the 21.2 % of women who did not drink alcohol, estradiol levels were significantly lower than in the 78.8 % of women who reported alcohol use. In further analyses, inclusion of alcohol as a variable increased the amount of explained variance in estradiol as did the proxy for aromatization, the estradiol-testosterone ratio logarithm. When examining the results of moderate alcohol use in context with other clinical diagnoses such as with hepatic problems, bone loss, breast cancer and cardiovascular disease (CVD), only the risk of CVD was found to be significantly reduced (Gavaler and Van Thiel 1992a). The conversion of androgens, primarily testosterone, into estradiol is completed by the enzyme aromatase that may be induced by alcohol consumption (Chung 1990), and is a potential pathway to increase the incidence of breast cancer in women who drink alcohol even moderately (Chen et al. 2011).
Estrogen. Alcohol is shown to increase the risk of developing breast cancer (Singletary and Gapstur 2001; Chen et al. 2011; Fernandez 2011) and does so in a linear dose-dependent manner (Smith-Warner et al. 1998). Several hypotheses have been proposed to explain the increased risk of breast cancer such as reactive oxygen species and acetaldehyde (Dumitrescu and Shields 2005) and via insulin like growth factor 1 (IGF-1; Etique et al. 2004). Alcohol also increases serum estrogens (Hankinson et al. 1995; Zhang et al. 2007) and promotes human epidermal growth factor receptor 2 (HER2) breast cancer via the estrogen signaling pathway (Wong et al. 2011). HER2 positive breast cancer is responsible for one-quarter of all breast cancers and has a poor prognosis (Nahta et al. 2009). Wong et al. (2011), demonstrated in mice, that alcohol promoted tumor development only in the presence of physiological levels of estrogen, and that the effect of alcohol on tumor development in HER2 mice is blocked by ovariectomy. One mechanism hypothesizes that alcohol increases the estrogen level is by increasing the level of aromatase, the enzyme that converts androgens to estrogens (Chen et al. 2002; Chung 1990; Etique et al. 2004). High levels of estrogen may then facilitate the development of breast cancer by activating the estrogen pathway (Tan et al. 2009; Wong et al. 2011). Furthermore, after menopause, estrogens are formed from androgens by aromatase (Longcope 1999) and as little as 1 or 2 drinks a day can increase dihydroepiandrosterone and estrone levels and therefore increase the risk of breast cancer in postmenopausal women (Dorgan et al. 2001).
Potentially contributing to the increased risk of breast cancer in women through the pathways just described, is that estrogens are also found in alcoholic beverages themselves. Plants are used to produce alcohol and contain phytoestrogens that are estrogen like substances (Gavaler 1988; Gavaler and Van Thiel 1988; Van Thiel et al. 1991; Rosenblum et al. 1991). Moreover, estrogens affect men as well as women. It is shown that men with alcohol related cirrhosis often suffer from testicular failure and show signs and symptoms of feminization e.g. gynecomastia and redistribution of body fat similar to that seen in women and consistent with elevated estrogen levels (Karagiannis and Harsoulis 2005). These signs and symptoms of feminization, although also present in patients with other forms of cirrhosis (e.g. HCV-related cirrhosis), are particularly important in patients with alcoholic cirrhosis, as two factors are hypothesized to facilitate the feminization of men with alcoholic cirrhosis. The first is chronic exposure to phytoestrogens contained in alcohol and the second is the inability of the alcohol damaged liver to properly metabolize and excrete phytoestrogens (Gavaler et al. 1987; Gavaler 1998).
Hormonal Influences of the Menstrual Cycle. It has long been observed that there is a link between the menstrual cycle and alcohol-seeking behavior by both AD (Belfer et al. 1971; Allen 1996), and non-dependent women (Perry et al. 2004). This link is rather complex as dysfunction caused by alcohol drinking women seems to vary by phase (follicular, ovulation, or luteal phases), whether pre- or post-menopausal, AD or not, drinking status (active, withdrawal, or abstinence) and family history of alcoholism. Moreover, women dependent on alcohol often experience menstrual cycle irregularities such as amenorrhea and infertility (Augustyńska et al. 2007).
A relationship between pre-menstrual syndrome and alcohol consumption was demonstrated in college women who were relatively early in their drinking careers and before the development of severe alcohol-related problems (Perry et al. 2004). Accordingly, it was also demonstrated that family history positive (FHP) women with AD increase their drinking and report less premenstrual symptoms during the luteal phase with respect to the follicular phase than family history negative (FHN) women (Svikis et al. 2006). Evans and Levin (2011) administered various doses of alcohol to 21 FHN and 24 FHP women during the mid-follicular and late luteal phases of the menstrual cycle while administering several cognitive and behavioral tasks. Though less pronounced than previously reported in men, compared to FHN women, FHP women were less impaired by alcohol on a balance task but reported greater dysphoric mood during the luteal phase.
In a study of 30 pre-menopausal women not taking hormones or contraceptives entered into a 30-day alcohol detoxification program, serum levels of hormones including, prolactin, FSH, LH, estradiol, progesterone, and testosterone were assessed during the follicular (between days 5-7), ovulation (days 11-14) and luteal (days 19-22) phases of one full menstrual cycle (Augustyńska et al. 2007). Compared to reference values for healthy women and consistent with data previously presented above, this study reported that mean prolactin levels in all 3 phases were higher and testosterone was lower. Moreover, the authors also reported that the progesterone level was higher but only during the follicular phase and the LH level during the ovulation phase was lower. Significant positive correlations were reported between age and FSH and estradiol during the follicular phase and between testosterone during the luteal phase and a longer duration of AD (Augustyńska et al. 2007).
The acute effect of alcohol on hormones after alcohol intake of 0.5 g/kg (about 2 drinks), was also assessed in 3 substudies of pre-menopausal women: 1) women were either not taking oral contraceptives (n = 30) or taking them (n = 31) across the entire menstrual cycle; 2) women were either not taking oral contraceptives (n = 40) or taking them (n = 47) at the mid-cycle phase; and 3) 10 participants used oral contraceptives without regard to phase (Sarkola et al. 1999). In all the participants who used oral contraceptives, estradiol levels significantly increased and progesterone levels significantly decreased after intake of alcohol. No effect on progesterone was observed in the third substudy on the 10 participants who used oral contraceptives. No effect was reported for changes in estrone levels, though a significant increase was found in the estradiol-to-estrone ratio. Among women using oral contraceptives, progesterone levels decreased significantly after intake of alcohol but no changes in levels were seen in estradiol, estrone, or the estradiol-to-estrone ratio during midcycle in this study group. A transient elevating effect of alcohol on prolactin levels was observed in both study groups and alcohol had no significant effect on LH levels among women not using oral contraceptives (Sarkola et al. 1999). The authors suggest that the changes on prolactin levels may reflect activation of opioid peptides and dopamine in the hypothalamus (Sarkola et al. 1999).
As noted by Lenz and colleagues (2012) the interplay of alcohol consumption and reproductive hormones is complex and several limitations of human association studies between alcohol consumption and sex hormones must be carefully considered. These include gender and the level of maturation in participants, circadian and seasonal rhythms of sex hormones, significant changes of acute and chronic alcohol use, the heterogeneity of AD, and a probable degree of bidirectionality in association studies.
Posterior Pituitary Hormones
Oxytocin. Oxytocin is an endogenous ligand for the oxytocin receptor (OXTR), and differs in two amino acids from the 9-amino acid peptide, arginine vasopressin (antidiuretic hormone, AVP), with which it shares some biological activity (Kostoglou-Athanassiou, et al. 1994; Landgraf and Neumann 2004). Oxytocin is produced in specialized cells in the supraoptic (SO) and paraventricular (PV) nuclei of the hypothalamus, and functions both as a peripheral hormone and as a central nervous system (CNS) signal molecule (Buijs 1983). In its role as a peripheral hormone (see Fig. 3), oxytocin is released into the circulation from the posterior pituitary, enhancing uterine contractions during parturition and with prolactin enhancing milk-letdown during lactation (Ott and Scott 2010). Maternal alcohol use before or during lactation can interfere with the proper function of both prolactin and oxytocin (Heil and Subramanian 1998), a balance which is important, as “Oxytocin serves today’s meal and prolactin prepares tomorrow’s” (Short 1984, p36).

Regulation of posterior pituitary function. To the left, regulation of oxytocin with positive feedback. Oxytocin stimulates the uterus to contract and causes the cervix to stretch. Oxytocin secreted in response to suckling forms and open-loop feedback system in which positive input is interrupted when the infant is satisfied and stops suckling. To the right argentine vasopressin (AVP) negative feedback loops. Increasing blood osmolality or decreased blood volume in the brain or thorax, respectively, increasing vasopressin secretion. Vasopressin acting primarily on the kidney produces changes that restore osmolality and volume thereby attenuating further secretion in a negative feedback loop. [permission to reprint granted by Elsevier, Basic Medical Endocrinology, 2nd Edition H. Maurice Goodman 2003]
In its role as a CNS signaling molecule, oxytocin is released in limbic and forebrain areas from terminals that arise from SO and PV neurons, as well as from extrahypothalamic oxytocin neurons in the amygdala and bed nucleus of the stria terminalis (Ross et al. 2009). CNS release of oxytocin occurs in the amygdala, septum and bed nucleus, in relation to stress (Ebner et al. 2005; Neumann 2007). In the CNS, oxytocin facilitation of social behaviors, and attenuation of social stress and anxiety is complex (Ring 2011), and CNS and peripheral oxytocin release do not always correlate (Neumann 2007; Engelmann et al. 1999; Veening et al. 2010).
Alcohol enhances reward via dopamine-dependent mesolimbic reward pathways (Weiss et al. 1993). Two major dopamine pathways interact with oxytocin: tuberoinfundibular dopamine neurons synapse with oxytocin neurons in the paraventricular nucleus of the hypothalamus and stimulate peripheral oxytocin release (Baskerville et al. 2009); and VTA dopamine neurons and oxytocin neurons from the hypothalamus both project to the amygdala and other limbic areas (Baskerville and Douglas 2010). VTA dopamine acting in the nucleus accumbens enhances social bond formation in both male and female prairie voles (Aragona et al. 2003). In addition, oxytocin neurons in the VPN project to the VTA and also stimulate dopamine release (Melis et al. 2007). In rats, social behaviors that increase oxytocin also activate dopamine release and oxytocin infusion into the VTA of mother rats increased nucleus accumbens dopamine release (Shahrokh et al. 2010), suggesting a direct action of oxytocin on dopamine neurons. While acute alcohol releases dopamine, chronic alcohol use results in hypoactivity of dopamine systems that contributes to allostasis and maintains alcohol consumption (Volkow et al. 2002; Baskerville and Douglas 2010).
In humans, sexual behavior is associated with increases in blood and brain oxytocin activity and increased bonding with and positive feelings about one’s partner (Carmichael et al. 1987; Bancroft 2005). Oxytocin is important for mother-infant bonding in humans (Uvnäs-Moberg 1996) and has been demonstrated to work through the parasympathetic nervous system to enhance response and increase motivational value of facial expressions (Gamer and Büchel 2012). In couples who received either double-blind intranasal oxytocin or placebo before a standardized conflict discussion, oxytocin significantly increased positive communication during the conflict discussion and reduced post-conflict salivary cortisol (Ditzen et al. 2009).
Alcohol dependence is associated with poor interpersonal relationships and poor social coping skills (Monti et al. 1994; Anacker and Ryabinin 2010). Chronic alcoholics show specific deficits in social behaviors that could be mediated at least partially by oxytocin (Domes et al. 2007a; b). Alcohol’s effects on oxytocin depend on alcohol dose and the release rate of oxytocin, but oxytocin release is completely inhibited at alcohol doses of 1 g/kg (Fuchs and Wagner 1963; Cobo 1973). Alcohol blocks the breast stimulation-induced increase in plasma oxytocin levels in lactating (Mennella et al. 2005) and non-lactating women (Coiro 1992).
The effect of oxytocin to enhance attachment and social behaviors (Kosfeld et al. 2005; Zak et al. 2007; Ditzen et al. 2009; Hurlemann et al. 2010) suggests that oxytocin could be an effective adjunct for psychotherapy as positive alliances with therapists are related to better psychotherapy outcomes. The social facilitation effects of oxytocin, including increasing trust, empathy and attachment, and reducing conflict behaviors (Kosfeld et al. 2005; Zak et al. 2007; Ditzen et al. 2009; Hurlemann et al. 2010) may be expected to have a positive effect on client engagement and disclosure. A placebo-controlled trial of intranasal oxytocin as an adjunct to exposure therapy in social anxiety disorders did not improve anxiety; however, those receiving oxytocin did have a more positive self-image (Guastella et al. 2009).
Several reviews propose that the oxytocin system is a major contributor to drug and AD (Sarnyai and Kovács 1994; Kovács et al. 1998; McGregor et al. 2008). Oxytocin and its analogue AVP are involved in the acute and chronic actions of alcohol, including the development of tolerance (Tabakoff and Hoffman 1988). The tolerance appears to be due to an alcohol induced decrease in calcium-activated potassium channels on the cells (Knott et al. 2002). Oxytocin and AVP show opposite effects of alcohol tolerance and withdrawal. In mice, tolerance to ethanol hypothermia was blocked if the mice were treated with a range of doses of oxytocin along with the ethanol (Szabó et al. 1985). In contrast, low doses of AVP facilitated the development of hypothermic tolerance. Oxytocin also inhibits the development of tolerance to opiates (Kovács and Telegdy 1987). Decreased alcohol tolerance and increased sensitivity to alcohol effects have been proposed as endophenotypes that reduce risk of developing AD (Schuckit 1994).
Few studies have administered oxytocin to AD humans to determine its effect on drinking. Marchesi et al. (1997) followed 13 newly abstinent AD men over 28 days and found plasma oxytocin levels to be elevated, with the highest levels on day 1 after withdrawal. Oxytocin levels dropped over the 4 weeks but were still slightly elevated on day 28.
Anxiety is a risk factor for AD both at an individual (Schuckit 1996; Willinger et al. 2002) and at a population genetic level (Merikangas et al. 1998; Enoch et al. 2008). Alcohol has anxiolytic properties, for example reducing visual and limbic system MRI bold responses to fearful faces (Gilman et al. 2008); however, rapid tolerance also develops to alcohol’s anxiolytic effect (Sakharkar et al. 2011). Oxytocin shows anxiolytic effects in both animals and humans. In humans, the majority of studies find an inverse relationship between anxiety and oxytocin levels (Scantamburlo et al. 2007). In a small double-blind crossover study in 15 men, intranasal oxytocin compared to placebo, reduced fear response and amygdala activation by fear-inducing visual stimuli (Kirsch et al. 2005). Conflicting results are observed in patients with social anxiety disorder, in which oxytocin levels positively correlate with levels of anxiety (Hoge et al. 2008) and a study of social anxiety disorder patients found no benefit of a single weekly dose of oxytocin administered prior to exposure therapy (Guastella et al. 2009).
The anxiolytic effects of oxytocin have possible implications for AD treatment. Alterations in several neurotransmitters in the amygdala contribute to both increased anxiety and increased alcohol intake, including neuropeptide Y (Pandey, et al. 2005), GABA (Hyytia and Koob 1995; Kumar et al. 2009), and CRF (Enoch et al. 2008). Interventions that reduce anxiety may be effective alcoholism treatments. For example, in a clinical trial with anxious alcoholics, buspirone, an anxiolytic 5-HT1 agonist that also induces oxytocin release (Uvnäs-Moberg 1996), was found to reduce anxiety and improve study retention and drinking outcomes better than placebo (Kranzler et al. 1994).
Arginine Vasopressin (AVP) and atrial natriuretic peptide (ANP). In early abstinent AD patients, studies describe changes in volume regulating hormones (Kiefer et al. 2002; Hillemacher et al. 2006), that may be linked with alcohol craving and dependence (Hillemacher et al. 2009b). Among these volume-regulating hormones, AVP and ANP that is released to reduce blood pressure by atrial myocytes in response to high blood pressure, have received the greatest attention (Döring et al. 2003), as detailed next.
In the periphery, AVP is the endogenous ligand for the AVP receptor AVPR2 and decreases urine volume and increases osmolality by enhancing water reabsorption in renal tubules. In the CNS specifically, AVP acts at the vasopressin/V1a (AVPR1a) and vasopressin/V1b (AVPR1b) receptors to affect social behaviors and cognition, often opposite to oxytocin (Israel et al. 2008). For example, intranasal AVP enhances and oxytocin impairs word recall in humans (Heinrichs et al. 2004) and may possess anxiolytic properties (Landgraf 2005), while in the amygdala, AVP and oxytocin also have opposite effects on neuronal excitation (Raggenbass 2008).
Clearly, prolonged alcohol intake leads to degeneration of neurons of the hypothalamic magnocellular nucleus of the PVN (Silva et al. 2002). Further, that AVP plasma levels in AD patients are decreased over months of alcohol abstinence (Döring et al. 2003). In particular, plasma AVP was continuously suppressed in older healthy men as well as recently abstinent alcoholics anticipating novel stimuli, compared to healthy young men where AVP levels were significant increased (Ehrenreich et al. 1997). This suppression in peripheral levels of AVP however, does not appear to affect the central AVP response to increased plasma osmolality in long-term abstinent alcoholics (Jahn et al. 2004).
The exact role of AVP as a stress hormone is still being studied, though several lines of evidence suggest that AVP is involved in the pathophysiology of stress-related behaviors (Surget and Belzung 2008; Chrousos 2009). For example, AVP is known to potentiate the effects of CRF on the HPA axis as the gradual development of excessive alcohol use is associated with a negative emotional state typified by symptoms of depression and anxiety (Koob and Le Moal 1997). Though the data is from rodents, emerging evidence also suggests a role for centrally located AVP acting on the AVPR1b in the regulation of stress- and anxiety-like behaviors in rodents. Edwards et al. (2012) reported that AVPR1b antagonist SSR149415 dose-dependently reduced ethanol self-administration in ethanol-dependent rats and that this effect was specific for the ethanol-dependent but not for non-dependent rodents. An additional interesting observation by the same investigators was that ethanol self-administration reduced AVPR1b levels in the basolateral amygdala of non-dependent animals while AVPR1b levels were seemingly restored in ethanol-dependent rats, suggesting that the AVPR1b may be involved in the processes responsible for the transition to AD in humans (Edwards et al. 2012).
Modulating the effects of AVP, ANP has been demonstrated to have a powerful anxiolytic action (Ströhle and Holsboer 2003; Ströhle et al. 2006), and has been shown to inhibit the secretion of CRH, ACTH and cortisol (Antoni et al. 1992). Dysregulation of ANP plasma levels has been found in depression, eating disorders and alcohol withdrawal (Ohashi et al. 1988; Kiefer et al. 2002; Ströhle and Holsboer 2003). Epigenetic changes of DNA methylation and of promoter-specific DNA methylation have been described in AD. For example, significantly greater homocysteine-induced endoplasmic reticulum protein (HERP) promoter DNA methylation and lower HERP mRNA expression have been reported, in AD patients as compared to healthy controls (Bleich et al. 2006). DNA methylation changes have also been investigated in studies assessing the role of volume-regulating hormones in AD. For example, Hillemacher and colleagues (2009b) investigated alterations of the promoter-related DNA methylation of the ANP and AVP precursor genes and related mRNA-expression of these genes in early alcohol withdrawal in blood samples from 57 healthy controls and 111 patients with AD admitted for detoxification. ANP mRNA-expression was significantly elevated in alcoholic patients while promoter- related DNA methylation of ANP was significantly decreased. Additionally in these AD patients, craving (assessed by OCDS) was significantly associated with promoter-related DNA methylation of ANP suggesting significant alterations of the mRNA-expression and promoter-related DNA methylation of AVP and particularly ANP precursor genes.
Conclusion
Recent neuroendocrine research suggests that there is a significant modulating role for many hormones in the etiology of alcoholism. Moreover, appetitive, reproductive and posterior pituitary hormones discussed in this review, have significant importance to the development and maintenance of alcoholism in humans. Changes in hormone levels are associated with alcohol craving, drinking, withdrawal, abstinence and relapse, as neuroendocrine mechanisms are highly associated with the neurobiology of alcoholism. Whether hormonal changes are caused by, or are the result of alcoholism, are only beginning to be understood as most data only demonstrate associations between neuroendocrine systems and dysfunctional drinking, not causality.
As has been noted throughout this review there are significant methodological differences among many of these studies that must be considered in evaluating these data. Hormone levels vary based on a multitude of factors such as age, nutritional status, differences of when and how many blood samples are drawn, assay variability, trials with and without pharmacological treatment, the various lengths of trials, and subtypes of alcoholics. Moreover, it is also evident that hormone levels fluctuate during stages of alcohol drinking, withdrawal and abstinence. Additionally, alcohol-induced changes in hormones vary across individuals and also depend on gender, specific phases of the menstrual cycle in women, family history of alcoholism, and predisposing genotypes. How all of these changes in hormonal levels perpetuate drinking or the interaction with genetics contribute to relapse in humans, are still largely unknown.
Substantial research suggests that alcoholics are a heterogeneous population and that several genetic polymorphisms involving hormones may predispose some to increased alcohol craving and dysfunctional alcohol use, whereas other gene variations may actually confer protection. Furthermore, numerous endocrine mechanisms could represent future pharmacological targets. Whether pharmacotherapy could be potent enough to make an impact on underlying neuroendocrine function, to lead to, and promote abstinence, is only speculation. Certainly, inducing craving by administering a hormone e.g. ghrelin or prolactin, could lead to lab models to test the efficacy of pharmacologically active antagonists, or combining oxytocin with talk therapy could putatively lead to therapeutic breakthroughs, saving considerable time, money and treatment failures. It is possible that in the future, subtypes of alcoholic populations may gain the most benefit from developed treatments that are tailored and targeted for smaller homogeneous groups. Knowledge about how hormones fluctuate, particularly in alcoholic subtypes e.g. age, gender, phase of drinking, withdrawal, abstinence or relapse, may 1 day help lead to treatments that reduce craving and ultimately alcohol consumption.
In conclusion, due to the complex mechanism of action of alcohol, it is not unrealistic that several treatments, primarily pharmacological, will be required to be combined in order to provide a requisite statistically and clinically robust reduction in alcohol consumption, or as in some cases such as alcoholic liver disease or hepatitis, complete abstinence. While abstinence is a worthy goal for all treatments, perhaps achieving perfection is clinically unrealistic. Furthermore, given that so many individual drugs have not demonstrated a consistent reduction in alcohol use in clinical trials, gives rise to the notion of a more aggressive polyneuronal pharmacological approach that can only be pursued by combining drugs that, as long as there is no evidence of harmful proximal or distal adverse effects, can provide drug action that cannot be duplicated with a single drug. Such poly-pharmacy is the state-of-the-art approach for many disease-states such as diabetes, oncology, hypertension, infectious and cardiovascular diseases, as well as many psychiatric diseases. Why alcohol researchers should be so surprised that our current treatments continue to fall short of our own, and our patients’ expectations, should be no surprise at all.
References
Addolorato, G., Capristo, E., Leggio, L., Ferrulli, A., Abenavoli, L., Malandrino, N., et al. (2006). Relationship between ghrelin levels, alcohol craving, and nutritional status in current alcoholic patients. Alcoholism: Clinical and Experimental Research., 30, 1933–1937.
Adinoff, B., Junghanns, K., Kiefer, F., & Krishnan-Sarin, S. (2005). Suppression of the HPA axis stress-response: implications for relapse. Alcoholism: Clinical and Experimental Research, 7, 1351–1355.
Ahima, R. S. (2004). Adiponectin acts in the brain to decrease body weight. Nature Medicine., 10, 524–529.
Allen, D. (1996). Are alcoholic women more likely to drink premenstrually? Alcohol Alcoholism., 31, 145–147.
Anacker, M. J., & Ryabinin, A. E. (2010). Biological contribution to social influences on alcohol drinking: evidence from animal models. International Journal Environmental Research Public Health, 7, 473–493.
Anton, R. F., O'Malley, S. S., Ciraulo, D. A., Cisler, R. A., Couper, D., Donovan, D. M., et al. (2006). Combined pharmacotherapies and behavioral interventions for alcohol dependence: the COMBINE study: a randomized controlled trial. Journal of the American Medical Association., 295, 2003–2017.
Antoni, F. A., Hunter, E. F., Lowry, P. J., Noble, J. M., & Seckl, J. R. (1992). Atriopeptin: an endogenous corticotropin-release inhibiting hormone. Endocrinology, 130, 1753–5.
Aragona, B. J., Liu, Y., Curtis, J. T., Stephan, F. K., & Wang, Z. (2003). A critical role for nucleus accumbens dopamine in partner-preference formation in male prairie voles. Journal of Neuroscience., 23, 3483–3490.
Augustyńska, B., Ziółkowski, M., Odrowaz-Sypniewska, G., Kiełpiński, A., Gruszka, M., & Kosmowski, W. (2007). Menstrual cycle in women addicted to alcohol during the first week following drinking cessation–changes of sex hormones levels in relation to selected clinical features. Alcohol Alcoholism, 42, 80–3.
Badaoui, A., De Saeger, C., Duchemin, J., Gihousse, D., de Timary, P., & Stärkel, P. (2008). Alcohol dependence is associated with reduced plasma and fundic ghrelin levels. European Journal of Clinical Investigation, 38, 397–403.
Bancroft, J. (2005). The endocrinology of sexual arousal. Journal of Endocrinology., 186, 411–27.
Baskerville, T. A., & Douglas, A. J. (2010). Dopamine and oxytocin interactions underlying behaviors: potential contributions to behavioral disorders. CNS Neuroscience & Therapeutics, 16, e92–e123.
Baskerville, T. A., Allard, J., Wayman, C., & Douglas, A. J. (2009). Dopamine–oxytocin interactions in penile erection. European Journal of Neuroscience, 30, 2151.
Baumgartner, A., Rommelspacher, H., Otto, M., Schmidt, L. G., Kürten, I., Gräf, K. J., et al. (1994). Hypothalamic pituitary thyroid (HPT) axis in chronic alcoholism. I. HPT axis in chronic alcoholics during withdrawal and after 3 weeks of abstinence Alcoholism. Clinical and Experimental Research, 18, 284–294.
Belfer, M. L., Shader, R. I., Corroll, M., & Harmatz, J. S. (1971). Alcoholism in women. Archives of General Psychiatry, 25, 540–544.
Bellem, A., Meiyappan, S., Romans, S., & Einstein, G. (2011). Measuring estrogens and progestagens in humans: an overview of methods. General Medicine, 8, 283–99.
Bleich, S., Lenz, B., Ziegenbein, M., Beutler, S., Frieling, H., Kornhuber, J., et al. (2006). Epigenetic DNA hypermethylation of the HERP gene promoter induces down-regulation of its mRNA expression in patients with alcohol dependence. Alcoholism: Clinical and Experimental Research, 30, 587–91.
Blum, K., Chen, T. J., Meshkin, B., Downs, B. W., Gordon, C. A., Blum, S., et al. (2007). Genotrim, a DNA-customized nutrigenomic product, targets genetic factors of obesity: hypothesizing a dopamine-glucose correlation demonstrating reward deficiency syndrome (RDS). Medicine Hypotheses, 68, 844–52.
Buijs, R. M. (1983). Vasopressin and oxytocin—their role in neurotransmission. Pharmacology and Therapeutics, 22, 127–141.
Bulik, C. M., Klump, K. L., Thornton, L., Kaplan, A. S., Devlin, B., Fichter, M. M., et al. (2004). Alcohol use disorder comorbidity in eating disorders: a multicenter study. Journal of Clinical Psychiatry, 65, 1000–6.
Calissendorff, J., Danielsson, O., Brismar, K., & Röjdmark, S. (2005). Inhibitory effect of alcohol on ghrelin secretion in normal man. European Journal of Endocrinology, 152, 743–747.
Calissendorff, J., Danielsson, O., Brismar, K., & Röjdmark, S. (2006). Alcohol ingestion does not affect serum levels of peptide YY but decreases both total and octanoylated ghrelin levels in healthy subjects. Metabolism, 55, 1625–1629.
Carmichael, M. S., Humbert, R., Dixen, J., Palmisano, G., Greenleaf, W., & Davidson, J. M. (1987). Plasma oxytocin increases in the human sexual response. Journal of Clinical Endocrinological Metabolism, 64, 27–31.
Castilla-García, A., Santolaria-Fernández, F. J., González-Reimers, C. E., Batista-López, N., González-García, C., Jorge-Hernández, J. A., et al. (1987). Alcohol-induced hypogonadism: reversal after ethanol withdrawal. Drug and Alcohol Dependence, 20, 255–60.
Chamberlain, N. L., Driver, E. D., & Miesfeld, R. L. (1994). The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Research, 22, 3181–6.
Chen, S., Zhou, D., Okubo, T., Kao, Y. C., Eng, E. T., Grube, B., et al. (2002). Prevention and treatment of breast cancer by suppressing aromatase activity and expression. Annals New York Academy of Science, 963, 229–38.
Chen, W. Y., Rosner, B., Hankinson, S. E., Colditz, G. A., & Willett, W. C. (2011). Moderate alcohol consumption during adult life, drinking patterns, and breast cancer risk. Journal of the American Medical Association, 306, 1884–90.
Chrousos, G. P. (2009). Stress and disorders of the stress system. Nature Review Endocrinology, 5, 374–81.
Chung, K. W. (1990). Effects of chronic ethanol intake on aromatization of androgens and concentration of estrogen and androgen receptors in rat liver. Toxicology, 62, 285–295.
Ciccocioppo, R., Gehlert, D. R., Ryabinin, A., Kaur, S., Cippitelli, A., Thorsell, A., et al. (2009). Stress-related neuropeptides and alcoholism: CRH, NPY, and beyond. Alcohol, 43, 491–8.
Cobo, E. (1973). Effect of different dose of alcohol on the milk-ejecting reflex in lactating women. American Journal of Obstetrics and Gynecology, 115, 817.
Coiro, V. (1992). Inhibition by ethanol of the oxytocin response to breast stimulation in normal women and the role of endogenous opioids. Acta Endocrinologia, 126, 3–16.
Connelly, D. M., Unwin, J. W., & Taberner, P. V. (1983). The role of the blood glucose level in determining voluntary ethanol consumption in the LACG and diabetogenic C57BL strains of mice. Biochemical Pharmacology, 32, 221–6.
Cooper, D. S., Klibanski, A., & Ridgway, E. C. (1983). Dopaminergic modulation of TSH and its subunits: in vivo and in vitro studies. Clinical Endocrinology, 18, 265–275.
Corl, A. B., Rodan, A. R., & Heberlein, U. (2005). Insulin signaling in the nervous system regulates ethanol intoxication in Drosophila melanogaster. Nature Neuroscience, 8, 18–19.
Delitala, G., Devilla, L., Canessa, A., & D'Asta, F. (1981). On the role of dopamine receptors in the central regulation of human TSH. Acta Endocrinology, 98, 521–527.
Ditzen, B., Schaer, M., Gabriel, B., Bodenmann, G., Ehlert, U., & Heinrichs, M. (2009). Intranasal oxytocin increases positive communication and reduces cortisol levels during couple conflict. Biological Psychiatry, 65, 728–31.
Domes, G., Heinrichs, M., Gläscher, J., Büchel, C., Braus, D. F., & Herpertz, S. C. (2007). Oxytocin attenuates amygdala responses to emotional faces regardless of valence. Biological Psychiatry, 62, 1187–90.
Domes, G., Heinrichs, M., Michel, A., Berger, C., & Herpertz, S. C. (2007). Oxytocin improves “mind-reading” in humans. Biological Psychiatry, 61, 731–3.
Dorgan, J. F., Baer, D. J., Albert, P. S., Judd, J. T., Brown, E. D., Corle, D. K., et al. (2001). Serum hormones and the alcohol-breast cancer association in postmenopausal women. Journal of the National Cancer Institute, 93, 710–5.
Döring, W. K., Herzenstiel, M. N., Krampe, H., Jahn, H., Pralle, L., Sieg, S., et al. (2003). Persistent alterations of vasopressin and N-terminal proatrial natriuretic peptide plasma levels in long-term abstinent alcoholics. Alcoholism: Clinical and Experimental Research, 7, 849–61.
Dumitrescu, R. G., & Shields, P. G. (2005). The etiology of alcohol-induced breast cancer. Alcohol, 35, 213–25.
Ebner, K., Bosch, O. J., Kromer, S. A., Singewald, N., & Neumann, I. D. (2005). Release of oxytocin in the rat central amygdala modulates stress-coping behavior and the release of excitatory amino acids. Neuropsychopharmacology, 30, 223–230.
Edwards, S., Kenna, G. A., Swift, R. M., & Leggio, L. (2011). Current and promising pharmacotherapies, and novel research target areas in the treatment of alcohol dependence: a review. Current Pharmacological Design, 17, 1323–32.
Edwards, S., Guerrero, M., Ghoneim, O. M., Roberts, E., & Koob, G. F. (2012). Evidence that vasopressin V(1b) receptors mediate the transition to excessive drinking in ethanol-dependent rats. Addiction Biology, 17, 76–85.
Ehrenreich, H., tom Dieck, K., Gefeller, O., Kaw, S., Schilling, L., Poser, W., et al. (1997). Sustained elevation of vasopressin plasma levels in healthy young men, but not in abstinent alcoholics, upon expectation of novelty. Psychoneuroendocrinology, 22, 13–24.
Emanuele, M. A., & Emanuele, N. (2001). Alcohol and the male reproductive system. Alcohol Research and Health, 25, 282–7.
Engelmann, M., Ebner, K., Landgraf, R., Holsboer, F., & Wotjak, C. T. (1999). Emotional stress triggers intrahypothalamic but not peripheral release of oxytocin in male rats. Journal of Neuroendocrinology, 11, 867–872.
Enoch, M. A., Shen, P. H., Ducci, F., Yuan, Q., Liu, J., White, K. V., et al. (2008). Common genetic origins for EEG, alcoholism and anxiety: the role of CRH-BP. PLoS ONE, 3, e3620.
Eriksson, C. J., Kaprio, J., Pulkkinen, L., & Rose, R. J. (2005). Testosterone and alcohol use among adolescent male twins: testing between-family associations in within-family comparisons. Behavioral Genetics, 35, 359–68.
Etique, N., Chardard, D., Chesnel, A., Merlin, J. L., Flament, S., & Grillier-Vuissoz, I. (2004). Ethanol stimulates proliferation, ER alpha and aromatase expression in MCF-7 human breast cancer cells. International Journal of Molecular Medicine, 13, 149–55.
Evans, S. M., & Levin, F. R. (2011). Response to alcohol in women: role of the menstrual cycle and a family history of alcoholism. Drug and Alcohol Dependence, 114, 18–30.
Fernandez, S. V. (2011). Estrogen, alcohol consumption, and breast cancer. Alcoholism: Clinical and Experimental Research, 35, 389–91.
Figlewicz, D. P. (2003). Insulin, food intake, and reward. Sem Clinical Neuropsychiatry, 8, 82–93.
Franko, D. L., Dorer, D. J., Keel, P. K., Jackson, S., Manzo, M. P., & Herzog, D. B. (2008). Interactions between eating disorders and drug abuse. Journal of Nervous and Mental Disease, 196, 556–61.
Frias, J., Torres, J. M., Miranda, M. T., Ruiz, E., & Ortega, E. (2002). Effects of acute alcohol intoxication on pituitary-gonadal axis hormones, pituitary-adrenal axis hormones, beta-endorphin and prolactin in human adults of both sexes. Alcohol Alcoholism, 37, 169–73.
Fuchs, A. R., & Wagner, G. (1963). The effect of ethyl alcohol on the release of oxytocin in rabbits. Acta Endocrinology, 44, 593–605.
Galvão-Teles, A., Burke, C. W., Anderson, D. C., Marshall, J. C., Corker, C. S., Brown, et al. (1973). Biologically active androgens and oestradiol in men with chronic liver disease. Lancet, 27, 173–7.
Gamel-Didelon, K., Corsi, C., Pepeu, G., Jung, H., Gratzl, M., & Mayerhofer, A. (2002). An autocrine role for pituitary GABA: activation of GABA-B receptors and regulation of growth hormone levels. Neuroendocrinology, 76, 170–7.
Gamer, M., & Büchel, C. (2012). Oxytocin specifically enhances valence-dependent parasympathetic responses. Psychoneuroendocrinology, 37, 87–93.
Garbutt, J. C., Mayo, J. P., Little, K. Y., Gillette, G. M., Mason, G. A., Dew, B., et al. (1996). Dose-response studies with thyrotropin-releasing hormone: evidence for differential pituitary responses in men with major depression, alcoholism, or no psychopathology. Alcoholism: Clinical and Experimental Research, 20, 717–722.
Gavaler, J. S. (1988). Effects of moderate consumption of alcoholic beverages on endocrine function in postmenopausal women. Bases for hypotheses. Recent Developments in Alcoholk, 6, 229–51.
Gavaler, J. S. (1998). Alcoholic beverages as a source of estrogens. Alcohol Health Research World, 22, 220–7.
Gavaler, J. S., & Van Thiel, D. H. (1988). Gonadal dysfunction and inadequate sexual performance in alcoholic cirrhotic men. Gastroenterology, 95, 1680–3.
Gavaler, J. S., & Van Thiel, D. H. (1992a). The association between moderate alcoholic beverage consumption and serum estradiol and testosterone levels in normal postmenopausal women: relationship to the literature. Alcoholism: Clinical and Experimental Research, 16, 87–92.
Gavaler, J. S., & Van Thiel, D. H. (1992b). Hormonal status of postmenopausal women with alcohol-induced cirrhosis: Further findings and a review of the literature. Hepatology, 16, 312–319.
Gavaler, J. S., Rosenblum, E. R., Van Thiel, D. H., Eagon, P. K., Pohl, C. R., Campbell, I. M., et al. (1987). Biologically active phytoestrogens are present in bourbon. Alcoholism: Clinical and Experimental Research, 11, 399–406.
Gessa, G. L., Muntoni, F., Collu, M., Vargiu, L., & Mereu, G. (1985). Low doses of ethanol activate dopaminergic neurons in the ventral tegmental area. Brain Research, 348, 201–3.
Gilman, J. M., Ramchandani, V. A., Davis, M. B., Bjork, J. M., & Hommer, D. W. (2008). Why we like to drink: an fMRI Study of the rewarding and anxiolytic effects of alcohol. Journal of Neuroscience, 28, 4583–4591.
Goodman, H. M. (2003). Basic medical endocrinology (2nd ed.). Burlington: Elsevier.
Gordon, G. G., & Lieber, C. S. (1992). Alcohol, hormones and metabolism. In C. S. Lieber (Ed.), Medical and nutritional complications of alcoholism (pp. 55–90). New York: Plenum.
Gordon, G. G., Southren, A. L., Vittek, J., & Lieber, C. S. (1979). Effect of alcohol ingestion on hepatic aromatase activity and plasma steroid hormones in the rat. Metabolism, 28, 20–24.
Guastella, A. J., Howard, A. L., Dadds, M. R., Mitchell, P., & Carson, D. S. (2009). A randomized controlled trial of intranasal as an adjunct to exposure therapy for social anxiety disorder. Psychoneuroendocrinology, 34, 917–23.
Hankinson, S. E., Manson, J. E., Spiegelman, D., Willett, W. C., Longcope, C., & Speizer, F. E. (1995). Reproducibility of plasma hormone levels in postmenopausal women over a 2-3-year period. Cancer Epidemiological Biomarkers & Prevention, 4, 649–54.
Heil, S. H., & Subramanian, M. G. (1998). Alcohol and the hormonal control of lactation. Alcohol Health Research World, 22, 178–84.
Heilig, M., Goldman, D., Berrettini, W., & O'Brien, C. P. (2011). Pharmacogenetic approaches to the treatment of alcohol addiction. Nature ReviewNeuroscience, 12, 670–84.
Heinrichs, M., Meinlschmidt, G., Wippich, W., Ehlert, U., & Hellhammer, D. H. (2004). Selective amnesic effects of oxytocin on human memory. Physiology and Behavior, 83, 31–8.
Heinz, A., Siessmeier, T., Wrase, J., Hermann, D., Klein, S., Grüsser, S. M., et al. (2004). Correlation between dopamine D(2) receptors in the ventral striatum and central processing of alcohol cues and craving. American Journal of Psychiatry, 161, 1783–1789.
Hermann, D., Heinz, A., & Mann, K. (2002). Dysregulation of the hypothalamic–pituitary–thyroid axis in alcoholism. Addiction, 97, 1369–1381.
Hillemacher, T., Bayerlein, K., Wilhelm, J., Reulbach, U., Frieling, H., Bönsch, D., et al. (2005). Alteration of prolactin serum levels during alcohol withdrawal correlates with craving in female patients. Addiction Biology, 10, 337–43.
Hillemacher, T., Bayerlein, K., Wilhelm, J., Poleo, D., Frieling, H., Ziegenbein, M., et al. (2006). Volume intake and craving in alcohol withdrawal. Alcohol Alcoholism, 41, 61–5.
Hillemacher, T., Bleich, S., Frieling, H., Schanze, A., Wilhelm, J., Sperling, W., et al. (2007). Evidence of an association of leptin serum levels and craving in alcohol dependence. Psychoneuroendocrinology, 32, 87–90.
Hillemacher, T., Kraus, T., Rauh, J., Weiss, J., Schanze, A., Frieling, H., et al. (2007). Role of appetite-regulating peptides in alcohol craving: an analysis in respect to subtypes and different consumption patterns in alcoholism. Alcoholism: Clinical and Experimental Research, 31, 950–4.
Hillemacher, T., Bayerlein, K., Frieling, H., Wilhelm, J., Ziegenbein, M., Kornhuber, J., et al. (2007). Elevated prolactin serum levels and history of alcohol withdrawal seizures. Journal on Psychiatric Research, 41, 702–6.
Hillemacher, T., Frieling, H., Luber, K., Yazici, A., Muschler, M. A., Lenz, B., et al. (2009). Epigenetic regulation and gene expression of vasopressin and atrial natriuretic peptide in alcohol withdrawal. Psychoneuroendocrinology, 34, 555–60.
Hillemacher, T., Weinland, C., Heberlein, A., Gröschl, M., Schanze, A., Frieling, H., et al. (2009). Increased levels of adiponectin and resistin in alcohol dependence-possible link to craving. Drug and Alcohol Dependence, 99, 333–337.
Hillemacher, T., Kahl, K. G., Heberlein, A., Muschler, M. A., Eberlein, C., Frieling, H., et al. (2010). Appetite- and volume-regulating neuropeptides: role in treating alcohol dependence. Current Opinion Investigating Drugs, 11, 1097–106.
Hoge, E. A., Pollack, M. H., Kaufman, R. E., Zak, P. J., & Simon, N. M. (2008). Oxytocin levels in social anxiety disorder. CNS Neuroscience & Therapeutics, 14, 165–170.
Hurlemann, R., Patin, A., Onur, O. A., Cohen, M. X., Baumgartner, T., Metzler, S., et al. (2010). Oxytocin enhances amygdala-dependent, socially reinforced learning and emotional empathy in humans. Journal of Neuroscience, 30, 4999–5007.
Hyytia, P., & Koob, G. (1995). GABAA receptor antagonism in the extended amygdala decreases ethanol self-administration in rats. European Journal of Pharmacology, 283, 151–159.
Imperato, A., & Di Chiara, G. (1986). Preferential stimulation of dopamine release in the nucleus accumbens of freely moving rats by ethanol. Journal of Pharmacology Experimental Therapy, 239, 219–28.
Inui, A. (1999). Feeding and body-weight regulation by hypothalamic neuropeptides—mediation of the actions of leptin. Trends in Neuroscience, 2, 62–67.
Irvine, R. A., Ma, H., Yu, M. C., Ross, R. K., Stallcup, M. R., & Coetzee, G. A. (2000). Inhibition of p160-mediated coactivation with increasing androgen receptor polyglutamine length. Human Molecular Genetics, 9, 267–74.
Israel, S., Lerer, E., Shalev, I., Uzefovsky, F., Reibold, M., Bachner-Melman, R., et al. (2008). Molecular genetic studies of the arginine vasopressin 1a receptor (AVPR1a) and the oxytocin receptor (OXTR) in human behaviour: from autism to altruism with some notes in between. Progress in Brain Research, 170, 435–449.
Jahn, H., Döring, W. K., Krampe, H., Sieg, S., Werner, C., Poser, W., et al. (2004). Preserved vasopressin response to osmostimulation despite decreased basal vasopressin levels in long-term abstinent alcoholics. Alcoholism: Clinical and Experimental Research, 28, 1925–30.
Jerlhag, E. (2008). Systemic administration of ghrelin induces conditioned place preference and stimulates accumbal dopamine. Addiction Biology, 13, 358–363.
Jerlhag, E., Egecioglu, E., Dickson, S. L., Douhan, A., Svensson, L., & Engel, J. A. (2007). Ghrelin administration into tegmental areas stimulates locomotor activity and increases extracellular concentration of dopamine in the nucleus accumbens. Addiction Biology, 12, 6–16.
Jerlhag, E., Egecioglu, E., Dickson, S. L., Svensson, L., & Engel, J. A. (2008). Alpha-conotoxin MII-sensitive nicotinic acetylcholine receptors are involved in mediating the ghrelin-induced locomotor stimulation and dopamine overflow in nucleus accumbens. European Neuropsychopharmacology, 18, 508–518.
Jerlhag, E., Egecioglu, E., Landgren, S., Salomé, N., Heilig, M., Moechars, D., et al. (2009). Requirement of central ghrelin signaling for alcohol reward. Procedures of the National Academy of Science, U S A, 106, 11318–23.
Karagiannis, A., & Harsoulis, F. (2005). Gonadal dysfunction in systemic diseases. European Journal of Endocrinology, 152, 501–13.
Kenna, G. A., McGeary, J. E., & Swift, R. M. (2004). Pharmacotherapy, pharmacogenomics and the future of alcohol dependence therapy. American Journal of Health-System Pharmacy, 61(Part 2), 2380–2390.
Kenna, G. A., Hanna-Roder, N., Leggio, L., Zywiak, W. H., Clifford, J., Edwards, S., et al. (2012). The association of the 5-HTT gene-linked promoter region (5-HTTLPR) polymorphism to psychiatry: a review of psychopathology and pharmacotherapy. Pharmacogenomics Personalized Medicine, 5, 19–35.
Kiefer, F., Jahn, H., Jaschinski, M., Holzbach, R., Wolf, K., Naber, D., et al. (2001). Leptin: a modulator of alcohol craving? Biological Psychiatry, 49, 782–787.
Kiefer, F., Jahn, H., Wolf, K., Kämpf, P., Knaudt, K., & Wiedemann, K. (2001). Free-choice alcohol consumption in mice after application of the appetite regulating peptide leptin. Alcoholism: Clinical and Experimental Research, 25, 787–789.
Kiefer, F., Andersohn, F., Jahn, H., Wolf, K., Raedler, T. J., & Wiedemann, K. (2002). Involvement of plasma atrial natriuretic peptide in protracted alcohol withdrawal. Acta Psychiatric Scandinavian, 105, 65–70.
Kiefer, F., Jahn, H., Otte, C., Demiralay, C., Wolf, K., & Wiedemann, K. (2005). Increasing leptin precedes craving and relapse during pharmacological abstinence maintenance treatment of alcoholism. Journal of Psychiatric Research, 39, 545–451.
Kim, D. J., Yoon, S. J., Choi, B., Kim, T. S., Woo, Y. S., Kim, W., et al. (2005). Increased fasting plasma ghrelin levels during alcohol abstinence. Alcohol Alcoholism, 40, 76–79.
Kirsch, P., Esslinger, C., Chen, Q., Mier, D., Lis, S., & Siddhanti, S. (2005). Oxytocin modulates neural circuitry for social cognition and fear in humans. Journal of Neuroscience, 25, 11489–11493.
Knott, T. K., Dopico, A. M., Dayanithi, G., Lemos, J., & Treistman, S. N. (2002). Integrated channel plasticity contributes to alcohol tolerance in neurohypophysial terminals. Molecular Pharmacology, 62, 135–142.
Kojima, M., Hosoda, H., Date, Y., Nakazato, M., Matsuo, H., & Kangawa, K. (1999). Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature, 402, 656–660.
Koob, G. F. (2009). Brain stress systems in the amygdala and addiction. Brain Res., 1293, 61–75.
Koob, G. F., & Le Moal, M. (1997). Drug abuse: hedonic homeostatic dysregulation. Science, 278, 52–8.
Koopmann, A., von der Goltz,C., Grosshans,M., Dinter,C., Vitale,M., Wiedemann, K., et al. (2011). The association of the appetitive peptide acetylated ghrelin with alcohol craving in early abstinent alcohol dependent individuals. Psychoneuroendocrinology, [Epub ahead of print]
Kosfeld, M., Heinrichs, M., Zak, P. J., Fischbacher, U., & Fehr, E. (2005). Oxytocin increases trust in humans. Nature, 435, 673–6.
Kostoglou-Athanassiou, I., Treacher, D. F., & Forsling, M. L. (1994). Is oxytocin naturetic in man? J Endocrinology, 143(suppl O), 39.
Kovács, G. L., & Telegdy, G. (1987). Endorphin tolerance is inhibited by oxytocin. Pharmacology Biochemical Behavior, 26, 57–60.
Kovács, G. L., Sarnyai, Z., & Szabó, G. (1998). Oxytocin and addiction: a review. Psychoneuroendocrinology, 23(8), 945–62.
Kranzler, H. R., & Edenberg, H. J. (2010). Pharmacogenetics of alcohol and alcohol dependence treatment. Current Pharmacological Designs, 16, 2141–8.
Kranzler, H. R., Burleson, J. A., Del Boca, F. K., Babor, T. F., Korner, P., Brown, J., et al. (1994). Buspirone treatment of anxious alcoholics. A placebo-controlled trial. Archives of General Psychiatry, 51, 720–31.
Kraus, T., Reulbach, U., Bayerlein, K., Mugele, B., Hillemacher, T., Sperling, W., et al. (2004). Leptin is associated with craving in females with alcoholism. Addiction Biology, 9, 213–219.
Kraus, T., Schanze, A., Gröschl, M., Bayerlein, K., Hillemacher, T., Reulbach, U., et al. (2005). Ghrelin levels are increased in alcoholism. Alcoholism: Clinical and Experimental Research, 29, 2154–2157.
Kumar, S., Porcu, P., Werner, D. F., Matthews, D. B., Diaz-Granados, J. L., Helfand, R. S., et al. (2009). The role of GABAA receptors in the acute and chronic effects of ethanol: a decade of progress. Psychopharmacology (Berl), 205, 529–564.
La Grange, L., Jones, T. D., Erb, L., & Reyes, E. (1995). Alcohol consumption: biochemical and personality correlates in a college student population. Addiction Behavior, 20, 93–103.
Landgraf, R. (2005). Neuropeptides in anxiety modulation. Handbook Experimental Pharmacology, 169, 335–69.
Landgraf, R., & Neumann, I. D. (2004). Vasopressin and oxytocin release within the brain: a dynamic concept of multiple and variable modes of neuropeptide communication. Frontiers in Neuroendocrinology, 25, 150–176.
Leggio, L. (2009). Understanding and treating alcohol craving and dependence: recent pharmacological and neuroendocrinological findings. Alcohol Alcoholism, 44, 341–352.
Leggio, L. (2010). Role of the ghrelin system in alcoholism: acting on the growth hormone secretagogue receptor (GHS-R) to treat alcohol-related disorders. Drug News Perspect., 23, 157–166.
Leggio, L., Ferrulli, A., Malandrino, N., Miceli, A., Capristo, E., Gasbarrini, G., et al. (2008). Insulin but not insulin growth factor-1 correlates with craving in currently drinking alcohol-dependent patients. Alcoholism: Clinical and Experimental Research, 32, 450–458.
Leggio, L., Ferrulli, A., Cardone, S., Malandrino, N., Mirijello, A., D'Angelo, C., et al. (2008). Relationship between the hypothalamic-pituitary-thyroid axis and alcohol craving in alcohol-dependent patients: a longitudinal study. Alcoholism: Clinical and Experimental Research, 32, 2047–2053.
Leggio, L., Kenna, G. A., Fenton, M., Bonenfant, E., & Swift, R. M. (2009). Typologies of alcohol dependence. From Jellinek to genetics and beyond. Neuropsychology Review., 19, 115–129.
Leggio, L., Ray, L. A., Kenna, G. A., & Swift, R. M. (2009). Blood glucose level, alcohol heavy drinking, and alcohol craving during treatment for alcohol dependence: results from the Combined Pharmacotherapies and Behavioral Interventions for Alcohol Dependence (COMBINE) Study. Alcoholism: Clinical and Experimental Research, 33, 1539–1544.
Leggio, L., Ferrulli, A., Cardone, S., Nesci, A., Miceli, A., Malandrino, N., et al. (2011). Ghrelin system in alcohol-dependent subjects: role of plasma ghrelin levels in alcohol drinking and craving. Addict Biol. doi:10.1111/j.1369-1600.2010.00308.x. epub pub ahead of print.
Leggio, L., Addolorato, G., Cippitelli, A., Jerlhag, E., Kampov-Polevoy, A. B., & Swift, R. M. (2011). Role of feeding-related pathways in alcohol dependence: a focus on sweet preference, NPY, and ghrelin. Alcoholism: Clinical and Experimental Research, 35, 194–202.
Leggio, L., Kenna, G., Zywiak, W., Edwards, S., Fricchione, S., & Tavares, T. (2011). Baclofen as a novel pharmacotherapy for alcohol dependence: preliminary findings from a human laboratory double-blind placebo-controlled randomized study. Neuropsychopharmacology, 36(Supplement 1s), S428.
Lenz, B., Jacob, C., Frieling, H., Jacobi, A., Hillemacher, T., Muschler, M., et al. (2009). Polymorphism of the long polyglutamine tract in the human androgen receptor influences craving of men in alcohol withdrawal. Psychoneuroendocrinology, 34, 968–71.
Lenz, B., Frieling, H., Jacob, C., Heberlein, A., Kornhuber, J., Bleich, S., et al. (2010). The modulating effect of the androgen receptor on craving in alcohol withdrawal of men is partially mediated by leptin. Pharmacogenomics Journal., 10, 226–31.
Lenz, B., Muller, C. P., Stoessel, C., Sperling, W., Biermann, T., Hillemacher, T., et al. (2012). Sex hormone activity in alcohol addiction: integrating organizational and activational effects. Progress in Neurobiology, 96, 136–163.
Lesch, O. M., Kefer, J., Lentner, S., Mader, R., Marx, B., Musalek, M., et al. (1990). Diagnosis of chronic alcoholism–classificatory problems. Psychopathology, 23, 88–96.
Longcope, C. (1999). The endocrinology of the menopause. In R. A. Lobo (Ed.), Treatment of the postmenopausal woman: basic and clinical aspects (2nd ed., pp. 35–40). Philadelphia: Lippincott.
Marchesi, C., Chiodera, P., Brusamonti, E., Volpi, R., & Cioro, V. (1997). Abnormal plasma oxytocin and beta-endorphin levels in alcoholics after short and long term abstinence. Prog Neuro-Psychopharmacol & Biol Psychiatry, 21, 797–807.
McGregor, I. S., Callaghan, P. D., & Hunt, G. E. (2008). From ultrasocial to antisocial: a role for oxytocin in the acute reinforcing effects and long-term adverse consequences of drug use? Br J Pharmacology, 154, 358–68.
Melis, M. R., Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., et al. (2007). Oxytocin injected into the ventral tegmental area induces penile erection and increases extracellular dopamine in the nucleus accumbens and paraventricular nucleus of the hypothalamus of male rats. European Journal of Neuroscience., 26, 1026–1035.
Mendelson, J. H., & Mello, N. K. (1988). Chronic alcohol effects on anterior pituitary and ovarian hormones in healthy women. Journal of Pharmacology and Experimental Therapeautics, 245, 407–12.
Mennella, J. A., Pepino, M. Y., & Teff, K. L. (2005). Acute alcohol consumption disrupts the hormonal milieu of lactating women. Journal of Clinical Endocrinological Metabolism, 90, 1979–85.
Merikangas, K. R., Mehta, R. L., Molnar, B. E., Walters, E. E., Swendsen, J. D., Aguilar-Gaziola, S., et al. (1998). Comorbidity of substance use disorders with mood and anxiety disorders: results of the International Consortium in Psychiatric Epidemiology. Addiction Behavior, 23, 893–907.
Messini, C. I., Dafopoulos, K., Chalvatzas, N., Georgoulias, P., Anifandis, G., & Messinis, I. E. (2010a). Effect of ghrelin and thyrotropin-releasing hormone on prolactin secretion in normal women. Hormones and Metabolism Research, 42, 204–8.
Messini, C. I., Dafopoulos, K., Chalvatzas, N., Georgoulias, P., Anifandis, G., & Messinis, I. E. (2010b). Blockage of ghrelin-induced prolactin secretion in women by bromocriptine. Fertil Steril, 94, 1478–81.
Miller, F., Barasch, A., Sacks, M., Levitan, J., & Ashcroft, L. (1986). Serum prolactin correlates with depressed mood during alcohol withdrawal. Drug and Alcohol Dependence, 17, 331–338.
Monti, P. M., Gulliver, S. B., & Myers, M. G. (1994). Social skills training for alcoholics: assessment and treatment. Alcohol and Alcoholism, 29, 627–637.
Murphy, V. E., Smith, R., Giles, W. B., & Clifton, V. L. (2006). Endocrine regulation of human fetal growth: the role of the mother, placenta, and fetus. Endocrinology Review, 27, 141–69.
Nahta, R., Shabaya, S., Ozbay, T., & Rowe, D. L. (2009). Personalizing HER2-targeted therapy in metastatic breast cancer beyond HER2 status: what we have learned from clinical specimens. Current Pharmacogenomics and Personalized Medicine, 7, 263–274.
Neumann, I. D. (2007). Stimuli and consequences of dendritic release of oxytocin within the brain. Biochem Soc Trans, 35, 1252–1257.
Nicolás, J. M., Fernández-Solà, J., Fatjó, F., Casamitjana, R., Bataller, R., Sacanella, E., et al. (2001). Increased circulating leptin levels in chronic alcoholism. Alcoholism: Clinical and Experimental Research, 25, 83–8.
Ohashi, M., Fujio, N., Nawata, H., Kato, K., Matsubayashi, S., Tamai, H., et al. (1988). Human atrial natriuretic polypeptide in plasma of patients with anorexia nervosa. Hormone Metabolism Research, 20, 705–8.
Oscar-Berman, M., & Marinković, K. (2007). Alcohol: effects on neurobehavioral functions and the brain. Neuropsychology Review, 17, 239–57.
Ott, I., & Scott, J. C. (2010). The action of infundibulin upon the mammary secretion. Proceeds of the Society on Experimental and Biological Medicine, 8, 48–49.
Ozsoy, S., Esel, E., Turan, T., & Kula, M. (2007). Growth hormone response to the GABA-B agonist baclofen in 3-week abstinent alcoholics. Alcohol, 41, 551–6.
Pandey, S. C., Zhang, H., Roy, A., & Xu, T. (2005). Deficits in amygdaloid cAMP-responsive element–binding protein signaling may play a role in genetic predisposition to anxiety and alcoholism. Journal Clinical Investigation., 115, 2762–2773.
Perry, B. L., Miles, D., Burruss, K., & Svikis, D. S. (2004). Premenstrual symptomatology and alcohol consumption in college women. Journal of Studies on Alcohol, 65, 464–8.
Pijl, H., de Meijer, P. H., Langius, J., Coenegracht, C. I., van den Berk, A. H., Chandie Shaw, P. K., et al. (2001). Food choice in hyperthyroidism: potential influence of the autonomic nervous system and brain serotonin precursor availability. Journal of Clinical Endocrinology & Metabolism, 86, 5848–5853.
Pristupa, Z. B., Wilson, J. M., Hoffman, B. J., Kish, S. J., & Niznik, H. B. (1994). Pharmacological heterogeneity of the cloned and native human dopamine transporter: disassociation of [3 H] WIN 35,428 and [3 H] GBR 12,935 binding. Molecular Pharmacology, 45, 125–135.
Qi, Y., Takahashi, N., Hileman, S. M., Patel, H. R., Berg, A. H., Pajvani, U. B., et al. (2004). Adiponectin acts in the brain to decrease body weight. Nature Medicine, 10, 524–529.
Raggenbass, M. (2008). Overview of cellular electrophysiological actions of vasopressin. European Journal of Pharmacology, 583, 243–54.
Ring, R. H. (2011). A complicated picture of oxytocin action in the central nervous system revealed. Biological Psychiatry, 69, 818–819.
Rosenblum, E. R., Van Thiel, D. H., Campbell, I. M., & Gavaler, J. S. (1991). Quantitation of beta-sitosterol in bourbon. Alcoholism: Clinical and Experimental Research, 15, 205–6.
Ross, H. E., Freeman, S. M., Spiegel, L. L., Ren, X., Terwillige, E. F., & Young, L. J. (2009). Variation in oxytocin receptor density in the nucleus accumbens has differential effects on affiliative behaviors in monogamous and polygamous voles. Journal of Neuroscience, 29, 1312–1318.
Ruusa, J., Bergman, B., & Sundell, M. L. (1997). Sex hormones during alcohol withdrawal: a longitudinal study of 29 male alcoholics during detoxification. Alcohol Alcoholism, 32, 591–7.
Sakharkar, A. J., Zhang, H., Tang, L., Shi, G., & Pandey, S. C. (2011). Histone Deacetylases (HDAC)-induced histone modifications in the amygdala: a role in rapid tolerance to the anxiolytic effects of ethanol alcoholism. Clinical and Experimental Research, 35, 1–11.
Salvante, K. G., Brindle, E., McConnell, D., O'connor, K., & Nepomnaschy, P. A. (2012). Validation of a new multiplex assay against individual immunoassays for the quantification of reproductive, stress, and energetic metabolism biomarkers in urine specimens. American Journal of Human Biology, 24, 81–6.
Santolaria, F., Perez-Cejas, A., Aleman, M. R., González-Reimers, E., Milena, A., de la Vega, M. J., et al. (2003). Low serum leptin levels and malnutrition in chronic alcohol misusers hospitalized by somatic complications. Alcohol Alcoholism, 38, 60–66.
Sarkola, T., Mäkisalo, H., Fukunaga, T., & Eriksson, C. J. (1999). Acute effect of alcohol on estradiol, estrone, progesterone, prolactin, cortisol, and luteinizing hormone in premenopausal women. Alcoholism: Clinical and Experimental Research, 23, 976–82.
Sarnyai, Z., & Kovács, G. L. (1994). Role of oxytocin in the neuroadaptation to drugs of abuse. Psychoneuroendocrinology, 19, 85–117.
Scantamburlo, G., Hansenne, M., Fuchs, S., Pitchot, W., Marechal, P., Pequeux, C., et al. (2007). Plasma oxytocin levels and anxiety in patients with major depression. Psychoneuroendocrinology, 32, 407–410.
Schuckit, M. A. (1994). Low level of response to alcohol as a predictor of future alcoholism. American Journal of Psychiatry, 151, 184–189.
Schuckit, M. A. (1996). Alcohol, anxiety and depressive disorders. Alcohol Health Res World., 20, 81–85.
Schuckit, M. A., Gold, E., & Risch, C. (1987). Serum prolactin levels in sons of alcoholics and control subjects. American Journal of Psychiatry, 144, 854–9.
Seidman, S. N., Araujo, A. B., Roose, S. P., & McKinlay, J. B. (2001). Testosterone level, androgen receptor polymorphism, and depressive symptoms in middle-aged men. Biological Psychiatry, 50, 371–6.
Shahrokh, D. K., Zhang, T. Y., Diorio, J., Gratton, A., & Meaney, M. J. (2010). Oxytocin-dopamine interactions mediate variations in maternal behavior in the rat. Endocrinology, 151, 2276–86.
Short, R. V. (1984). Breastfeeding. Scientific American., 250, 35–41.
Silva, S. M., Madeira, M. D., Ruela, C., & Paula-Barbosa, M. M. (2002). Prolonged alcohol intake leads to irreversible loss of vasopressin and oxytocin neurons in the paraventricular nucleus of the hypothalamus. Brain Research, 925, 76–88.
Singletary, K. W., & Gapstur, S. M. (2001). Alcohol and breast cancer: review of epidemiologic and experimental evidence and potential mechanisms. Journal of the American Medical Association., 286, 2143–51.
Smith, J. J. (1950). The treatment of acute alcoholic states with ACTH and adrenocortical hormones. Quarterly Journal of Studies on Alcohol., 11, 190–8.
Smith-Warner, S. A., Spiegelman, D., Yaun, S. S., van den Brandt, P. A., Folsom, A. R., Goldbohm, R. A., et al. (1998). Alcohol and breast cancer in women: a pooled analysis of cohort studies. Journal of the American Medical Association., 279, 535–40.
Ströhle, A., & Holsboer, F. (2003). Stress responsive neurohormones in depression and anxiety. Pharmacopsychiatry, 36(Suppl 3), S207–14.
Ströhle, A., Feller, C., Strasburger, C. J., Heinz, A., & Dimeo, F. (2006). Anxiety modulation by the heart? Aerobic exercise and atrial natriuretic peptide. Psychoneuroendocrinology, 31, 1127–30.
Sturgess, J. E., George, T. P., Kennedy, J. L., Heinz, A., & Müller, D. J. (2011). Pharmacogenetics of alcohol, nicotine and drug addiction treatments. Addiction Biology, 16, 357–76.
Sullivan, E. V. (2007). Alcohol and drug dependence: brain mechanisms and behavioral impact. Neuropsychology Review, 17, 235–8.
Surget, A., & Belzung, C. (2008). Involvement of vasopressin in affective disorders. European Journal Pharmacology, 583, 340–9.
Suzuki, R., Allen, N. E., Appleby, P. N., Key, T. J., Dossus, L., Tjønneland, A., et al. (2009). Lifestyle factors and serum androgens among 636 middle aged men from seven countries in the European Prospective Investigation into Cancer and Nutrition (EPIC). Cancer Causes Control., 20, 811–21.
Suzuki, K., Jayasena, C. N., & Bloom, S. R. (2011). The gut hormones in appetite regulation. Journal of Obesity, 2011, 528401.
Svikis, D. S., Miles, D. R., Haug, N. A., Perry, B., Hoehn-Saric, R., & McLeod, D. (2006). Premenstrual symptomatology, alcohol consumption, and family history of alcoholism in women with premenstrual syndrome. Journal of Studies on Alcohol., 67, 833–6.
Szabó, G., Kovács, G. L., Székeli, S., & Telegdy, G. (1985). The effects of neurohypophyseal hormones on tolerance to the hypothermic effect of ethanol. Alcohol., 2, 567–674.
Tabakoff, B., & Hoffman, P. L. (1988). Tolerance and the etiology of alcoholism: hypothesis and mechanism. Alcoholism: Clinical and Experimental Research, 12, 184–186.
Tan, G. H., Choo, W. Y., Taib, N. A., & Yip, C. H. (2009). Factors associated with HER2 overexpression in breast cancer: experience in an Asian developing country. Asian Pacific Journal Cancer Prevention, 10, 837–40.
Tessner, K. D., & Hill, S. Y. (2010). Neural circuitry associated with risk for alcohol use disorders. Neuropsychology Review, 20, 1–20.
Tut, T. G., Ghadessy, F. J., Trifiro, M. A., Pinsky, L., & Yong, E. L. (1997). Long polyglutamine tracts in the androgen receptor are associated with reduced trans-activation, impaired sperm production, and male infertility. Journal of Clinical Endocrinology & Metabolism, 82, 3777–82.
Uvnäs-Moberg, K. (1996). Neuroendocrinology of the mother-child interaction. Trends Endocrinological Metabolism, 7, 126–131.
Välimäki, M., Pelkonen, R., Härkönen, M., & Ylikahri, R. (1984). Hormonal changes in noncirrhotic male alcoholics during ethanol withdrawal. Alcohol Alcoholism, 19, 235–242.
Van Thiel, D. H., Galvao-Teles, A., Monteiro, E., Rosenblum, E., & Gavaler, J. S. (1991). The phytoestrogens present in de-ethanolized bourbon are biologically active: a preliminary study in a postmenopausal woman. Alcoholism: Clinical and Experimental Research, 15, 822–3.
Vassaf, E. G., & Hall, V. R. (1946). The tonic effects of insulin in acute alcoholism. New England Journal of Medicine, 235, 190–3.
Vatsalya, V., Issa, J. E., Hommer, D. W., & Ramchandani, V. A. (2012). Pharmacodynamic effects of intravenous alcohol on hepatic and gonadal hormones: influence of age and sex. Alcoholism: Clinical and Experimental Research, 36, 207–13.
Veening, J. G., de Jong, T., & Barendregt, H. P. (2010). Oxytocin-messages via the cerebrospinal fluid: behavioral effects; a review. Physiology & Behavior, 101, 193–210.
Vescovi, P. P., & Coiro, V. (2001). Different control of GH secretion by gamma-amino- and gamma-hydroxy-butyric acid in 4-year abstinent alcoholics. Drug and Alcohol Dependence., 61, 217–21.
Vescovi, P. P., Volpi, R., & Coiro, V. (1998). Alcoholism abolishes the gamma-aminobutyric acid (GABA)ergic control of GH secretion in humans. Alcohol, 16, 325–8.
Volkow, N. D., & Wise, R. A. (2005). How can drug addiction help us understand obesity? Nature Neuroscience, 8, 555–560.
Volkow, N. D., Wang, G. J., Maynard, L., Fowler, J. S., Jayne, B., Telang, F., et al. (2002). Effects of alcohol detoxification on dopamine D2 receptors in alcoholics: a preliminary study. Psychiatry Research, 116, 163–72.
Volkow, N. D., Wang, G. J., Fowler, J. S., & Tomasi, D. (2012). Addiction circuitry in the human brain. Annual Review of Pharmacology and Toxicology, 52, 321–36.
Volpi, R., Chiodera, P., Gramellini, D., Cigarini, C., Papadia, C., Caffarri, G., et al. (1994). Endogenous opioid mediation of the inhibitory effect of ethanol on the prolactin response to breast stimulation in normal women. Life Science, 54, 739–44.
Weiss, F., Lorang, M.T., Bloom, F. E., Koob, G. F. (1993). Oral alcohol self-administration stimulates dopamine release in the rat nucleus accumbens: genetic and motivational determinants. Journal of Pharmacology and Experimental Therapeutics, 267, 250–8.
Westberg, L., Baghaei, F., Rosmond, R., Hellstrand, M., Landén, M., Jansson, M., et al. (2001). Polymorphisms of the androgen receptor gene and the estrogen receptor beta gene are associated with androgen levels in women. Journal of Clinical Endocrinological Metabolism, 86, 2562–8.
Wilhelm, J., Heberlein, A., Karagülle, D., Gröschl, M., Kornhuber, J., Riera, R., et al. (2011). Prolactin serum levels during alcohol withdrawal are associated with the severity of alcohol dependence and withdrawal symptoms. Alcoholism: Clinical and Experimental Research, 35, 235–9.
Willinger, U., Lenzinger, E., & Hornik, K. (2002). Anxiety as a predictor of relapse in detoxified alcohol-dependent patients. Alcohol Alcoholism, 37, 609–612.
Wong, A. W., Dunlap, S. M., Holcomb, V. B., & Nunez, N. P. (2011). Alcohol promotes mammary tumor development via the estrogen pathway in estrogen receptor alpha-negative HER2/neu mice. Alcoholism: Clinical and Experimental Research. doi:10.1111/j.1530-0277.2011.01654.x.
Wurst, F. M., Rasmussen, D. D., Hillemacher, T., Kraus, T., Ramskogler, K., Lesch, O., et al. (2007). Alcoholism, craving, and hormones: the role of leptin, ghrelin, prolactin, and the pro-opiomelanocortin system in modulating ethanol intake. Alcoholism: Clinical and Experimental Research, 31, 1963–7.
Zak, P. J., Stanton, A. A., & Ahmadi, S. (2007). Oxytocin increases generosity in humans. PLoS ONE, 2, e1128.
Zhang, S. M., Lee, I. M., Manson, J. E., Cook, N. R., Willett, W. C., & Buring, J. E. (2007). Alcohol consumption and breast cancer risk in the Women’s Health Study. American Journal of Epidemiology, 165, 667–76.
Zimmermann, U. S., Buchmann, A., Steffin, B., Dieterle, C., & Uhr, M. (2007). Alcohol administration acutely inhibits ghrelin secretion in an experiment involving psychosocial stress. Addiction Biology, 12, 17–21.
Zito, K. A., Vickers, G., Telford, L., & Roberts, D. C. (1984). Experimentally induced glucose intolerance increases oral ethanol intake in rats. Alcohol, 1, 257–61.
Disclosures
Drs. Kenna and Swift disclose that they are paid consultants to Laboratorio CT Sanremo, Sanremo, IT.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kenna, G.A., Swift, R.M., Hillemacher, T. et al. The Relationship of Appetitive, Reproductive and Posterior Pituitary Hormones to Alcoholism and Craving in Humans. Neuropsychol Rev 22, 211–228 (2012). https://doi.org/10.1007/s11065-012-9209-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11065-012-9209-y