
Abstract
Maternal stress can disturb normal fetal neurodevelopmental progress, and lead to negative behavioral and neuroendocrine consequences for the offspring. These effects may be related to alterations in the hypothalamic–pituitary–adrenal (HPA) axis. Early life events disrupting the function of the HPA axis may be associated with epigenetic modification. This study investigated the effect of maternal stress on the methylation rate of the corticotrophin-releasing hormone (CRH) promoter and HPA axis response to acute stress in the adolescent offspring of Sprague–Dawley rats. Pregnant dams were randomly assigned to two groups: restraint stress group and normal control group. Adolescent male and female offspring were used from each group. The results showed that prenatal stress is associated with the demethylation of the CRH promoter, and leads to anxiety-like behaviors in adolescent life stages, as well as hyper-responsiveness of the HPA axis. Together, these results imply that prenatal stress alters the normal HPA function, which may be via the epigenetic mechanism.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
During pregnancy, stress can have a negative effect on the offspring as a result of neuroendocrine disturbances [1–3]. Several clinical and preclinical studies have shown that exposure to maternal mood disorders, maternal stress and other forms of maternal hardship can affect the developing hypothalamic–pituitary–adrenal (HPA) axis [4–8]. Many studies have demonstrated that early life experiences, including maternal stress, are linked to anxiety and depressed behavior in adolescence and adulthood, in parallel with hyperactivity of the HPA axis [9–13].
In rodents, prenatal restraint stress (PRS) has been found to be associated with HPA axis alterations in the offspring. The long-term function of the HPA axis is affected in the PRS offspring, resulting in a prolonged corticosterone–stress response, and reduced levels of both mineralocorticoid and glucocorticoid receptors in the hippocampus in the adolescent and adult [2]. PRS offspring rats also show an increase in anxiety and depression-related behavior, as well as hyper-responsiveness of the HPA axis [14].
With regard to the question of how early life events disturb HPA axis function, most research has focused on epigenetic mechanisms. Several studies have confirmed that early childhood experiences are associated with the epigenetic modification of a neuron-specific exon I7 glucocorticoid receptor (NR3C1) promoter, both in rodents [15–17] and in humans [18, 19]. In 2012, Chen et al. [20] reported that maternal deprivation in rats was associated with corticotrophin-releasing hormone (CRH) promoter hypomethylation, enhancing the CRH transcriptional responses to stress. However, all of these studies have focused on early childhood life events, and for rodents, mostly maternal deprivation. Although prenatal maternal stress can lead to similar behavioral and neuroendocrinology alterations, no one has reported whether prenatal maternal stress can alter the methylation levels of related genes in offspring.
In this study, we used the bisulfite sequencing method to investigate whether maternal restraint stress can alter the methylation level of the CRH gene promoter in the hypothalamus of rat offspring. A quantitative real-time polymerase chain reaction (RT-qPCR) method was used to investigate the expression of CRH mRNA. This work provides insights into the mechanism of how maternal stress alters the activity of the HPA axis in adolescent offspring.
Materials and Methods
Animals
Adult female Sprague–Dawley rats (250 g, n = 8) were used for this study. All of the study animals were maintained at a constant temperature (23 ± 2 °C) and humidity (50–55 %), with a 12-h light/dark cycle and access to food and water. Manipulation of the animals was carried out following the National Institutes of Health guidelines for the Care and Use of Laboratory Animals and the animal experimentation regulations of Fudan University.
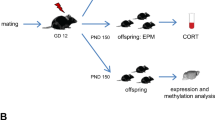
The female rats were mated overnight, and the sight of a vaginal plug the following morning was designated as the first gestational day (GD). The pregnant rats were individually housed and randomly assigned to a restraint stress (PRS) group (n = 4) or a control group (n = 4). In the PRS group, each dam underwent twice-daily restraint stress sessions in a transparent cylinder (7.0 cm diameter, 19.0 cm length) under bright light for 30 min from GD8 to GD21. The dams were allowed to raise their own litters, until postnatal day (PN) 23, when same sex litters were housed, three or four in a cage, until PN30.
At PN30, 24 offspring rats (12 male and 12 female) were randomly chosen from each group. Between PN32 and PN35, the offspring were subjected to behavioral tasks. At PN38, the offspring were killed by rapid decapitation either under basal conditions, or after undergoing restraint stress for 30 or 60 min. The restraint consisted of placing them into a plastic cylinder (5.0 cm diameter, 14.0 cm length). All experimental procedures were carried out between 9 am and 11 am. Trunk blood was collected in Eppendorf tubes and the serum was separated by centrifugation and stored at −80 °C for corticosterone determination. The brains were rapidly removed and also stored at −80 °C until microdissection.
Microdissection of the Rat Brain
For microdissection of the hypothalamus, two coronal cuts were made into the rat brain using razor blades; one through the optic chiasm (approx. Bregma 0.02), the second through the mammillary bodies (approx. Bregma 3.08). Under an anatomical microscope (Leica M205FA, Germany), a tissue block of 2 × 2 mm (width × height) was obtained by cutting just dorsal of the third ventricle and 1 mm to the left and right of the midline from the resulting slice.
Quantitative Real-Time PCR for CRH mRNA
The hypothalamus tissues were homogenized in 1 mL of TRIzol reagent (Invitrogen, USA), and the total RNA was extracted. The cDNA was then prepared using a PrimeScriptTM RT reagent kit (TAKARA, Japan). For the detection of the CRH primary transcript levels, we used the forward primer 5′-CAACCTCAGCCGATTCTGAT-3′ and the reversed primer 5′-AACACGCGGAAAAAGTTAGC-3′. For the GAPDH mRNA, the forward primer was 5′-CCCCAATGTATCCGTTGTG-3′, and the reversed primer was 5′-CTCAGTGTAGCCCAGGATGC-3′. Any relative expression changes of the target genes were calculated using the 2−ΔΔCt method, with GAPDH as the reference gene. Quantitative real-time PCR was performed using a Rotor-Gene 3000 real-time DNA analysis system (Corbett Research, Australia). Each 25 µL real-time PCR reaction, containing 2 µl cDNA, was performed using SYBR Premix ExTaqTM reagent kits (TAKARA, Japan). The PCR parameters were: 95 °C for 30 s, 40 cycles of 95 °C for 5 s, 55 °C for 30 s and 72 °C for 30 s.
Corticosterone Assays
The serum corticosterone level was determined by the enzyme-linked immunosorbent assay (ELISA), according to the manufacturer’s instructions (ENZO, USA).
Genomic DNA Extraction and Bisulfite Sequencing
The genomic DNA from the microdissection tissues of the hypothalamus was extracted using the TIANamp Genomic DNA Kit (Tiangen, China). To convert unmethylated cytosine into uracil, we used the Qiagen EpiTect Bisulfite Kit (Qiagen, USA) to treat the genomic DNA samples, following the manufacturer’s instructions. The converted DNA was purified, and then used for the amplified polymerase chain reaction (PCR). Cycling conditions were 95 °C for 5 min, followed by 50 cycles of 94 °C for 1 min, 50 °C for 1 min and 72 °C for 1 min, with a final extension of 10 min at 72 °C. The PCR reaction used HotStarTaq DNA Polymerase (Qiagen) and the following primers: ADS582, forward primer was 5′-AAGAATTTTTGTTAATGGATAAGT-3′, and reverse primer was 5′-AAATCTCACATCCAATTATATCAA-3′; and ADS583, forward primer was 5′-TTGGATGTGAGATTTAGTGTTGAA-3′, and reverse primer was 5′-ACCTTTCCCCTTTCTCTTCAAT-3′. The two forward primers were used as sequencing primers for each segment.
Open Field and Elevated Plus Maze
To assess the prenatal stress on anxiety behaviors, the offspring were evaluated using open field and elevated plus maze tests at PN30. The Sound Attenuating Cubicle Package for the Rat Open Field (Med Associates Inc., USA) was used in the open field test. The open field (43.2 cm × 43.2 cm, height 30.5 cm) was situated in a brightly lit sound-attenuating cubicle (73.7 cm × 59.7 cm, height 59.7 cm) in a dimly illuminated room. Each rat was placed by itself in a corner of the open field and allowed to explore for 5 min. The rat’s behavior was recorded and analyzed using Activity Monitor software (Med Associates Inc.). For analysis, the open field was divided into a 16-square virtual grid floor (4 × 4 square, 10.8 cm/side). The counts of crossing the central squares were measured.
The near-infrared Backlit Elevated Plus Maze Package for Rats (Med Associates Inc.) was used in the elevated plus maze test. The apparatus consisted of two opposite open arms (50.8 cm × 10.2 cm) and two opposite closed arms (50.8 cm × 10.2 cm, with a 40.6 cm high wall). The apparatus was 72.4 cm above the floor and exposed to dim illumination. The task was initiated by placing the test animal in the center of the maze facing an open arm. Activity was monitored by a mounted overhead camera for 5 min, and the rat’s behavior was analyzed using a computer-based system (Med Associates Inc.). The exploration into each arm, and the time spent in the open arms, closed arms, and center of the maze were evaluated.
Statistics
All data were expressed as the mean ± SEM, and the statistical analysis was performed using two-way ANOVA and the Tukey post hoc to adjust for the multiple tests, or Student’s t test for paired data, as indicated. A p < 0.05 was considered to indicate a significant difference.
Results
Open Field and Elevated Plus Maze
In the open field, PRS produced anxiety behaviors in both male and female offspring. The number of central zone crossings was significantly decreased in the PRS offspring when compared to the normal control, in both males (t = 5.812, p < 0.001) and females (t = 3.506, p < 0.005); furthermore, the anxiety behavior was more serious in the male PRS offspring (t = 2.27, p < 0.05) (Fig. 1a). The results of the elevated plus maze test show that the number of explorations in the open arms was significantly decreased in the PRS group, in both male (t = 5.728, p < 0.001) and female (t = 6.053, p < 0.001) offspring, and more serious in male PRS offspring (t = 2.224, p < 0.05) (Fig. 1b).

Behavioral test at PN32 and PN35. PRS offspring n = 24, normal control offspring n = 24, half male and half female in each group. a Open field test result is displayed by counts of central area entries. b Elevated plus maze test result is displayed by counts of exploration in open-arm. ***p < 0.001; **p < 0.005; *p < 0.05. (Student’s t test)
Circulation Corticosterone Level
Before the restraint stress, both the male and female offspring from the PRS group exhibited a higher serum corticosterone level than the control group. After the 30 min restraint stress, the corticosterone levels increased in both experimental groups, with the PRS group showing significantly higher levels when compared to the control group. After 60 min from the baseline, the serum corticosterone levels showed a tendency to decrease in both groups, although the PRS group levels were still significantly higher than in the control group (male: F2,66 = 89.614, p < 0.001; female: F2,66 = 169.878, p < 0.001) (Fig. 2).

The serum corticosterone level in response to acute (30 min) restraint stress. Rats were killed by decapitation in basal conditions or following 30 and 60 min of restraint stress. a Male offspring, n = 24. b Female offspring, n = 24. ***p < 0.001 compared to basal, ### p < 0.001 compared to control group, by two-way ANOVA
CRH mRNA Expression in the Hypothalamus
Alterations in CRH mRNA expression before and after 30 min of restraint stress were detected using quantitative real-time PCR. In the baseline situation (before restraint stress) CRH mRNA expression was decreased in the PRS group, when compared to those in the normal control group. After 30 min of restraint stress, the PRS group showed a significant increase in CRH mRNA expression. In the 60 min from baseline rats, the PRS group still exhibited a higher expression level (male: F2,66 = 222.738, p < 0.001; female: F2,66 = 165.049, p < 0.001) (Fig. 3).

The CRH mRNA expression in response to acute (30 min) restraint stress. Rats were killed by decapitation in basal conditions or following 30 and 60 min of restraint stress. The relative expression changes of target genes were calculated using the 2−ΔΔCt method with GAPDH as the reference gene. a Male offspring, n = 24. b Female offspring, n = 24. ***p < 0.001 compared to basal, ### p < 0.001 compared to control group, by two-way ANOVA
CRH Promoter Methylation
In order to determine whether the PRS-induced increased CRH mRNA expression was related to epigenetic alterations, we performed a methylation analysis of the CpG dinucleotides of the CRH promoter region (Fig. 4) in the hypothalamus tissue from the PRS and control groups. Prenatal restraint stress resulted in a lower percentage of DNA methylation (male t = 2.449, p < 0.05; female t = 2.966, p < 0.05) (Fig. 5), specifically in 3 CpG sites at −148 to −96 bp of the CRH promoter, compared to the normal control rats (Fig. 5).

Sequence of the corticotrophin-releasing hormone promoter −254 to −55 bp showing the CpG dinucleotides present in this region. The numbers after each CpG identify the dinucleotides subjected to methylation analysis

Methylation analysis of the CpG dinucleotide island within the rat CRH promoter region in the hypothalamus of PRS and control offspring. a The methylation rate of all CpG islands, PRS group: male n = 9, female n = 9; control group: male n = 10, female n = 8. b Significantly lower methylation of CpG 4, CpG 5 and CpG 6 (−148 to −96 bp) in the CRH promoter was observed in male offspring. c Significantly lower methylation of CpG 5 and CpG 6 was observed in female offspring. *p < 0.05. (Student’s t test)
Discussion
Our findings confirmed that adolescent rat offspring suffering PRS stress exhibit increased anxiety-like behavior and hyper-activity of the HPA axis in response to acute restraint stress. We also observed that the hyper-responsiveness of the HPA axis in PRS rats might be associated with the demethylation of CpG dinucleotides in the CRH promoter.
In this study, we found that PRS can lead to anxiety-like behavior during the adolescent period, which is in partial agreement with many previous reports [14, 21–26]. We observed anxiety-like behavior in both male and female adolescent offspring, although it was more serious in the male offspring. Some previous research studies have reported sex differences in the increased anxiety-like behavior; recently a few researchers have reported anxiety-like behavior only in the male offspring [21, 25, 26]. The reasons for these differences might be because of the different maternal stress paradigms used, and different offspring behavioral test times (early adolescent vs. adult), because estrogen could relieve the anxiety-like behavior [27–29].
Our data demonstrate that, in both male and female rat offspring, the hypothalamus CRH mRNA and serum corticosterone response to acute restraint stress were significantly higher in the PRS group when compared to the controls. This indicates that PRS was effective in inducing the hyper-responsiveness of the HPA axis, as shown by previous studies, which have demonstrated that early life experiences, including maternal stress, are linked to anxiety and depressed behavior in adolescence and adulthood, in parallel with hyperactivity of the HPA axis [9–13].
In our study, we observed that PRS is associated with reduced methylation of the CpG dinucleotides in the CRH promoter. In 2010, Elliott et al. [30] reported that in adult male mice subjected to chronic social stress, there was increased CRH expression associated with long-term demethylation of the CRH gene promoter. Mueller and Bale [31] have also found increased CRH gene expression in the amygdala in conjunction with the demethylation of the CRH promoter. Both of these studies reported that epigenetic alteration was observed in a cyclic AMP-response element (CRE) located site, between −228 and −221 bp of the CRH gene promoter, and this specific site is crucial for the regulation of CRH transcription.
It was demonstrated that the methylation of promoter CREs could decrease CREB binding and gene transcription [32, 33]. In 2012, Chen et al. [20] reported that in both male and female rats, maternal deprivation was associated with the reduced methylation of specific CpG dinucleotides of the CRH promoter, also at this CREB binding area (−232 and −226 bp). These data suggest that demethylation of the CRE in the CRH promoter is part of the mechanism mediating the increased CRH transcriptional responses to stress. However, in our study, we found demethylation mainly in sites at −148 to −96 bp. In Elliott’s study [30], they also reported demethylation within this segment, but not in Chen’s results [20]. This variability in the site specificity of the different studies is likely dependent on the type of stress, animal species and strain, or timing of the stress exposure.
In summary, our data confirms that PRS used as a model of early stress could lead to hyper-responsiveness of the HPA axis, and increased anxiety-like behavior in rat offspring. Our findings also indicate that PRS related hyper-responsiveness of the HPA axis is associated with demethylation in the CRH promoter. The parallel occurrence of the demethylation of the CRH gene in the hypothalamus suggests that altered methylation of the CRH promoter is one of the mechanisms underlying the enhanced CRH transcriptional responses to stress observed in these rats. Such epigenetic modification of the CRH gene may contribute to revealing the pathology of some psychiatric diseases which are associated with adverse experiences during early-life.
References
Dunkel Schetter C, Tanner L (2012) Anxiety, depression and stress in pregnancy: implications for mothers, children, research, and practice. Curr Opin Psychiatry 25(2):141–148
Darnaudery M, Maccari S (2008) Epigenetic programming of the stress response in male and female rats by prenatal restraint stress. Brain Res Rev 57(2):571–585
Charil A, Laplante DP, Vaillancourt C et al (2010) Prenatal stress and brain development. Brain Res Rev 65(1):56–79
O’Connor TG, Bergman K, Sarkar P et al (2013) Prenatal cortisol exposure predicts infant cortisol response to acute stress. Dev Psychobiol 55(2):145–155
Green MK, Rani CS, Joshi A et al (2011) Prenatal stress induces long term stress vulnerability, compromising stress response systems in the brain and impairing extinction of conditioned fear after adult stress. Neuroscience 192:438–451
Abe H, Hidaka N, Kawagoe C et al (2007) Prenatal psychological stress causes higher emotionality, depression-like behavior, and elevated activity in the hypothalamo-pituitary-adrenal axis. Neurosci Res 59(2):145–151
Davis EP, Glynn LM, Waffarn F et al (2011) Prenatal maternal stress programs infant stress regulation. J Child Psychol Psychiatry 52(2):119–129
Van den Bergh BR, Van Calster B, Smits T et al (2008) Antenatal maternal anxiety is related to HPA-axis dysregulation and self-reported depressive symptoms in adolescence: a prospective study on the fetal origins of depressed mood. Neuropsychopharmacology 33(3):536–545
Pawluski JL, Brain UM, Underhill CM et al (2012) Prenatal SSRI exposure alters neonatal corticosteroid binding globulin, infant cortisol levels, and emerging HPA function. Psychoneuroendocrinology 37(7):1019–1028
Pawluski JL, Rayen I, Niessen NA et al (2012) Developmental fluoxetine exposure differentially alters central and peripheral measures of the HPA system in adolescent male and female offspring. Neuroscience 220:131–141
Pawluski JL (2012) Perinatal selective serotonin reuptake inhibitor exposure: impact on brain development and neural plasticity. Neuroendocrinology 95(1):39–46
Ansorge MS, Zhou M, Lira A et al (2004) Early-life blockade of the 5-HT transporter alters emotional behavior in adult mice. Science 306(5697):879–881
Belay H, Burton CL, Lovic V et al (2011) Early adversity and serotonin transporter genotype interact with hippocampal glucocorticoid receptor mRNA expression, corticosterone, and behavior in adult male rats. Behav Neurosci 125(2):150–160
Maccari S, Morley-Fletcher S (2007) Effects of prenatal restraint stress on the hypothalamus-pituitary-adrenal axis and related behavioural and neurobiological alterations. Psychoneuroendocrinology 32(Suppl 1):S10–S15
Weaver IC, Cervoni N, Champagne FA et al (2004) Epigenetic programming by maternal behavior. Nat Neurosci 7(8):847–854
Weaver IC, D’Alessio AC, Brown SE et al (2007) The transcription factor nerve growth factor-inducible protein a mediates epigenetic programming: altering epigenetic marks by immediate-early genes. J Neurosci 27(7):1756–1768
Kosten TA, Huang W, Nielsen DA (2013) Sex and litter effects on anxiety and DNA methylation levels of stress and neurotrophin genes in adolescent rats. Dev Psychobiol
McGowan PO, Sasaki A, D’Alessio AC et al (2009) Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat Neurosci 12(3):342–348
Hompes T, Izzi B, Gellens E et al (2013) Investigating the influence of maternal cortisol and emotional state during pregnancy on the DNA methylation status of the glucocorticoid receptor gene (NR3C1) promoter region in cord blood. J Psychiatr Res 47(7):880–891
Chen J, Evans AN, Liu Y et al (2012) Maternal deprivation in rats is associated with corticotrophin-releasing hormone (CRH) Promoter hypomethylation and enhances CRH transcriptional responses to stress in adulthood. J Neuroendocrinol 24(7):1055–1064
Zuena AR, Mairesse J, Casolini P et al (2008) Prenatal restraint stress generates two distinct behavioral and neurochemical profiles in male and female rats. PLoS ONE 3(5):e2170
Weinstock M (2008) The long-term behavioural consequences of prenatal stress. Neurosci Biobehav Rev 32(6):1073–1086
Maccari S, Darnaudery M, Morley-Fletcher S et al (2003) Prenatal stress and long-term consequences: implications of glucocorticoid hormones. Neurosci Biobehav Rev 27(1–2):119–127
Zagron G, Weinstock M (2006) Maternal adrenal hormone secretion mediates behavioural alterations induced by prenatal stress in male and female rats. Behav Brain Res 175(2):323–328
Morley-Fletcher S, Mairesse J, Soumier A et al (2011) Chronic agomelatine treatment corrects behavioral, cellular, and biochemical abnormalities induced by prenatal stress in rats. Psychopharmacology 217(3):301–313
Rayen I, van den Hove DL, Prickaerts J et al (2011) Fluoxetine during development reverses the effects of prenatal stress on depressive-like behavior and hippocampal neurogenesis in adolescence. PLoS ONE 6(9):e24003
Frye CA, Walf AA (2004) Estrogen and/or progesterone administered systemically or to the amygdala can have anxiety-, fear-, and pain-reducing effects in ovariectomized rats. Behav Neurosci 118(2):306–313
Estrada-Camarena E, Fernandez-Guasti A, Lopez-Rubalcava C (2003) Antidepressant-like effect of different estrogenic compounds in the forced swimming test. Neuropsychopharmacology 28(5):830–838
Walf AA, Frye CA (2005) Antianxiety and antidepressive behavior produced by physiological estradiol regimen may be modulated by hypothalamic–pituitary–adrenal axis activity. Neuropsychopharmacology 30(7):1288–1301
Elliott E, Ezra-Nevo G, Regev L et al (2010) Resilience to social stress coincides with functional DNA methylation of the Crf gene in adult mice. Nat Neurosci 13(11):1351–1353
Mueller BR, Bale TL (2008) Sex-specific programming of offspring emotionality after stress early in pregnancy. J Neurosci 28(36):9055–9065
Zhang X, Odom DT, Koo SH et al (2005) Genome-wide analysis of cAMP-response element binding protein occupancy, phosphorylation, and target gene activation in human tissues. Proc Natl Acad Sci U S A 102(12):4459–4464
Rishi V, Bhattacharya P, Chatterjee R et al (2010) CpG methylation of half-CRE sequences creates C/EBPalpha binding sites that activate some tissue-specific genes. Proc Natl Acad Sci U S A 107(47):20311–20316
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Xu, L., Sun, Y., Gao, L. et al. Prenatal Restraint Stress is Associated with Demethylation of Corticotrophin Releasing Hormone (CRH) Promoter and Enhances CRH Transcriptional Responses to Stress in Adolescent Rats. Neurochem Res 39, 1193–1198 (2014). https://doi.org/10.1007/s11064-014-1296-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-014-1296-0