
Abstract
Parkinson’s disease (PD) is a common neurodegenerative disease, and its etiology remains obscure. Increasing evidence has suggested an important role for environmental factors such as exposure to pesticides in increasing the risk of developing PD and inflammation is the early incident during the process of PD. In this study, we measure the pro-inflammatory cytokines by enzyme-linked immunosorbnent assay and RT-PCR methods; analyze the reactive oxygen species by DCFH-DA; detected nuclear factor κB (NFκB) translocation by western blot and immunofluorescence methods; and analyze the phosphorylation of mitogen-activated protein (MAP) kinase and protein level of Nurr1 by western blot. Results showed that rotenone could induce tumor neurosis factor α (TNFα) and interleukin 1β (IL-1β) release from BV-2 cells, enhance TNFα and IL-1β mRNA levels in substantia nigra lesioned by rotenone; also, rotenone could increase the phosphorylation of inhibitor of κB (IκB), extracellular regulated protein kinase , c-Jun N-terminal kinase, p38 MAP kinases and promote p65 subunit of NFκB translocation to nuclear; at the same time, rotenone could decrease the protein level of Nurr1 in nuclear. So, rotenone exerted toxicity through activating microglia, and its mechanism might be associated with NFκB signal pathway.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Parkinson’s disease (PD) is a progressive neurodegenerative disorder affecting approximately 1 % of the population at age of 55 and increases in prevalence to roughly 5 % by the age of 85. PD was first described in 1,817, and after almost 200 years exploration by researchers, the etiology and pathogenesis of PD remain not fully elucidated. Although the mechanism of the death of neurons in PD is extremely complicated and unilluminated, inflammatory process has implicated as one of the active contributors to the death of dopaminergic neurons recently [1]. Numerous activated microglias are present in the vicinity of degenerating neurons in the substantia nigra of PD patients, and these microglias could release pro-inflammatory cytokines such as tumor neurosis factor α (TNFα), interleukin 1β (IL-1β) and so on, which would lead to the neuron damage [2].
Parkinson’s disease is thought to arise from the convergence of genetic susceptibility, environmental exposure, and ageing. It is currently believed that PD is largely sporadic, meaning that the environmental factor plays a key function in the development of PD. Epidemiological studies suggest that rural living and exposure to pesticides are associated with increased risk of PD [3]. Rotenone, the active ingredient in hundreds of pesticide products widely used as a household insecticide and as a tool for eradicating nuisance fish population in lakes and reservoirs, has been identified as an inducer of PD [4]. Researchers have demonstrated that rotenone-induced animals could mimic some clinical symptoms and pathology of PD patients [5], but the toxic mechanism of rotenone still need further research, especially the relationship between inflammation and rotenone.
Inflammation occurs in the earlier period of PD process, and that how the inflammation development remains unclear. Pesticides are strongly implicated in the etiology of PD, and several lines of evidence indicate that the pesticide is a source of stimuli of microglia activation. Microglias are resident brain macrophages that respond to environmental stresses and immunological challenges. Chronically activated microglias could overproduce soluble inflammatory mediators such as TNFα, IL-1β [6], and these pro-inflammatory cytokines could damage neuron directly or indirectly. Microglias are assembled in substantia nigra where is the predominant neurodegenerative position of PD. So this suggested that substantia nigra might precede inflammation, and neuron in substantia nigra might be very susceptible to environmental exposure.
Nuclear factor κB (NFκB) is a significant transcription factor which is associated with neuroinflammation, and is regulated by MAPK and Nurr1. In this study, we found that BV-2 microglial cells could respond to rotenone exposure, expressing and releasing pro-inflammatory cytokines; at the same time, rotenone could lead to the phosphorylation of MAPK, induce NFκB translocation from cytoplasm to nuclear and also decrease the protein level of Nurr1. So, that rotenone activated microglia might be associated Nurr1-mediated NFκB signal pathway.
Materials and Methods
Cell Culture
BV-2 microglial cells were grown in DMEM (Invitrogen, Gibco, USA) medium supplemented with 10 % heat-inactivated fetal bovine serum. Cells were maintained at 37 °C in a humidified atmosphere with 5 % CO2. Media were changed every 2 or 3 days and cells were passaged once or twice per week. Cells were transfected with appropriate plasmids by the transfection reagent Lipofectamine™ 2000 (Invitrogen) according to the manufacturer’s protocol. Cells were used for investigation at 24–48 h after transfection.
Animals
Adult male C57BL/6 J mice (6 months) were housed under a standard 12 h light/dark cycle. Animals were provided with food and water. All experiments were carried out with the approval of the local animal use committee. Efforts were made to minimize animal suffering.
Intranigral Injection of Rotenone
The skull was exposed and a hole drilled above the position of the substantia nigra pars compacta (SNpc) which lies −2.9 mm anterior, −1.3 mm lateral and −4.1 mm ventral [7]. Injections of 1 μl of rotenone solutions(10−7 mol/L) were made using a graduated glass capillary tube (Drummond Scientific Company, Broomall, PA, USA) over 5 min (0.2 μl/min) followed by 2 min of rest, to allow diffusion of the injected material, prior to removal of the needle. Phosphate-buffered saline (pH 7.4) (PBS) was the vehicle in the control group.
Vector Construction
Nuclear factor κB was a widespread transcription factor, and could regulate the expression of many inflammation factors. Within the nervous system, heterogenic NFκB is most frequently composed of two DNA-binding subunits (e.g. p50 or p65) [8]. So we used green fluorescent protein (GFP) fusion technology to attach a fluorescent tag to the p65 subunit to monitor the NFκB translocation. cDNA fragments of p65 were obtained through RT-PCR methods from BV-2 cells. The forward primer was: 5′-CCGCTCGAGCTATGGACGATCTGTTTCCCCTC-3′, the reverse primer was: 5′-CGGAATTCACCTTAGGAGCTGATCTGAC-3′, the p65 cDNA fragments and vector pEGFP-C1 were cut by XhoI/EcoRI restriction enzymes, and then were ligated by the T4 DNA ligase. The recombinant vectors were confirmed by DNA sequencing.
Western Blot Analysis
BV-2 cells were exposed to rotenone for 24 h, then washed with PBS and lysed in NP-40 lysis buffer (1 % Nonidet P-40, 150 mM NaCl, 50 mM Tris, pH 7.4, and 1 mM EDTA with protease and phosphatase inhibitors). Mice were executed and substantia nigra was separated and washed with PBS. The nuclear protein was extracted with a kit (Applygen Technologies, Beijing, China). Protein concentrations were measured with a BCA kit (Pierce). The cell lysates were solubilized in SDS sample buffer and separated by SDS-PAGE gel, then transferred to a PVDF membrane (Millipore). The membrane was blocked with 3 % BSA (Sigma) and incubated with primary antibody, followed by horseradish peroxidase (HRP)-conjugated secondary antibody, then detected with the enhanced chemiluminescence (ECL) plus detection system (Molecular Device, Lmax). Signals from specific protein brands were quantified by Gel-Pro analyzer 3.2 software.2.4.
ELISA Method
Microglial BV-2 cells were cultured in the presence of various concentrations of rotenone for 24 h. TNFα and IL-1β contents in the medium were evaluated by a colorimetric, commercial ELISA kit (NeoBioscience Technology Company, Beijing, China). Briefly, the plate was coated with TNFα or IL-1β antibody and kept at 4 °C. Before adding sample, the plate was warm to room temperature and cultured with samples at 36 °C for 90 min. After five times wash, the well was cultured with biotin-conjugated antibody for 60 min at 36 °C, and then continued with HRP-coupled antibody. For colorimetric detection, plate was incubated for 16 min with substrate TMB. The enzymatic reaction was stopped by adding sulfuric acid (2 M). The optical density of each well was measured at 450 nm using a microplate reader (Bio-Rad, Hercules, CA, USA). All samples were analyzed in triplicate.
Immunofluorescence Staining
BV-2 Cells were transfected with pEGFP-C1 or pEGFP-C1-p65 by Lipofectamine™ 2000. 24 h post transfection, cells were fixed and stained with DAPI, then imaged with laser scanning confocal microscope fluorescence microscopy (Leica TCS SP2) to capture images, and confocal data were analyzed using LCS lite Leica confocal software.
RT-PCR Method
After injected with rotenone or vehicle for 6 h, mice were executed and substantia nigra was separated and washed with PBS, then lysed with TRIzol reagent. Total RNA was extracted according to standard methods. RNA concentration was calculated from the optical density at 260 nm and the purity was determined by the 260/280 nm absorbance ratio. Total RNA (300 ng) from each sample was reverse transcribed into cDNA and amplified by polymerase chain reaction (PCR) with a gene specific for TNFα (forward primer: 5′-ATGAGCACAGAAAGCATG-3′, reverse primer: 5′-AGACAGAAGAGCGTGGTGGC-3′), IL-1β (Forward primer: 5′-ATGGCAACTGTTCCTGAACT-3′, reverse primer: 5′-TTTGAAGCTGGATGCTCTCA-3′) and β-actin (Forward primer: 5′-ATGGATGACGATATCGCTGC-3′, reverse primer: 5′-TTCTGACCCATTCCCACCATC-3′). Amplified cDNA was separated by electrophoresis in 1 % agarose gel, stained with EB, and visualized using electrophoresis gel imaging analysis system QuantiGel software. Experiments were repeated three times.
Intracellular ROS Measurement
Intracellular reactive oxygen species (ROS) were examined using DCFH-DA as described before [9]. After treated with rotenone for 6 h, BV-2 cells were incubated with 25 μmol/L DCFH-DA for 30 min at 37 °C under 5 % CO2, washed twice with PBS and stained by DAPI for counting the total cells, then visualized using the fluorescent microscope (Olympus,IX70-142). Fluorescence images were captured through a 505–530 nm band pass filter (10×).
Statistical Analysis
The data were statistically analyzed using one-way ANOVA for independent samples. Values shown represent mean ± SD. Differences were considered significant (*) when P < 0.05.
Results
Rotenone Could Activate the Microglial Cells and Induce Pro-Inflammation Cytokines Expression
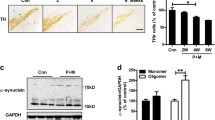
ELISA results showed that TNFα and IL-1β contents in the BV-2 medium increased significantly than that of cells exposed to solvent, and the contents of the two pro-inflammatory factors increased with rotenone in a dose-dependent manner (Fig. 1a). And mRNA level of TNFα and IL-1β from substaintia nigra injected with rotenone increase much than that from the sham group (Fig. 1b, c). Western blot results demonstrated that iNOS expression in BV-2 cells was increased after BV-2 cells exposure to rotenone for 24 h (Fig. 2a), and graph (Fig. 2b) showed the gray scale of iNOS from western blot results in BV-2 cells exposed to rotenone was increased.

Rotenone could induce TNFα and IL-1β expression. (a) Rotenone could increase TNFα and IL-1β release from BV-2 cells. BV-2 cells were exposed to rotenone for 24 h, and the TNFα and IL-1β contents in medium were measured by ELISA assay. Values represent the mean ± SD (*P < 0.05) of three independent experiments with three wells in each treatment condition. (b) Rotenone could increase TNFα and IL-1β mRNA level. After injected with rotenone or vehicle, substantia nigra was measured with RT-PCR methods, and (c) showed the gray scale of mRNA level

Rotenone could induce iNOS expression. After treated with rotenone (100 nM) for 24 h, BV-2 cells were lysis and analyzed by western blot. (a) The upper strap showed the protein level of iNOS, and the lower was β-actin expression; (b) the column showed the gray scale for iNOS
Rotenone Could Stimulate Intracellular ROS Generation
In order to gain more insight into the mechanism of how rotenone induces the expression of TNFα and IL-1β, we further analyze the ROS level. When exposed to rotenone for 6 h, BV-2 cells were measured for intracellular ROS. Results showed that the fluorescent intensity of rotenone-treated group was increased than that in control cells (Fig. 3), this result suggests that rotenone could promote ROS production in BV-2 cells.

Rotenone could promote intracellular ROS production. BV-2 cells were cultured with different content rotenone for 6 h, intracellular ROS was measured with DCFH-DA. The fluorescence demonstrated the intracellular ROS. Results showed that fluorescence in BV-2 cells increased after exposure to rotenone
Rotenone Could Lead NFκB Translocation to Nuclear
In most cases, IκB degradation is the hallmark of NFκB activation, before degradation, IκB should be phosphorylated. So, the phosphor-IκB could reflect the activation of NFκB pathway. In this study, the phospho-specific IκB in cytoplasm of substantia nigra lesioned by rotenone was increased significantly (Fig. 4a, b). When measured the expression of endogenous p65 in substantia nigra by western blot methods, we found that endogenous nuclear p65 in substantia nigra lesioned by rotenone increased much than that of control group (Fig. 4c, d); Immunofluorescence results demonstrated that the recombinant GFP-p65 expressed in BV-2 cells was located in cytoplasm under normal condition, but when exposed to rotenone, GFP-p65 was translocated from cytoplasm into nuclear (Fig. 4e). These results suggest that rotenone could induce NFκB tansactivity.

Rotenone facilitated p65 subunit of NFκB to translocate into nuclear. After injected with rotenone for 24 h, substantia nigra was lysis and analyzed by western blot. a Rotenone could increase the phosphorylation of IκB. The upper showed the increased express of phosphorylated IκB, and its protein level was shown in graph b; c p65 in nuclear increased after rotenone injection; d Column showed the gray scale of p65 in nuclear of substantia nigra. e BV-2 cells were transfected with plasmids pEGFP-C1-p65 by Lipofectame 2000™, and stimulated with rotenone for 24 h, then imaged by confocal microscope
Rotenone Could Activate MAPK Signal Pathway
Nuclear factor κB signal pathway is regulated by many modulators, and the classical pathway is MAPK mediated signal pathway. MAPK is important for neuron survival, and MAPK is activated through the phosphorylation of specific tyrosine and threonine residues. Western blot results showed that the phosphoyrlated p38, JNK and ERK were increased after rotenone exposure (Fig. 5a, b), these results showed that rotenone could activate MAPK pathway.

Rotenone could increase the phosphorylation of MAPK. After treated with rotenone for 24 h, BV-2 cells were lysis and analyzed by western blot. a Results showed that rotenone could increase the phosphorylation level of ERK, JNK and p38, b the column was the gray scale for each kinase
Rotenone Could Decrease the Protein Level of Nurr1
Nurr1, an orphan nuclear receptor, plays an essential role not only in the generation and maintenance of dopaminergic neurons, but also in protecting dopaminergic neurons from inflammation-induced death [2]. In this study, after exposure to rotenone for 24 h, Nurr1 protein level decreased in BV-2 in a dose-dependent manner (Fig. 6a), and after injected with rotenone for 24 h, nuclear Nurr1 protein level in the substantia nigra also decreased significantly (Fig. 6b, c).

Rotenone decreased Nurr1 expression. a BV-2 cells were treated with rotenone for 24 h, its lysis were analyzed by western blot. Rotenone could decrease Nurr1 expression in a dose-dependent manner. b After injected with rotenone for 24 h, substantia nigra was lysis and analyzed by western blot, the upper showed the decreased express of Nurr1 in nuclear. c Graph showed that the gray scale of nuclear Nurr1 in substantia nigra
Discussion
Parkinson’s disease is a common neurodegenerative disease with movement disorder. As the vast majority of PD cases are sporadic, environmental factors are likely to influence the onset of most cases of sporadic PD. Although the etiology of PD remains poorly understood, the disease is generally associated with an inflammatory component that is manifested in part by the presence of activated microglia and elevated serum or cerebrospinal fluid levels of pro-inflammatory factors [1]. Reports demonstrated that neurotoxin rotenone was associated with neurodegenerative disease and could imitate some features of PD, so, research of the mechanism of rotenone could help to understand the molecular mechanism PD.
Response of central nervous system towards a pathological event is characterized activation of microglia, which results in neuron death. Therefore, understanding mechanisms that regulate microglia activation is an important area of investigation that may enhance the possibility of finding a primary or an adjunct therapeutic approach against incurable neurodegenerative disorders. Under normal condition, the brain is vulnerable to oxidative stress. Report demonstrated that complex I inhibition by rotenone may result in opening of the mitochondrial permeability transition pore (PTP) [10], and the mitochondrial PTP opening induces a specific conformational change of complex I that dramatically increases ROS production [11, 12]. In this study, rotenone could stimulate intracellular ROS generation in BV-2 cells, this result might be associated with the inhibition of rotenone to mitochondrial complex I.
Reactive oxygen species could elicit a variety of pathological changes, and elevated ROS could activate MAPK and inflammatory transcription factors [13, 14]. MAPK signal transduction pathways are ubiquitous, and play important roles in the process of inflammation. Mammals express multiple MAPK pathways. The majority of these are, along with the NFκB pathway, recruited by stress and inflammatory stimuli. Extracellular signal-regulate kinase (ERK), c-Jun NH2-terminal kinases and p38MAPK, belonging to MAPK family, are activated by phosphorylation specific Ser/Thr residues, and are capable of phosphorylating both other protein kinase, transcription factors, highlighting the importance of these MAPKs to the inflammation and stress response. In this study, rotenone could induce phosphorylation of ERK, JNK, p38, and the phosphorylated MAPKs could regulate NFκB mediated signal pathways.
Nuclear factor κB is an important transcription factor and could regulate many genes expression. When binding to the specific DNA sequence, NFκB then modulates the target genes expression, its target genes include TNFα, IL-1β, iNOS and so on. In most cells, NFκB complexes are inactive, residing predominantly in the cytoplasm in a complex with inhibitory IκB proteins. When signaling pathways are activated, the IκB protein is phosphorylated and NFκB dimmers enter the nucleus to modulate target gene expression. In almost all cases, the common step in this process is mediated by the IκB kinase (IKK) complex, which phosphorylates IκB and targets it for proteasomal degradation [15]. Once activation by different stimulus and phosphorylation of IκB exposes nuclear localization signals on the p50/p65 complex resulting in nuclear translocation then flowed by gene expression and cytokine release [16]. The phosphorylation of IκB was regulated by MAPK activation [17, 18]. So, in this study, rotenone induce the activation of ERK, JNK and p38, this might be the cause of the elevated phosphorylation of IκB. Also, reporter suggested that constitutive NFκB does not trigger destructive inflammation unless accompanied by MAPK activation [19], so, in this study, the activated ERK, JNK and p38 could promote NFκB signal pathway. In turn, the activated IKK also regulate the MAPK pathway [20], then lead to a positive feedback cycle.
After phophorylation of IκB, p65 subunit of NFκB translocated into nuclear and promote genes expression. Although entering into nuclear, NFκB transactivity was also influence by another transcription factor Nurr1. Previous reports demonstrated that Nurr1 could promote the differention of dopaminergic neuron and has protective effect in nervous system [21]. Recently, researchers showed that Nurr1 is also expressed in nonneuronal cells and could impair pro-inflammatory cytokines expression through promoting the clearance of NFκB-p65 and transcriptional repression [2]. In this study, rotenone could decrease Nurr1 expression both in nuclear and cytoplasm, this result suggest that rotenone could impair the anti-inflammation effect of Nurr1 and lead to the enhanced NFκB transactivity.
Inflammation regulations often act as feedback loop. In this study, rotenone, as a mitochondria complex I, could stimulate intracellular ROS generation. Increased ROS then promotes the MAPK-NFκB signal activation, iNOS and TNFα expression, and the elevated TNFα can bind to its specific receptor, promote MAPK-NFκB signal transduction and ROS production. Then a positive feedback was induced and promoted the toxicity of rotenone.
In conclusion, rotenone could lead to the increase of ROS in microglial BV-2 cells, which then activate ERK and p38; Phosphorylation of the MAPK promotes transcription factor NFκB to nuclear. Also, decreased Nurr1 contributes NFκB to bind to target genes and then express associated protein.
Change history
16 May 2024
A Correction to this paper has been published: https://doi.org/10.1007/s11064-024-04149-1
References
Block ML, Zecca L, Hong JS (2007) Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat Rev Neurosci 8:57–69. doi:10.1038/nrn2038
Saijo K, Winner B, Carson CT, Collier JG, Boyer L, Rosenfeld MG, Gage FH, Glass CK (2009) A Nurr1/CoREST pathway in microglia and astrocytes protects dopaminergic neurons from inflammation-induced death. Cell 137:47–59. doi:10.1016/j.cell.2009.01.038
Mayeux R (2003) Epidemiology of neurodegeneration. Annu Rev Neurosci 26:81–104. doi:10.1146/annurev.neuro.26.043002.094919
Betarbet R, Sherer TB, MacKenzie G, Garcia-Osuna M, Panov AV, Greenamyre JT (2000) Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat Neurosci 3:1301–1306. doi:10.1038/81834
Bashkatova V, Alam M, Vanin A, Schmidt WJ (2004) Chronic administration of rotenone increases levels of nitric oxide and lipid peroxidation products in rat brain. Exp Neurol 186:235–241. doi:10.1016/j.expneurol.2003.12.005
Nagatsu T, Sawada M (2005) Inflammatory process in Parkinson’s disease: role for cytokines. Curr Pharm Des 11:999–1016. doi:10.2174/1381612053381620
Couch Y, Alvarez-Erviti L, Sibson NR, Wood MJ, Anthony DC (2011) The acute inflammatory response to intranigral alpha-synuclein differs significantly from intranigral lipopolysaccharide and is exacerbated by peripheral inflammation. J Neuroinflammation 8:166–179. doi:10.1186/1742-2094-8-166
Tergaonkar V (2006) NFkappaB pathway: a good signaling paradigm and therapeutic target. Int J Biochem Cell Biol 38:1647–1653. doi:10.1016/j.biocel.2006.03.023
Song JX, Shaw PC, Sze CW, Tong Y, Yao XS, Ng TB, Zhang YB (2010) Chrysotoxine a novel bibenzyl compound inhibits 6-hydroxydopamine induced apoptosis in SH-SY5Y cells via mitochondria protection and NF-kappaB modulation. Neurochem Int 57:676–689. doi:10.1016/j.neuint.2010.08.007
Li B, Chauvin C, DePaulis D, DeOliveira F, Gharib A, Vial G, Lablanche S, Leverve X, Bernardi P, Ovize M, Fontaine E (2012) Inhibition of complex I regulates the mitochondrial permeability transition through a phosphate-sensitive inhibitory site masked by cyclophilin D. Biochim Biophys Acta 1817:1628–1634. doi:10.1016/j.bbabio.2012.05.011
Zorov DB, Filburn CR, Klotz LO, Zweier JL, Sollott SJ (2000) Reactive oxygen species (ROS)-induced ROS release: a new phenomenon accompanying induction of the mitochondrial permeability transition in cardiac myocytes. J Exp Med 192:1001–1014. doi:10.1084/jem.192.7.1001
Batandier C, Leverve X, Fontaine E (2004) Opening of the mitochondrial permeability transition pore induces reactive oxygen species production at the level of the respiratory chain complex I. J Biol Chem 279:17197–17204. doi:10.1074/jbc.M310329200
Tsai HH, Lee WR, Wang PH, Cheng KT, Chen YC, Shen SC (2013) Propionibacterium acnes-induced iNOS and COX-2 protein expression via ROS-dependent NF-kappaB and AP-1 activation in macrophages. J Dermatol Sci 69:122–131. doi:10.1016/j.jdermsci.2012.10.009
Torres M, Forman HJ (2003) Redox signaling and the MAP kinase pathways. BioFactors 17:287–296. doi:10.1002/biof.5520170128
Liu S, Chen ZJ (2011) Expanding role of ubiquitination in NF-kappaB signaling. Cell Res 21:6–21. doi:10.1038/cr.2010.170
Moynagh PN (2005) The NF-kappaB pathway. J Cell Sci 118:4589–4592. doi:10.1242/jcs.02579
Singh AP et al (2012) CXCL12/CXCR4 protein signaling axis induces sonic hedgehog expression in pancreatic cancer cells via extracellular regulated kinase- and akt kinase-mediated activation of nuclear factor kappaB: implications for bidirectional tumor-stromal interactions. J Biol Chem 287:39115–39124
Zhang G, He JL, Xie XY, Yu C (2012) LPS-induced iNOS expression in N9 microglial cells is suppressed by geniposide via ERK p38 and nuclear factor-kappaB signaling pathways. Int J Mol Med 30:561–568. doi:10.1074/jbc.M112.409581
Guma M, Stepniak D, Shaked H, Spehlmann ME, Shenouda S, Cheroutre H, Vicente-Suarez I, Eckmann L, Kagnoff MF, Karin M (2011) Constitutive intestinal NF-kappaB does not trigger destructive inflammation unless accompanied by MAPK activation. J Exp Med 208:1889–1900. doi:10.1084/jem.20110242
Gantke T, Sriskantharajah S, Sadowski M, Ley SC (2012) IkappaB kinase regulation of the TPL-2/ERK MAPK pathway. Immunol Rev 246:168–182. doi:10.1111/j.1600-065X.2012.01104.x
Perlmann T, Wallen-Mackenzie A (2004) Nurr1 an orphan nuclear receptor with essential functions in developing dopamine cells. Cell Tissue Res 318:45–52. doi:10.1007/s00441-004-0974-7
Acknowledgments
This work was supported by National Natural Science Foundation of China Grants (No. 81274122, 81102831, 81073078), Special Purpose for New Drug Development (2012ZX09301002-004), Studies on Structure and function of Bioactive Substances from Natural Medicines (IRT1007), National Natural Science Foundation of Beijing Grants (7131013), Research Fund for the Doctoral Program of Higher Education of China (20121106130001).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yuan, Yh., Sun, Jd., Wu, Mm. et al. Rotenone Could Activate Microglia Through NFκB Associated Pathway. Neurochem Res 38, 1553–1560 (2013). https://doi.org/10.1007/s11064-013-1055-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-013-1055-7