
Abstract
Spatiotemporal changes of nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3) in the spinal cords of chick embryonic stage day 7 (E7) and day 14 (E14) were examined by using immunohistochemistry and Western blot. Intensive NGF immunoreaction (IR) was detected in the white matter of the spinal cords, while BDNF-IR in perikaryon and neurite, and NT-3-IR in the nucleus and cytoplasm were seen in the neurons of the ventral horn in the gray matter. Comparatively, the expressions for three growth factors have expanded largely into the dorsal horn at E14, and the level of proteins for these growth factors increased significantly in the spinal cords from E7 to E14. Morphological observation showed that the lumbar spinal cords of E7 appeared rectangular, whereas it gave a butterfly shape in the gray matter consisting of the typical ventral horn, dorsal horn and intermediate zone at E14. The present findings indicated that the spatiotemporal changes of NGF, BDNF and NT-3 could be associated to the morphological changes of developing spinal cords, suggesting the possible roles of three growth factors in the development of spinal cords.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3) are three crucial members of the neurotrophic factors (NTF). NGF has the property in promoting both the survival of sympathetic, sensory and neural crest-derived neurons [1, 2], but the neurite outgrowth from cultured dorsal root ganglion (DRG) [3, 4]. BDNF can exert crucial effects in promoting survival for several neuronal types, like sensory ganglion [5], retinal ganglion [6], basal ganglion (cholinergic neurons) [7], midbrain dopaminergic neurons [8], neural crest-derived and basal lamina-derived neurons [9–11], serine neurons [12], γ-amino butyric acid and corticospinal neurons [5]. NT-3 plays an important role in maintaining the primary sensory neurons [13], sympathetic neurons [14], motor neurons [15], hippocampus and cerebral cortical neurons [16].
While the benefit effects of NTF were determined in vitro, their role in vivo is largely unknown, especially in developing spinal cord. Up to now, there exists no report to show the expressional changes of BDNF and NT-3 in the spinal cord of embryonic chicks. It is also not clear whether there is any relationship between these growth factors and the development of the spinal cord in vivo. Previously, by using the techniques of immunoblotting and simple protein band fishing by cell (SPBFC) [17], we have found that the extract of chick spinal cords plays a crucial role in promoting the neuronal survival and the neurite outgrowth from developing DRG. The present study, therefore, was undertaken to investigate the spatiotemporal changes of NGF, BDNF and NT-3 in the spinal cord of the chick embryos, so as to find some insights to examine their possible roles in the developing spinal cord.
Materials and Methods
Characterization of Antibodies
The specificities of the three antibodies against NGF, BDNF and NT-3 (received as gifts from Professor Xin-Fu Zhou, the Department of Human Physiology and Centre for Neuroscience, Flinders University of South Australia) were confirmed by Western Blot analysis, using spinal cord homogenates of chicken in our lab. The polyclonal antisera for respective NGF, BDNF, and NT-3 were specific and no cross-reaction with other NTF was detected. As little as 10 ng of each growth factor could be visualized by using these antisera. Control of immunostaining specificity was performed by omitting the primary antibody and antibodies pre-adsorbed with the appropriate NTF. These controls did not exhibit any specific immunostaining.
Tissue Preparation
For immunohistochemical stain, the lumbar spinal cords were harvested from chick embryos at Hamburger stage 30 (n = 5) and stage 40 (n = 5). They are identical to those aged 7 days (E7) and 14 days (E14), respectively. The tissues were then fixed in 4% paraformaldehyde for 6 h, and put in 20% sucrose-PBS overnight, then sectioned at 20 μm thickness in cryostat (Leica) to later use. For Western blot, lumbar spinal cords were harvested from eight chick embryos at Hamburger stage 30 and stage 40, respectively.
Morphological Observation
The spinal cords aged E7 and E14 from chick embryos were subjected to routing hemotoxylin staining. The morphology of the spinal cords at E14 were compared with that of E7 in a light microscope.
Immunohistochemical Procedure
To show the distribution of three growth factors, sections from developing spinal cords aged E7 and E14 were processed immunohistochemical stain respectively. After 3 rinsing in PBS, the sections were treated sequentially in the following processes: immersed in 3% H2O2 for 20 min under room temperature (25°C); rinsed in PBS three times for 5 min each; blocked in 3% goat serum (prepared by mixing 0.3%Triton X-100 and 0.01 mol/l PBS) for 30 min at 37°C; incubated in PBS (prepared by mixing 0.3% Triton X-100 and 3% goat serum) consisting rabbit anti-NGF, BDNF and NT-3 serum respectively at 37°C for 2 h, and at 4°C overnight. This was followed by another washing in PBS for 10 min twice, then incubated in goat anti-rabbit IgG (1:200, Vector, dispensed in 0.01 mol/l PBS) at 37°C for 90 min. After washed thoroughly in PBS for 10 min twice again, sections were incubated in the ABC complex solution (1:100, dispensed in 0.01 mol/l PBS) at 37°C for 1 h, and washed thoroughly in PBS for 10 min twice, then developed with 0.05% DAB (in 0.05 mol/l Tris–HCl and 0.03% H2O2, pH7.5) for 5 min. The immunoreaction (IR) for each growth factor was observed in the microscope (Leika). For control sections, rabbit anti-serum for NGF, BDNF and NT-3 was replaced by PBS, and the other procedures were exactly the same as above. These sections showed negative staining.
Western Blot
100 mg spinal cord tissues of chicken embryo from E7 and E14 were obtained respectively and immediately placed in a homogenizer containing 1 ml lysis buffer and 10 μl PMSF (10 mg/ml). They were thoroughly homogenized for 5 min on ice, and centrifuged at 12,000 g for 30 min at 4°C to collect the supernatant. The protein concentration of supernatant was determined by Bio-Rad DC protein assay (every 10 μl sample contain 120 μg protein). 10 μl sample solution was separated by 15% SDS Polyacrylamide Gel Electrophoresis for 75 min at 120 V and transferred from the gels to nitrocellulose membranes (Schleicher and Schuell, USA) for 6 h at 16 V. The membranes were then immersed with 5% non-fat dry milk in PBS containing 0.05% Tween-20 (PBST) overnight at 4°C, then respectively incubated by using 1:200 dilution of rabbit polyclonal anti NGF, 1:200 dilution of rabbit polyclonal anti BDNF, 1:400 dilution of rabbit polyclonal anti NT-3 and 1:400 dilution of mouse polyconal anti β-actin overnight. This was followed by incubation with 1:400 dilution of sheep anti rabbit HRP-antibody (sheep anti-mouse HRP-antibody) for 1 h at room temperature. It was then developed in ECL kit (Boshide) and exposed against X-ray film in a dark room. The analysis of β-actin was used as an internal control. The gray scale of bands in the autoradiograph was measured and analyzed by the synaptic gene tool software (LIVE Science, USA). Then the value of gray scale for each growth factor was expressed by relative intensity to the level of β-actin.
Statistical Analysis
All data were expressed as means ± standard error of mean (SEM). Statistical significance was assessed between groups, using a one-way ANOVA test with the SPSS 11.0 for windows covariance software package. The statistical significance was defined as P < 0.05.
Results
Specificity of Antisera
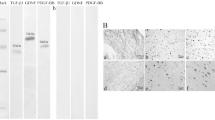
The specificity of antibodies for NGF, BDNF and NT-3 has been confirmed in our lab. The antibodies specifically recognized appropriate bands at a molecular weight of approximately 14.7, 14.2, and 13.6 kda for NGF, BDNF and NT-3, respectively.
Morphological Changes of Chick Embryonic Spinal Cords Aged E7 and E14
In transverse sections, the lumbar spinal cords at E7 appeared rectangular, and the width of both the ventral and the dorsal horn was shorter than that of the intermediate horn. Additionally, the shapes of the ventral, intermediate and dorsal horns were not typical as they were not fully formed. As a result, the central canal was comparatively wider. However, the spinal cords at E14 were about 2–3 times as big as those of E7. Meanwhile, the ventral and the dorsal horn had come into being, and the gray matter looked like a butterfly. It was easy to distinguish the ventral, intermediate and dorsal horns from one another (Fig. 1a and b).

NGF, BDNF, and NT-3 immunoreaction (IR) in the spinal cords during chicken embryonic development. NGF-IR in white matter at E7 (a, arrow showed the NGF-IR in white mater), but it was distributed in grey matter at E14, especially in dorsal horn (b, arrow showed the NGF-IR in gray mater of the dorsal horn). BDNF IR cells and neurites were seen in ventral horn at E7 (c, arrow showed the BDNF-IR products in ventral horn), but expended in dorsal horn at E14 (d, arrow). NT-3-IR is seen in neurons of the ventral horn at E7 (e, arrow), then it gradually expanded into dorsal horn at E14 (f, arrow). Magnifications: 40×
Changes in Distribution of NGF, BDNF and NT-3 in the Spinal Cords from E7 to E14 Embryo
Changes in NGF-IR Distribution
The NGF-IR was seen within the marginal region of the white matter in the lumbar spinal cord of the E7 chick embryo, especially in the ventral part of the white matter (Fig. 1a). There is no NGF-IR in the cells of gray matter. By E14, in addition to increased NGF-IR staining in the ventral white matter, its staining had also increased significantly in the gray matter, Most of the neurons in the dorsal horn exhibited NGF-IR, and the robust NGF-IR was also found within extracellular matrix (Fig. 1b).
Changes in BDNF IR Distribution
At E7, strong BDNF-IR was found in the cytoplasm of many neurons and their processes throughout the ventral horn. The positively stained fibers present crisscrossed each other in a reticular network. Many of the BDNF-IR fibbers extended from the ventral horn across the intermediate region to the base of the dorsal horn. But in most areas of the gray matter which was going to come into being the dorsal horn, the BDNF-IR was so weak that hardly any positive neurons and fibers could be seen (Fig. 1c). Nevertheless, at E14, strongly positive BDNF-IR neurons and fibers were found throughout the whole spinal dorsal horn (Fig. 1d).
Changes in NT-3 IR Distribution
At E7, only a small group of NT-3-IR positive neurons in the ventrolateral region of the ventral horn could be observed (Fig. 1e). However, when it reached E14, in addition to the NT-3-IR in this region, weakly stained NT-3-IR nucleus and cytoplasm of neurons could also be seen in the intermediate zone and the dorsal horn (Fig. 1f).
Changes in the Level of Three Growth Factors in the Spinal Cords from E7 to E14 Embryo
At E7, NGF, BDNF and NT3 protein could be detected in the spinal cords of chicken embryo by western blot analysis. Especially, they demonstrated a significant increase at E14, compared with that of E7 (Fig. 2).

Changes in the level of NGF, BDNF, and NT-3 in the spinal cords during chicken embryonic development. Western blots showed that NGF, BDNF and NT-3 increase significantly in the spinal cords from E7 (lane 1) to E14 (lane 2) within the development of embryonic chicken
Discussion
There existed prominent morphological changes in the spinal cords of chick embryos from E7 to E14. The spinal cord of E14 embryo was obviously larger than that of E7 embryo, and the outlines of the ventral, intermediate zone and dorsal horn of the spinal cord were more distinct by E14, facilitating their easy identification. Paralleling to the morphological changes is that intensive NGF-IR at E7 was seen in the white matter of the spinal cord, while BDNF-IR presents in perikaryon and neurite, and NT-3-IR in the nucleus and cytoplasm of the neurons in the ventral horn of the gray matter. Comparatively, at E14, the IR for three growth factors has expanded largely into the dorsal horn, and the level of proteins for these factors increased significantly in the spinal cords from E7 to E14. The present findings gave the crucial evidences for the distribution and changes of three growth factors in developing spinal cord. Although the mechanisms involved in the development of the spinal cord are completely unknown, the NTF look like very important in the procedure of neuronal development [5, 14, 15]. The administration of exogenous NTF has shown in the prevention of neuronal death and promotion of anatomical reorganization after various forms of spinal cord injury [18]. It was therefore interesting to focus on the role of endogenous NTF in the spinal cord development in this study.
Role of NGF in the Spinal Cord During Development
In the present study, NGF-IR was observed in the spinal white matter but not gray matter at E7 embryo. This suggested that NGF may be involved in promoting cell proliferation, migration and apoptosis, especially in the marginal region of the white matter and the neighboring ventral horn. Comparatively, NGF expressions at E14 have expanded into the gray matter, especially in the dorsal horn, suggesting that NGF was alternatively involved in the development of the sensory neurons in dorsal horn at this stage. NGF can promote the proliferation of developing DRG neurons, and induce the growth of DRG neurite under normal and lesion condition [3, 4]. Also, the differentiation of developing neurons and neurite outgrowth may be partially attributed to the induction of NGF [1, 2]. In addition, NGF induced motor neurons apoptosis by combined with p75 receptor in rat embryonic spinal cord in vitro [19], while it promoted the survival of motor neurons by combined with NGF receptor (NGFR). Motor neurons in both spinal cord and brain stem were positive for NGFR immunostaining between embryonic day 11 and postnatal day 10 by using a monoclonal antibody, 192-IgG, that specifically recognizes rat NGF receptor [20], indicating that motor neurons of the ventral horn is the target cells of NGF. Collectively, it looks like the role of NGF is heterogeneous on the development of sensory and motor neurons of the spinal cord.
Role of BDNF in the Spinal Cord During Development
BDNF and NT-3 are two important NTF involved in neuronal development. Recent studies have shown that BDNF could support the survival of spinal motor neurons in embryonic chicks [11]. The skeletal muscles and skin could express high levels of BDNF mRNA at E13 [21], and it gradually decreased in these tissues with development, while mRNA signals for BDNF and trkB could be detected in the spinal motor neurons of the ventral horn. These suggested that the motor neurons might receive the BDNF protein from the target area via a way of autocrine and/or paracrine action. In our study, strong positive BDNF-IR could be found in the cytoplasm of many neurons and their processes throughout the ventral horn. It expanded across the intermediate region, then went into the dorsal horn from E7 to E14. This suggested that BDNF is a crucial growth factor for the neuronal development of the gray matter in the spinal cord. It has been known that BDNF can rescue spinal motor neurons from axotomy-induced cell death [22]. Recent evidences also indicated BDNF may involve in the synaptic plasticity in the spinal dorsal horn [23]. Therefore, spatiotemporal changes in the expression of BDNF in the spinal cord of the chick embryo may be associated to the development both sensory neurons and motor neurons.
Role of NT-3 in the Spinal Cord During Development
NT-3 is known to have the property of being able to support the survival of cortex motor neurons [24], and enhance the activity of the CATH enzyme of cultured motor neurons [25]. Motor neurons in the spinal cord expressed trkC mRNA [15], and NT-3 could be retrogradely transported from sciatic nerves to the spinal motor neurons [26]. These suggested that NT-3 might play a crucial role in the development of motor neurons. In the present study, a significant increase for NT-3 IR in the spinal cords from E7 to E14 could be seen in the ventral horn. It suggested that NT-3 is an important factor in the development of spinal motor neurons. It has been shown that NT-3 has survival and trophic effects to support the survival of motor neurons [27–29], and the axotomy-induced reduction in firing sensitivities related to eye movements were also restored to normal values when NT-3 were administered [30].
Taken together, the present findings provided novel immunohistochemical evidences to demonstrate the spatiotemporal changes in the expression of NGF, BDNF and NT-3 in the spinal cord of the chick embryo during development. This would be available to understand the role that NGF, BDNF and NT-3 in the spinal cord with development in vivo.
References
Levi-Montalcini R, Angeletti PU (1968) Nerve growth factor. Physiol Rev 48:534–569
Levi-Montalcini R (1987) The nerve growth factor 35 years later. Science 237:1154–1162
Lindsay RM (1988) Nerve growth factors (NGF, BDNF) enhance axonal regeneration but are not required for survival of adult sensory neurons. J Neurosci 8:2394–2405
Horie H, Bando Y, Chi H, Takenaka T (1991) NGF enhances neurite regeneration from nerve-transected terminals of young adult and aged mouse dorsal root ganglia in vitro. Neurosci Lett 121:125–128
Barde YA, Edgar D, Thoenen H (1982) Purification of a new neurotrophic factor from mammalian brain. EMBO J 1:549–553
Johnson JE, Barde YA, Schwab M, Thoenen H (1986) Brain-derived neurotrophic factor supports the survival of cultured rat retinal ganglion cells. J Neurosci 6:3031–3038
Alderson RF, Alterman AL, Barde YA, Lindsay RM (1990) Brain derived neurotrophic factor increases survival and differentiated functions of rat septal cholinergic neurons in culture. Neuron 5:297–306
Hyman C, Hofer M, Barde YA et al (1991) BDNF is a neurotrophic factor for dopaminergic neurons of the substantia nigra. Nature 350:230–232
Yip HK, Johnson EM Jr (1984) Developing DRG neurons require trophic support from the central processes: evidence for a role of retrogradely transported NGF from the CNS to the peripheral. PNAS 81:6245–6249
Lindsay RM, Thonen H, Barde YA (1985) Placode and neural crest-derived sensory neurons are responsive at early developmental stages to brain derived neurotrophic factor. Dev Biol 112:319–328
Oppenheim RW, Yin QW, Prevette D et al (1992) Brain derived neurotrophic factor rescues developing avian motor neurons from cell death. Nature 360:755–757
Mamounas LA, Blue ME, Siuciak JA et al (1995) Brain derived neurotrophic factor promotes the survival and sprouting of serotonergic axons in rat brain. J Neurosci 15:7929–7939
Maisonpierre PC (1991) Human and rat brain-derived neurotrophic factor and neurotrophin-3: gene structure, distribution and chromosomal localization. Geromics 10:558–568
Fariñas I, Jones KR, Backus C et al (1994) Severe sensory and sympathetic deficits in mice lacking neurotrophin-3. Nature 369:658–661
Ernfors P, Ibáñez CF, Ebendal T et al (1990) Molecular cloning and neurotrophic activities of a protein with structure similarities to NGF: developmental and topographical expression in the brain. Proc Natl Acad Sci USA 87:5454–5458
Maisonpierre PC (1990) NT-3: a neurotrophic factor related to NGF and BDNF. Science 247:1446–1451
Wang TH, Feng ZT, Feng YY et al (2000) Preliminary exploration of the neurotrophic activity of dorsal horn extract of embryonic chicken during different developmental stage. Chinese J Neuroanat 16:379
Kao CH, Chen SH, Chio CC et al (2008) Exogenous administration of glial cell line-derived neurotrophic factor improves recovery after spinal cord injury. Resuscitation 77:395–400
Sedel F, Béchade C, Triller A (1999) Nerve growth factor (NGF) induces motor neurons apoptosis in rat embryonic spinal cord in vitro. Eur J Neurosci 11:3904–3912
Yan Q, Johnson EM Jr (1988) An immunohistochemical study of the nerve growth factor receptor in developing rats. J Neurosci 8:3481–3498
Saka Y, Yoshimura O, Tahara H et al (2007) The mRNA expression of neurotrophins in different skeletal muscles of young rats. Hiroshima J Med Sci 56:23–28
Yan Q, Elliott J, Snider WD (1992) Brain-derived neurotrophic factor rescues spinal motor neurons from axotomy-induced cell death. Nature 360:753–755
Wang TH, Wang XY, Li XL et al (2007) Effect of electro acupuncture on neurotrophin expression in cat spinal cord after partial dorsal rhizotomy. Neurochem Res 32:1415–1422
Giehl KM, Tetzlaff W (1996) BDNF and NT-3, but not NGF, prevent axotomy-induced death of rat corticospinal neurons in vivo. Eur J Neurosci 8:1167–1175
Kato AC, Lindsay RM (1994) Overlapping and additive effects of neurotrophins and CNTF on cultured human spinal cord neurons. Exp Neurol 130:196–201
Wang TH, Meng QS, Qi JG et al (2008) NT-3 expression in spared DRG and the associated spinal laminae as well as its anterograde transport in sensory neurons following removal of adjacent DRG in cats. Neurochem Res 33:1–7
Woolley A, Sheard P, Dodds K et al (1999) Alpha motor neurons are present in normal numbers but with reduced soma size in neurotrophin-3 knockout mice. Neurosci Lett 272:107–110
Gibbons A, Wreford N, Pankhurst J et al (2005) Continuous supply of the neurotrophins BDNF and NT-3 improve chick motor neuron survival in vivo. Int J Dev Neurosci 23:389–396
Uchida K, Nakajima H, Inukai T et al (2008) Adenovirus-mediated retrograde transfer of neurotrophin-3 gene enhances survival of anterior horn neurons of twy/twy mice with chronic mechanical compression of the spinal cord. J Neurosci Res 86:1789–1800
Davis-López de Carrizosa MA, Morado-Díaz CJ, Tena JJ et al (2009) Complementary actions of BDNF and neurotrophin-3 on the firing patterns and synaptic composition of motoneurons. J Neurosci 29:575–587
Author information
Authors and Affiliations
Corresponding authors
Additional information
Ying-Chun Ba and Ping Dai contributed equally to this work.
Rights and permissions
About this article
Cite this article
Ba, YC., Dai, P., Zhou, HL. et al. Spatiotemporal Changes of NGF, BDNF and NT-3 in the Developing Spinal Cords of Embryonic Chicken. Neurochem Res 35, 273–278 (2010). https://doi.org/10.1007/s11064-009-0052-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-009-0052-3