
Abstract
Methylmalonic acidemia (MMAemia) is an inherited metabolic disorder of branched amino acid and odd-chain fatty acid metabolism, involving a defect in the conversion of methylmalonyl-coenzyme A to succinyl-coenzyme A. Systemic and neurological manifestations in this disease are thought to be associated with the accumulation of methylmalonate (MMA) in tissues and biological fluids with consequent impairment of energy metabolism and oxidative stress. In the present work we studied the effect of MMA and two other inhibitors of mitochondrial respiratory chain complex II (malonate and 3-nitropropionate) on the activity of lactate dehydrogenase (LDH) in tissue homogenates from adult rats. MMA potently inhibited LDH-catalyzed conversion of lactate to pyruvate in liver and brain homogenates as well as in a purified bovine heart LDH preparation. LDH was about one order of magnitude less sensitive to inhibition by MMA when catalyzing the conversion of pyruvate to lactate. Kinetic studies on the inhibition of brain LDH indicated that MMA inhibits this enzyme competitively with lactate as a substrate (K i=3.02±0.59 mM). Malonate and 3-nitropropionate also strongly inhibited LDH-catalyzed conversion of lactate to pyruvate in brain homogenates, while no inhibition was observed by succinate or propionate, when present in concentrations of up to 25 mM. We propose that inhibition of the lactate/pyruvate conversion by MMA contributes to lactate accumulation in blood, metabolic acidemia and inhibition of gluconeogenesis observed in patients with MMAemia. Moreover, the inhibition of LDH in the central nervous system may also impair the lactate shuttle between astrocytes and neurons, compromising neuronal energy metabolism.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Methylmalonic acidemia (MMAemia) is an inherited autosomic recessive metabolic disorder affecting 1 in every 48,000 newborns [1, 2]. MMAemia is frequently caused by deficiency of methylmalonyl-coenzyme A (CoA) mutase or by abnormalities of its cofactor adenosylcobalamin [2]. Methylmalonyl-CoA mutase catalyses the conversion of methylmalonyl-CoA to succinyl-CoA, a key step in the metabolism of branched chain aminoacids isoleucine, valine, threonine and methionine, as well as odd-chain fatty acid and cholesterol degradation [2]. As a result, methylmalonyl-CoA and methylmalonate (MMA), among other metabolites, accumulate in tissues and body fluids. MMA is produced predominantly in tissues before reaching plasma and cerebrospinal fluid, where 0.2–3 mM levels have been reported during crises of metabolic decompensation [2–4]. MMAemia is characterized by generalized tissue damage with prominent effects in the brain [3–6], possibly because MMA accumulates at higher levels in this tissue, or due to the large dependence of this tissue on aerobic metabolism. In addition, our group has recently demonstrated that MMA added to Ca2+-loaded isolated brain mitochondria [7] acts as a potent inducer of membrane permeability transition, a process involved in necrotic and apoptotic cell death [8, 9].
MMAemia patients are at constant risk of metabolic decompensation with metabolic acidosis, ketonemia, hyperammonemia, hyperglycinemia, hypoglycemia, metabolic stroke, damage to basal ganglia and further neurological deterioration [2, 6, 10, 11]. The concentration of MMA does not explain the acidemia observed in MMAemia, suggesting the involvement of other processes and metabolites in this alteration. Hypoglycemia, which occurs mainly during crises [2, 12, 13], is attributed to inhibition of pyruvate carboxylase [14] and malate transport through the inner mitochondrial membrane [15], important steps in gluconeogenesis. Other enzymes involved in energy metabolism that are inhibited by MMA include succinate dehydrogenase ([16, 17]; but see [18, 19]), β-hydroxybutyrate dehydrogenase [16] and mitochondrial creatine kinase [20].
In the present study, we investigated the effect of MMA on lactate dehydrogenase (LDH) (EC 1.1.1.27) activity. LDH catalyses the interconversion of pyruvate, a product of glycolysis, and lactate. A first evidence of LDH inhibition by MMA was obtained by our group when evaluating the activity of this enzyme as a potential marker of cellular damage in brain slices exposed to MMA [7]. Based on the importance of LDH in cellular, tissue and whole body metabolism (for a recent review, see [21]), we conducted the present study to characterize the inhibitory effect of MMA and other related compounds on the activity of this enzyme.
Experimental Procedures
Reagents
Most chemicals, including l-lactic dehydrogenase from bovine heart, lactic acid, malonic acid, methylmalonic acid, β-nicotinamide adenine dinucleotide sodium salt (NAD+), β-nicotinamide adenine dinucleotide reduced disodium salt (NADH), 3-nitropropionic acid, propionic acid and pyruvic acid were obtained from Sigma-Aldrich (Sigma-Aldrich Inc., St. Louis, MO, USA). MMA, 3-nitropropionate (3-NP) and malonate (MA) were prepared by dissolving the respective acids in water and adjusting the pH to 7.2 with NaOH.
Preparation of tissue homogenates
Female Wistar rats weighing 200–250 g were used. Rats were sacrificed by decapitation. Brain, liver, heart or gastrocnemial muscles were rapidly removed (within 2 min) and put into 10 ml of ice-cold “isolation buffer” containing 225 mM mannitol, 75 mM sucrose, 1 mM K+-EGTA, 0.1 % bovine serum albumin (BSA) and 10 mM K+-HEPES pH 7.2. Cerebellum and underlying structures were removed from the brain and the remaining material (forebrain) was used. The tissues were cut into small pieces using surgical scissors and extensively washed with “isolation buffer”. The tissue was then manually homogenized in a Dounce homogenizer using both a loose-fitting and a tight-fitting pestle. The homogenate was centrifuged for 3 min at 2,000 g (forebrain), 10 min at 2,500 g (liver), 10 min at 800 g (heart) or 10 min at 700 g (muscle) in a Beckman JA 25.5 rotor (Beckman, Palo Alto, CA, USA). After the first centrifugation, the supernatant was recentrifuged for 10 min at 8,000 g (brain, liver and muscle) or 10 min at 6,000 g (heart). The supernatant from this second centrifugation, at an approximate protein concentration of 5–8 mg/ml, was used as “tissue homogenate” for the experiments. The purpose of these two centrifugations was to obtain a fraction rich in cytosolic content, but poor in heavy cellular components, nuclei and mitochondria. Control experiments showed no evidence of a considerable fraction of functioning mitochondria in the “tissue homogenates”, as verified by the lack of pyruvate-supported oxygen consumption (results not shown).
Protein determination
The protein content of brain, liver, heart and muscle homogenates was determined by the Biuret assay [22] in the presence of 0.2% deoxycholate, using BSA as a standard.
LDH activity assay
The experiments to evaluate LDH-catalyzed conversion of pyruvate to lactate were carried out at 37°C in a fluorimeter cuvette, with continuous magnetic stirring, in reaction medium containing 50 mM Tris–HCl buffer, pH 7.4, and different concentrations of pyruvate-Na+, as indicated in the figures. Experiments were conducted under these pH conditions to simulate enzymatic reactions in the cytosol. To evaluate LDH-catalyzed conversion of lactate to pyruvate the experiments were carried out in reaction medium containing different concentrations of lactate-Na+, as indicated in the figures. After 30 sec of pre-incubation of “tissue homogenates” (20–40 μg of protein) or bovine heart LDH (1.47 μg) in 2 ml of reaction medium, the reaction was started by the addition of 0.1 mM NADH or 0.8 mM NAD+, for measurement of conversion of pyruvate to lactate or lactate to pyruvate, respectively. The reaction was stopped after 3 min. Different concentrations of MMA, malonate, 3-nitropropionate, succinate and propionate were present since the beginning of the pre-incubation period. The reagent concentrations and the amount of protein added were chosen to assure linearity of the enzymatic reaction at least during the first 30–40 sec. Rates of produced or consumed NADH were estimated considering the reaction rates between 10 and 40 sec (5–15 sec, for heart and muscle) after NAD+ or NADH addition, respectively. LDH activity was followed by measuring NADH fluorescence levels recorded on a model F-4010 Hitachi spectrofluorometer (Hitachi Ltd., Tokyo, Japan) operating at excitation and emission wavelengths of 366 and 450 nm, respectively. Both slit widths used were at 2.5 nm or 5.0 nm, for reactions following NADH consumption or production, respectively.
LDH-catalyzed conversion of pyruvate to lactate was 0.97±0.12, 1.87±0.39, 7.72±1.31, and 14.48±6.10 mM NADH min−1 mg−1, for brain, liver, heart and muscle tissue homogenates, respectively, using 0.1 mM pyruvate. LDH-catalyzed conversion of lactate to pyruvate was 0.18±0.03 and 0.26±0.03 mM of NAD+ min−1 mg−1, for brain and liver tissue homogenates, respectively, using 2 mM lactate. Using a purified bovine heart LDH, enzymatic activity was 43.53±4.71 mM of NAD+ min−1 mg−1, for lactate/pyruvate conversion. It is important to note that to follow lactate/pyruvate conversion, a higher concentration of the substrate was used. In addition, some experiments were conducted in the presence of 50 mM hydroxylamine, a trapping agent for the pyruvate formed.
Statistical analysis
Data from the experiments were analyzed by one-way ANOVA followed by Tukey’s post-hoc test performed by OriginPro 7.5 SRO (OriginLab Corporation, Northampton, MA, USA). When one parameter was compared between two groups, Student’s t-test was used. Data are presented as averages±standard deviation (±SD) of at least three experiments conducted with different preparations.
Results
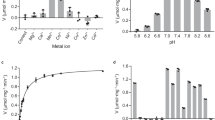
The in vitro effect of MMA, at concentrations varying from 5 to 50 mM, on LDH-catalyzed conversion of pyruvate to lactate was studied in tissue homogenates from adult rat brain (Fig. 1a, b), liver (Fig. 1c, d), heart (Fig. 1e) and muscle (Fig. 1f). Panels a and c depict representative traces of LDH-catalyzed pyruvate/lactate conversion in brain and liver homogenates, respectively. Similar patterns were obtained in heart and muscle homogenates (results not shown). MMA significantly inhibited LDH activity in a dose-dependent manner, with IC50 of 23.3±0.6 mM, 20.8±2.9 mM, 16.5±5.1 mM and 56.2±13.5 mM, for brain, liver, heart and muscle homogenates, respectively. The IC50 of MMA for pyruvate/lactate conversion in muscle homogenate was significantly higher than that observed in brain, liver and heart homogenates (P<0.05, Tukey’s test). The concentration of pyruvate used in these experiments was in the range of those in tissue and whole blood under physiological conditions [23–26]. Moreover, pyruvate concentrations above 250 μM resulted in a progressive inhibition of the reaction rate in brain homogenates (results not shown), in accordance with an early report [27]. Under our experimental conditions, no significant NADH oxidation was observed in the presence of brain homogenate but in the absence of added pyruvate, indicating that the presence endogenous NADH oxidases is not significantly affecting the measurements (results not shown).

Effect of methylmalonate (MMA) on lactate dehydrogenase (LDH)-catalyzed conversion of pyruvate to lactate in brain (a, b), liver (c, d), heart (e) and muscle (f) homogenates from adult rats. The reactions were started by the addition of 0.1 mM NADH to the reaction medium containing 0.1 mM pyruvate and tissue homogenate. Panels a and c depicted representative traces of LDH-catalyzed pyruvate/lactate conversion measured by NADH oxidation in brain and liver homogenates, respectively, in the absence (control) and presence of 25 mM MMA. Data in Panels b, d, e and f are mean±S.D. of 3–5 experiments performed in duplicate. *Significantly different from control, P<0.05. **Significantly different from control, P<0.01
Interestingly, LDH-catalyzed conversion of lactate to pyruvate in homogenates from rat brain (Fig. 2a, b) and liver (Fig. 2c, d) was inhibited by lower concentrations of MMA, with an IC50 of 4.6±0.39 mM and 5.6±1.3 mM, respectively. When lactate/pyruvate conversion in brain homogenate was determined in the presence of hydroxylamine, a trapping agent for the pyruvate formed [27], the MMA IC50 was 5.50 ± 0.86 mM. The concentration of lactate used in these experiments was in the range of physiological concentrations of lactate (venous blood: 0.6–2.2 mmol/l, arterial blood: 0.3–0.8 mmol/l) [25].

Effect of MMA on LDH-catalyzed conversion of lactate to pyruvate in brain (a, b) and liver (c, d) homogenates from adult rats. The reactions were started by the addition of 0.8 mM NAD+ to the reaction medium containing 2 mM lactate and tissue homogenate. Panels a and c depicted representative traces of LDH-catalyzed lactate/pyruvate conversion measured by NAD+ reduction in brain and liver homogenates, respectively, in the absence (control) and presence of 5 mM MMA. Data in Panels b and d are mean±S.D. of 3–5 experiments performed in duplicate. **Significantly different from control, P<0.01
Since tissue homogenates contain many enzymes in addition to LDH that could convert MMA to other metabolites [18, 19], the effect of MMA was tested on purified LDH from bovine heart (Fig. 3). This enzyme preparation was also sensitive to MMA in a dose-dependent manner, when converting lactate to pyruvate, with an IC50 of 5.2±0.57 mM.

Effect of MMA on purified bovine heart LDH-catalyzed conversion of lactate to pyruvate. The reactions were started by the addition of 0.8 mM NAD+ to the reaction medium containing 2 mM lactate and tissue homogenate. Data are mean±S.D. of 3 experiments performed in duplicate. **Significantly different from control, P<0.01
In order to better characterize the in vitro inhibitory effect of MMA on brain LDH activity, we carried out competition experiments between MMA and lactate. The K m for lactate as substrate was 1.79±0.32 mM (n=4). The Lineweaver-Burk plot indicated that MMA inhibited LDH activity by competition with lactate (Fig. 4). The K i value was 3.02±0.59 mM (n=4) for MMA, calculated using the Dixon plot [28] (result not shown).

Lineweaver-Burk competition plots between lactate and MMA on LDH activity (μM NAD+ min−1 mg−1) in rat brain homogenates. Data are representative of 4–5 experiments performed in duplicate. (◯) control (without inhibitor); (•) 1 mM; (■) 5 mM; (▴) 10 mM; (◆) 25 mM MMA
Since MMA [16, 17] or its metabolites [18, 19] inhibit succinate dehydrogenase, experiments were done to compare the effects of two other succinate dehydrogenase inhibitors: 3-nitropropionate (Fig. 5a) and malonate (Fig. 5b), on brain LDH-catalyzed conversion of lactate to pyruvate. 3-Nitropropionate and malonate significantly inhibited LDH activity in a dose-dependent manner, with an IC50 of 3.58±1.28 mM and 1.41±0.31 mM, respectively. In addition, the effects of propionate (Fig. 5c) and the physiological substrate succinate were also evaluated. Lactate/pyruvate conversion was not inhibited by succinate (results not shown) and was slightly (non-significantly) sensitive to propionate when present in concentrations up to 25 mM (Fig. 5c).

Effect of 3-nitropropionate (3-NP) (a), malonate (MA) (b) or propionate (PA) (c) on LDH-catalyzed conversion of lactate to pyruvate in rat brain homogenates. The reactions were started by the addition of 0.8 mM NAD+ to the reaction medium containing 2 mM lactate and tissue homogenate. Data are mean±S.D. of 3 experiments performed in duplicate. *Significantly different from control, P<0.05. **Significantly different from control, P<0.01
Discussion
In the present work we studied the inhibitory effect of MMA on LDH activity in homogenates obtained from adult rat brain, heart, liver and muscle. LDH-catalyzed conversion of either pyruvate to lactate or lactate to pyruvate was evaluated. Interestingly, MMA levels observed during crises of metabolic decompensation in MMAemia, i.e. up to 3 mM [2–4], extensively inhibit LDH-catalyzed conversion of lactate to pyruvate (Figs. 2–4), but have a minor inhibitory effect on the conversion of pyruvate to lactate (Fig. 1) when the experiments were conducted in the presence of pyruvate and lactate in the range of physiological concentrations. Since MMA [16, 17] or its metabolites [19] inhibit the respiratory chain enzyme succinate dehydrogenase, we also tested the effects of a competitive (malonate) and of a non-competitive and irreversible (3-nitropropionate) succinate dehydrogenase inhibitors. Both malonate and 3-nitropropionate potently inhibited lactate/pyruvate conversion (Fig. 5). However, LDH activity was not inhibited by the physiological substrate succinate. Recently, most of the inhibitory effects of MMA on respiratory chain have been attributed to intramitochondrial production of metabolites such as malonate and 2-methylcitrate [18, 19]. Under our conditions, we observed a potent inhibition of LDH-catalyzed conversion of lactate to pyruvate in a purified preparation of this enzyme (Fig. 3), in which the production of MMA metabolites is not expected. This indicates that MMA is the primary inhibitor of this reaction.
In mammalian cells, LDH isoenzymes are composed of four polypeptide chains, containing different ratios of two kinds of polypeptide: the A chain (also designated M for muscle) and the B chain (also designated H for heart) [29]. The A chain predominates in tissues subject to anaerobic conditions such as skeletal muscle (A4 isoenzyme) and the B chain predominates in aerobic tissues such as heart muscle (B4 isoenzyme). The A chain appears to mainly function in the reduction of pyruvate by NADH into lactate, whereas the B chain appears to be better adapted to catalyze the reverse reaction. In brain tissue, a predominance of B4 isoenzyme is observed in neurons, while astrocytes present both A4 and B4 isoenzymes [30], yielding a mixture of LDH isoenzymes in brain homogenates. Liver presents predominantly the A4 isoenzyme [31]. Considering that our results show that both liver (A4) and heart (B4), but not muscle (A4), have similar sensitivity to MMA-inhibited pyruvate/lactate conversion (Fig. 1), there is no apparent indication for different susceptibility to the inhibitory effect of MMA regarding the LDH isoenzyme.
Kinetics studies showed that MMA inhibits brain LDH by competition with the substrate lactate (Fig. 4). The K i value (3.02±0.59 mM) was in the range of the MMA plasma concentration observed in crises of metabolic decompensation in MMAemia (2–4). The K m for lactate under our experimental conditions (1.79±0.32 mM, brain homogenate) is in the range of physiological concentrations of lactate (venous blood: 0.6–2.2 mmol/l, arterial blood: 0.3–0.8 mmol/l) [25], indicating that LDH is not saturated in respect to lactate. These observations indicate that MMA could actually be a relevant competitive inhibitor of the lactate/pyruvate conversion under pathological situations.
In conclusion, our findings indicate that MMA is a potent inhibitor of LDH-catalyzed conversion of lactate to pyruvate. This inhibition may contribute to lactate accumulation and inhibition of gluconeogenesis observed in MMAemia [2, 13, 14, 32], since lactate is an important source of carbon atoms for glucose synthesis by gluconeogenesis. Impairment of mitochondrial function due to membrane permeability transition promoted by MMA and Ca2+ [7] would enhance glycolytic metabolism and also contribute to lactate accumulation in MMAemia. Moreover, LDH inhibition may impair the lactate shuttle between astrocytes and neurons [21, 33], compromising neuronal energy metabolism and glutamatergic neurotransmission when MMA reaches high concentrations in the CNS.
References
Coulombe JT, Shih VE, Levy HL (1981) Massachusetts metabolic disorders screening program. II. Methylmalonic aciduria. Pediatrics 67:26–31
Fenton WA, Gravel RA, Rosenblatt DS (2001) Disorders of propionate and methylmalonate metabolism. In: Scriver CR, Beaudet AL, Valle AD, Sly W (eds) The metabolic and molecular basis of inherited disease, 8th edn. McGraw-Hill, New York, pp 2165–2193
Hoffmann GF, Gibson KM, Trefz FK, Nyhan WL, Bremer HJ, Rating D (1994) Neurological manifestations of organic acid disorders. Eur J Pediatr 153:S94–S100
Hoffmann GF, Meier-Augenstein W, Stöckler S, Surtees R, Rating D, Nyhan WL (1993) Physiology and pathophysiology of organic acids in cerebrospinal fluid. J Inherit Metab Dis 16:648–669
Leonard JV (1995) The management and outcome of propionic and methylmalonic acidaemia. J Inherit Metab Dis 18:430–434
de Baulny HO, Benoist JF, Rigal O, Touati G, Rabier D, Saudubray JM (2005) Methylmalonic and propionic acidaemias: management and outcome. J Inherit Metab Dis 28:415–423
Maciel EN, Kowaltowski AJ, Schwalm FD, Rodrigues JM, Souza DO, Vercesi AE, Wajner M, Castilho RF (2004) Mitochondrial permeability transition in neuronal damage promoted by Ca2+ and respiratory chain complex II inhibition. J Neurochem 90:1025–1035
Kowaltowski AJ, Castilho RF, Vercesi AE (2001) Mitochondrial permeability transition and oxidative stress. FEBS Lett 495:12–15
Green DR, Kroemer G (2004) The pathophysiology of mitochondrial cell death. Science 305:626–629
Heidenreich R, Natowicz M, Hainline BE, Berman P, Kelley RI, Hillman RE, Berry GT (1988) Acute extrapyramidal syndrome in methylmalonic acidemia: “metabolic stroke” involving the globus pallidus. J Pediatr 113:1022–1027
Trinh BC, Melhem ER, Barker PB (2001) Multi-slice proton MR spectroscopy and diffusion-weighted imaging in methylmalonic acidemia: report of two cases and review of the literature. Am J Neuroradiol 22:831–833
Rosenberg LE (1978) Disorders of propionate, methylmalonate and cobalamin metabolism. In: Stanbury JB, Wyngaarden JB, Fredrickson DS (eds) The metabolic basis of inherited disease, 4th edn. McGraw-Hill, New York, p 411
Worthen HG, Al Ashwal A, Ozand PT, Garawi S, Rahbeeni Z, Al Odaib A, Subramanyam SB, Rashed M (1994) Comparative frequency and severity of hypoglycemia in selected organic acidemias, branched chain amino acidemia, and disorders of fructose metabolism. Brain Dev 16(Suppl):81–85
Oberholzer VG, Levin B, Burgess EA, Young WF (1967) Methylmalonic aciduria. An inborn error of metabolism leading to chronic metabolic acidosis. Arch Dis Child 42:492–504
Halperin ML, Schiller CM, Fritz IB (1971) The inhibition by methylmalonic acid of malate transport by the dicarboxylate carrier in rat liver mitochondria. A possible explanation for hypoglycemia in methylmalonic aciduria. J Clin Invest 50:2276–2282
Dutra JC, Dutra-Filho CS, Cardozo SE, Wannmacher CM, Sarkis JJ, Wajner M (1993) Inhibition of succinate dehydrogenase and beta-hydroxybutyrate dehydrogenase activities by methylmalonate in brain and liver of developing rats. J Inherit Metab Dis 16:147–153
Brusque AM, Borba Rosam R, Schuckm PF, Dalcin KB, Ribeiro CA, Silva CG, Wannmacher CM, Dutra-Filho CS, Wyse AT, Briones P, Wajner M (2002) Inhibition of the mitochondrial respiratory chain complex activities in rat cerebral cortex by methylmalonic acid. Neurochem Int 40:593–601
Okun JG, Horster F, Farkas LM, Feyh P, Hinz A, Sauer S, Hoffmann GF, Unsicker K, Mayatepek E, Kolker S (2002) Neurodegeneration in methylmalonic aciduria involves inhibition of complex II and the tricarboxylic acid cycle, and synergistically acting excitotoxicity. J Biol Chem 277:14674–14680
Kolker S, Schwab M, Horster F, Sauer S, Hinz A, Wolf NI, Mayatepek E, Hoffmann GF, Smeitink JA, Okun JG (2003) Methylmalonic acid, a biochemical hallmark of methylmalonic acidurias but no inhibitor of mitochondrial respiratory chain. J Biol Chem 278:47388–47393
Schuck PF, Rosa RB, Pettenuzzo LF, Sitta A, Wannmacher CM, Wyse AT, Wajner M (2004) Inhibition of mitochondrial creatine kinase activity from rat cerebral cortex by methylmalonic acid. Neurochem Int 45:661–667
Gladden LB (2004) Lactate metabolism: a new paradigm for the third millennium. J Physiol 558:5–30
Gornall AG, Bardawill CJ, Donid MM (1949) Determination of serum protein by means of the Biuret reaction. J Biol Chem 177:751–766
Sahlin K, Katz A, Broberg S (1990) Tricarboxylic acid cycle intermediates in human muscle during prolonged exercise. Am J Physiol 259:C834–841
Spencer MK, Katz A, Raz I (1991) Epinephrine increases tricarboxylic acid cycle intermediates in human skeletal muscle. Am J Physiol 260:E436–E439
Lehmann HP, Henry JB (2001) SI Units. In: Henry JB (ed) Clinical diagnosis and management by laboratory methods, 20th edn. W.B. Saunders Company, Philadelphia, USA, pp 1426–1441
Comar JF, Suzuki-Kemmelmeier F, Bracht A (2003) The action of oxybutynin on haemodynamics and metabolism in the perfused rat liver. Pharmacol Toxicol 93:147–152
Hakala MT, Glaid AJ, Schwert GW (1956) Lactic dehydrogenase. II. Variation of kinetic and equilibrium constants with temperature. J Biol Chem 221:191–209
Dixon M, Webb EC (eds) (1964) Enzyme kinetics, 2nd edn. Longman, London, UK
Kaplan NO, Everse J (1972) Regulatory characteristics of lactate dehydrogenases. Adv Enzyme Regul 10:323–336
Bittar PG, Charnay Y, Pellerin L, Bouras C, Magistretti PJ (1996) Selective distribution of lactate dehydrogenase isoenzymes in neurons and astrocytes of human brain. J Cereb Blood Flow Metab 16:1079–1089
Brooks GA, Dubouchaud H, Brown M, Sicurello JP, Butz CE (1999) Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. Proc Natl Acad Sci USA 96:1129–1134
Wajner M, Coelho JC (1997) Neurological dysfunction in methylmalonic acidaemia is probably related to the inhibitory effect of methylmalonate on brain energy production. J Inherit Metab Dis 20:761–768
Pellerin L, Magistretti PJ (1994) Glutamate uptake into astrocytes stimulates aerobic glycolysis: a mechanism coupling neuronal activity to glucose utilization. Proc Natl Acad Sci USA 91:10625–10629
Acknowledgments
We thank Dr. Alicia Kowaltowski for discussions and Edilene S. Santos and Elisangela J. Gomes for technical support. This study was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and National Institutes of Health (NIH) (grant number: 1 R03 HD047388–01A1). L.O.S and S.R.M. were supported by CNPq fellowships and E.N.M. by a FAPESP fellowship.
Author information
Authors and Affiliations
Corresponding author
Additional information
S. R. Mirandola and E. N. Maciel contributed equally to this work.
Rights and permissions
About this article
Cite this article
Saad, L.O., Mirandola, S.R., Maciel, E.N. et al. Lactate Dehydrogenase Activity is Inhibited by Methylmalonate in vitro . Neurochem Res 31, 541–548 (2006). https://doi.org/10.1007/s11064-006-9054-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-006-9054-6