
Abstract
MicroRNA is an important regulator of glioblastoma. This study aims at validating microRNA-221 (miR-221) as a biomarker for glioblastoma, and understanding how miR-221 regulates glioblastoma progression. Using clinical samples, miR-221 expression was analyzed by quantitative reverse-transcriptase PCR (qPCR). SHG-44 cells were treated with anti-miR-221 or U87MG-derived exosomes followed by monitoring changes in cell viability, migration and temozolomide (TMZ) resistance. Bioinformatics approach was used to identify targets of miR-221. The interaction between miR-221 and its target, DNM3 gene, was studied with dual-luciferase reporter assay, Spearman’s correlation analysis, and western blotting. To verify that RELA regulates miR-221 expression, RELA-expressing vector or shRNA was introduced into SHG-44 cells and its effect on miR-221 expression was monitored. Both tissue-level and exosomal miR-221 expression increased with glioma grades. In SHG-44 cells, downregulating miR-221 expression inhibited cell proliferation, migration, and TMZ resistance, whereas incubation with U87MG-derived exosomes exerted tumor-promoting effects. DNM3 gene is a target of miR-221. RELA induced miR-221 expression. In glioma, elevated miR-221 expression is a biomarker for glioma. DNM3 is a target of miR-221 and RELA regulates miR-221 expression. The RELA/miR-221 axis is a target for glioma diagnosis and therapy.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Human gliomas is the most common primary tumours of the central nervous system [1], which persists as one of the most aggressive and lethal cancer due to its high metastasis rate and frequently developed drug resistance [2]. Among all gliomas, glioblastoma (GBM) constitutes 55% all primary gliomas, and patients afflicted with GBM survive on average <1 year. Current therapies against GBM, such as surgery, radiotherapy and chemotherapy frequently lead to recurrence, which universally retaliates with unrelenting progression to death. Apart from the rapid growth, high invasiveness, and genetic heterogeneity of GBM, the dismal survival of GBM patients also originates from the poor understanding of the precise mechanism governing disease progression [3]. Therefore, it is crucial to elucidate how GBM progression is modulated.
MicroRNAs are a class of small non-encoding RNAs of 18–24 nucleotides, which regulate protein expression of specific mRNAs post-transcriptionally by either inhibition of gene expression or mRNA degradation [4]. They are gene regulators that play an important role in tumor growth. In GBM, aberrant miRNA expression is closely associated with enhanced tumor proliferation, invasion and metastasis and consequently can be used as an indicator of cancer malignancy [5]. Exosomes, which contain abundant miRNAs, can shuttle between cells, resulting in cross-cellular gene-regulation through exosome release and uptake. Tumour-derived (TD) exosomes contain miRNAs that modulate cancer progression. Its functions includes promoting tumor invasion and metastasis by remodeling the extracellular matrix, regulating stromal cells, and stimulating angiogenesis [6]. Recent studies also indicate that an increased drug resistance coincides with increased secretion of exosomes [7]. It is widely believed that miRNAs in exosomes alters the chemo-susceptibility by modulating cell cycle distribution and drug-induced apoptosis after binding, absorption, and internalization of exosomes.
The elevated miR-221 expression has been reported in a variety of cancer, including hepatocellular, prostate cancer colorectal cancers [8–11]. Previously, circulating miR-221 level was found to be markedly upregulated in glioma cells [12–14]. However, clinical data regarding miR-221 level and glioma malignancy is still lacking. Moreover, despite recent efforts in unraveling the mechanism of miR-221 in glioma progression [12–14], the target of miR-221 and how miR-221 is regulated in glioma progression remain unclear.
In the present study, we sought to investigate the correlation between miR-221 expression and glioma grade using clinical sample, and evaluate how miR-221 regulates cell proliferation, migration and TMZ resistance. Our data suggested that miR-221 is a pro-oncogenic factor in glioma and it exerts anti-apoptotic effect in a human glioblastoma cell lines, SHG-44. Further we proceeded to explore the underlying molecular mechanism of miR-221 in GBM progression. Bioinformatic analysis indicated that two putative binding sites for miR-221 exist in Dynamin-3 (DNM3) gene. DNM3, a member of the Dynamin family, is a GTPase that is recently revealed to correlate with malignant diseases [15]. To our knowledge, this is the first time that DNM3 is reported to correlate with miR-221 in glioma. DNM3 is the target of miR-221 and overexpression of DNM3 could reverse the miR-221’s tumour-promoting effect. Since RELA (also known as p65) is a crucial regulator of oncogenic pathways, we also investigated if miR-221 is regulated by RELA gene.
Materials and methods
Cell culture
All human glioma cell lines used in this study, including SHG-44, U87MG, HEB and U251, were acquired from American Type Culture Collection (ATCC, USA), and cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% Pen/Step at 37 °C in 5% CO2. Cells used for evaluating drug resistance were exposed to temozolomide at the concentration of 100 µM in culture medium.
Extraction of exosomes
Exosome extraction was performed when U87MG cells reached 80% confluency. Then the medium was collected, which was centrifuged at 2000×g for 20 min followed by filtration through a 0.22 µm filter to remove cell debris. Supernatant (10 mL) was mixed with 2 mL of ExosoQuick precipitation solution and incubated overnight at 4 °C, and centrifuged at 500×g for 30 min. Exosomes in the pellet were resuspended in 100 µL phosphate buffered saline (PBS) and washed twice with PBS.
Viability and apoptosis study
Cell viability and proliferation was evaluated by CCK-8 assay. Cells were firstly plated in 96-well plates and incubated with CCK-8 solution for 4 h. Absorption at 450 nm was measured with a plate reader to evaluate the cell viability. Flow cytometry was used to determine cell apoptotic rates after temozolomide treatment. At 48 h after incubation with temozolomide, DNA fragmentation was detected by flow cytometry after Propidium iodide (PI) staining. Cell apoptotic rates were expressed as ratios of the cells containing fractional DNA content relative to viable cells.
Migration assay
Both transwell matrigel invasion assay and scratch wound assay were used to evaluate the migratory ability of cells. In tranwell assay, cells (1 × 105) were seeded in the upper chamber of a 24-well transwell unit (Corning, New York, NY, USA) pre-coated with matrigel. DMEM (500 µL) was then applied to the lower chamber. Invading cells in the bottom chamber were fixed and measured by measuring absorbance at 570 nm at 24 h after incubation.
Oligonucleotides and cell transfection
The antisense oligonucleotide sequences against miR-221 (anti-miR-221) is 5′-AGCUACAUUGUCUGCUGGGUUUC-3′. The scramble oligonucleotide (anti-miR-NC) has a sequence of 5′-UCUACUCUUUCUAGGAGGUUGUGA-3′. Oligonucleotides were synthesized in solid phase and purified by high-performance liquid chromatography (GenePharma Co., Ltd., Shanghai, China). Cells plated in six-well plates using serum-free medium were transfected with 200 pmol oligonucleotides with Lipofectamine 2000 (Invitrogen, USA). At 6 h after transfection, medium was replaced again with complete medium and cultured at 37 °C in 5% CO2.
Lentivirus packaging
To induce miR-221, miR-NC, and DNM3 expression in SHG-44 cells, the lentivirus carrying plasmid cDNA encoding these sequences were packaged using the lentiviral packaging kit from Thermo Fisher Scintific according to the manufacturer’s guidelines. Briefly, at 48 h posttransfection, the resulting lentiviral supernatant in transfected SHG-44 culture was collected and further filtered through a 0.45-μm-pore-size filter and used to infect cells in the presence of 10 μg/mL of Polybrene (Millipore, Billerica, MA). The transduced cells were then selected with puromycin (2 μg/mL) to establish cell lines in which stable expression was achieved.
Quantitative reverse-transcriptase PCR
Glioma tissue or normal tissue were collected from The Second Hospital of Hebei Medical University. Written informed consent was obtained from all patients under institutional review board approved protocols. This study was approved by the ethics committee of The Second Hospital of Hebei Medical University. Total RNA from tissue or cells were extracted using Trizol reagent (Invitrogen), with genomic DNA removed with TurboDNase Kit (Amibion). Extracted RNA was quantified by measuring absorbance at 260 nm. Synthesis of cDNA was performed using PrimeScripRT reagent KIT (Takara Bio, Japan) using a thermal cycler (Eppendorf, Germany). Expression of miR-221 were determined using miScriopt primer assays and normalized using the 2−ΔΔT method, using Syber Green (Qiagen) real-time PCR relative to GAPDH expression.
Western blot analysis
Cell were collected with cell scraper, centrifuged at 14,000×g for 5 min in 4 °C and lysed using Cell Lysis Buffer (P0013, Beyotime Biotechnology, China) supplemented with PMSF (1 mM). Protein content in supernatant was measured with DC protein assay (Biorad, Hercules, CA, USA) and loaded in precast gels (20 µg/lane) and separated on SDS–PAGE. Polyvinylidene difluoride (PVDF) membranes were used in transferring. Membranes were blocked with 5% nonfat milk for 1 h at room temperature and incubated with primary antibody against DNM3 and GAPDH (Sigma, St. Louis, MO) overnight at 4 °C with gentle shaking. Blots were then washed with TBST (0.1%) and incubated with HRP-conjugated secondary antibodies (Life Technologies), followed by washing and band visualization using chemiluminescence reagents before exposure to gel imaging system.
Dual-luciferase reporter assay
Wild-type and mutant DNM3 genes were subcloned into pGL3 Basic luciferase reporter plasmid (Progema), and transfected with the aid of FuGENE 6 (Boehringer) according to manufacturer’s protocol. Renila luciferase (pRL-TK, Promega) was co-transfected in each sample as an internal control for evaluation of transfection efficiency.
Animal study
6 weeks old Balb/c athymic mice were provided by Hebei Medical University. All animal experiments were performed according to protocols approved by Animal Care Committee of the Second Hospital of Hebei Medical University. Mice were housed in a facility maintained at 20–25 °C with a 12-h light/dark cycle. Cells (1 × 106) dispersed in 100 µL medium were injected subcutaneously into each side of posterior flank of the mice (n = 6). Over a period of 20 days after tumor inoculation, tumor size were monitored every other day and the tumor volume were calculated as: volume = 0.5 × length × width2. Mice were sacrificed at the 20th day and tumor were collected. The tumor weight was photographed using a digital camera.
Statistical analysis
In all studies, one or two way ANOVA analysis followed by a Tukey’s post hoc test was performed to evaluate the significance of differences. Differences were considered statistically significant at p < 0.05. Spearman’s correlation analysis was used to determine relationship between miR-221 expression and DNM3 gene expression.
Results
Elevated tissue and exosomal miR-221 levels is positively correlated with glioma grades
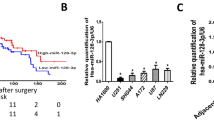
Firstly, glioma tissues were collected from patients with different grades. The miR-221 expression was evaluated by qRT-PCR. As shown in Fig. 1a, an increasing level of miR-221 can be seen in high-grade glioma tissue. In line with this, when cell-free miR-221 level and exosomal miR-221 level in serum were separately analyzed, a significant higher miR-221 level was also observed in glioma patients than normal patients (Fig. 1b). More importantly, serum exosomal miR-221 level increased with the glioma grades (Fig. 1c). In addition, we tested several glioma cell lines with different metastatic potential. Interestingly, miR-221 expression correlated with metastatic potential of the cancer cells (HEB < SHG-44 < U251 < U87MG; Fig. 1d).

Exosomal miR-221 expression levels are elevated in the serum of glioma patients and in the supernatant of glioma cell lines. a Relative miR-221 expression levels were analyzed by qRT-PCR in normal (n = 11) and glioma tissues (n = 48). U6 RNA levels were used as an internal control. b The box plot compares the relative values of cell-free and exosomal miR-221 in the serum of cancer-free (normal, n = 11) and glioma patients (n = 48). **Compared to cell-free/normal group, ##compared to exosome/normal group. c Relative expression levels of exosomal miR-221 in different stages of cancer tissues. d Relative expression levels of exosomal miR-221 in the supernatant of different glioma cell lines. Data were presented as the means ± SD from three independent experiments with triple replicates per experiment. **p < 0.01, ##p < 0.01
Inhibiting endogenous miR-221 expression and incubation with U87MG-derived exosome exerted opposite effect on glioma proliferation, migration and TMZ resistance
We next examined if downregulating miR-221 expression will affect the proliferation, migration and drug resistance of glioma. To this end, miR-221 siRNA (anti-miR-221) or control non-coding siRNA (anti-miR-NC) was introduced into SHG-44 cells and the effects on cell proliferation, migration and TMZ resistance were evaluated. RT-PCR was used to confirm the change of miR-221 levels after siRNA treatment (Fig. S1). Phenotypically, cells transfected with anti-miR-221 exhibited a decrease in proliferation, whereas transfection with anti-miR-NC did not induce such inhibitory effect on proliferation. In contrast, incubation with exosomes extracted from U87MG, which upregulated miR-221 expression (Fig. S1), promoted the proliferation of SHG-44 cells (Fig. 2a). Similarly, anti-miR-221 transfection inhibited the migration of SHG-44 cells as revealed by transwell (Fig. 2b) and scratch wound assay (Fig. 2c), which was not seen in cells transfected with anti-miR-NC. Unsurprisingly, incubation with U87MG-derived exosomes promoted the cell migration (Fig. 2b, c). Further, we exposed SHG-44 cells to 100 µM of TMZ, which induced cell apoptosis (Fig. 2d). However, cells transfected with anti-miR-221 suffered substantially higher apoptotic rate than cells transfected with anti-miR-221, and addition of exosomes extracted from U87MG exerted antiapoptotic effect on SHG-44 cells (Fig. 2d). It is worth noting that cells exposed to both anti-miR-221 and exosomes demonstrated phenotypical characteristics similar to those exposed to no treatments, and RT-PCR analysis revealed that anti-miR-221 and exosomes appeared to counteract each other in regulating miR-221 levels. Collectively, these data suggest that miR-221 has a pro-oncogenic role in glioma cells.

Downregulation of endogenous miR-221 inhibits the exsome induced cell functions and drug resistance of glioma cells. SHG-44 cells were transfected with anti-miR-221 inhibitor (Anti-miR-221), or control anti-sense RNA (anti-miR-NC) and cultured with the exosomes separated from the supernatant of U87MG or not. a CCK-8 assay was used to detected cell viability. **Compared to Anti-miR-221 group, #, ##compared to Anti-miR-221 group. &, &&compared to Anti-miR-NC + Exo. group. b Cells were treated as above, then a sterile 10 μL pipette tip was used to scratch the cells to form a wound when the cell densities were about 90% confluence. c Transwell invasion assay of the cells overexpressing miR-221 cells with or without DNM3 overexpression. After being fixed, the cells in the bottom of the invasion chamber were measured by the absorbance at 570 nm. d Cells were treated with 100 μM temozolomide or not, and cell apoptosis was analyzed by flow cytometry after 48 h. **p < 0.01, #p < 0.05, ##p < 0.01, &p < 0.05 and &&p < 0.01
Exosomal miR-221 directly targets and inhibits DNM3 expression
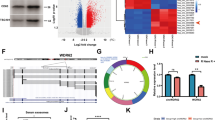
Given that miR-221 promotes glioma cell proliferation, migration and drug resistance, we subsequently investigated which oncogene is regulated by miR-221. Bioinformatic analysis predicted that wild-type DNM3 gene possesses two putative binding sites for miR-221, which were termed BS1 and BS2 regions respectively, while unpaired oligonucleotides occur exactly at these sequences in mutant DNM3 gene (Fig. 3a). Therefore, these regions in DNM3 could be targets of miR-22. To confirm this, we performed two-luciferase reporter assay for both wild-type DNM3 gene and mutant DNM3 gene spanning these two regions (Fig. S1B, Fig. 3b; Fig. S2B, Fig. 3c), and found that miR-221 expression downregulated the luciferase activity in wild-type DNM3 gene containing these two regions, but not the mutant DNM3 gene, indicating that BS1 and BS2 in wild-type DNM3 are the two regions that miR-221 interacts with. Indeed, DNM3 gene expression was found to be inversely correlated with the glioma grades (Fig. 3d) and Spearman’s analysis indicated a negative correlation between miR-221 and DNM3 expression (Fig. 3e). RT-PCR and western blot analysis also indicated that overexpression of miR-221 downregulated DNM3 gene expression (Fig. 3f, g). Similar effect can be achieved by incubating cells with exosomes extracted from U87MG, in which higher concentration of exosomes downregulated DNM3 mRNA and protein levels (Fig. 3h, i). Together, these evidences indicated that miR-221 targets and inhibits DNM3 expression.

Exosomal miR-221 directly targets and inhibits DNM3 expression. a Two putative seed-matching sites (in bold and italic) or mutant sites (red) between miR-221 and 3′-UTR of DNM3. b, c Luciferase reporter assay was performed to detect the relative luciferase activities of WT and mut DNM3 reporters. Renilla luciferase vector was used as an internal control. d Relative expression levels of miR-221 in different stages of cancer tissues. e Spearman’s correlation analysis was used to determine the correlation between the expression levels of DNM3 and miR-221 in human glioma specimens. f, g miR-221- and miR-NC-expression cells were subjected to qRT-PCR and western blotting and detected for DNM3 expression levels. h, i SHG-44 cells were cultured with different concentration (0, 50, 100 μg/mL) of exosomes separated from the supernatant of U87MG. After 48 h, cells were subjected to qRT-PCR and western blotting and detected for DNM3 expression levels. **p < 0.01
Overexpression of DNM3 reverses the stimulative effects of miR-221
Since DNM3 is a target of miR-221, we reasoned if ectopic expression of DNM3 would revert the stimulative effect of miR-221 on cell proliferation, migration and TMZ resistance. Thus, DNM3 gene was overexpressed in cells already transfected with vectors expressing miR-221 or miR-NC. As shown in Fig. 4a, b, while miR-221 transfection downregulated DNM3 expression, DNM3 ectopic expression upregulated DNM3 expression significantly. Notably, while miR-221 promoted cell proliferation, migration, and drug resistance (Fig. 4c–f), overexpression of miR-221 apparently compromised these effects (Fig. 4c–f).

Overexpression of DNM3 reverses the stimulative effects of miR-221. miR-221- or miR-NC-overexpression cells were transfected with vector or DNM3. a, b After 48 h, cells were subjected to qRT-PCR and western blotting and detected for DNM3 expression levels. c CCK-8 assay was used to detected cell viability. d Cells were treated as above, then a sterile 10 μL pipette tip was used to scratch the cells to form a wound when the cell densities were about 90% confluence. e Transwell invasion assay of the cells overexpressing miR-221 cells with or without DNM3 overexpression. After being fixed, the cells in the bottom of the invasion chamber were measured by the absorbance at 570 nm. f Cells were treated with 100 μM temozolomide or not, and cell apoptosis was analyzed by flow cytometry after 48 h. **p < 0.01 compared to miR-NC + vector group. ##p < 0.01 compared to miR-221 + vector group
RELA mediates exosomal miR-221 expression
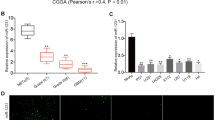
We were also interested in how miR-221 is regulated in glioma cells. RELA is a well-documented regulator of a variety of oncogenic pathways. Thus, the RELA levels in clinical samples were evaluated. It was found that RELA expression was positively correlated with the clinical grade of glioma (Fig. 5a). RELA expression also increased with the aggressiveness of glioma cells (Fig. 5b). Inspired by this, we performed bioinformatics studies and found that miR-221 is regulated by RELA promoter in a region spanning −1082 and −1073 (Fig. 5c). Promoter dual-luciferase reporter assay also indicated that RELA was able to promote wild-type miR-221 expression, rather than mutant miR-221 expression (Fig. 5d). When SHG-44 cells were transfected with RELA-expressing vector, miR-221 in cells was upregulated (Fig. 5e). This also holds true for exosomal miR-221 expression when SHG-44 receive RELA overexpression (Fig. 5f). On the contrary, both cell-level and exosomal miR-221 is downregulated when SHG-44 cells were exposed to RELA shRNA, which directly led to decreased RELA expression, cell proliferation, migration and resistance to TMZ (Fig. S2A–E). Collectively, these data indicated that miR-221 expression is regulated by RELA.

RELA mediated the overexpression of exosomal miR-221. a Relative expression levels of RELA in different stages of cancer tissues. **p < 0.01. b Relative RELA expression levels of different glioma cell lines. **p < 0.01. c Schematic diagrams show the potential RELA binding sites of miR-221 promoters boxes indicate the distribution of the putative RELA binding sites (BS). d Promoter luciferase reporter assay was performed on U87 to detect the relative luciferase activities of WT and mut PGL3-miR-221reporters. Renilla luciferase vector was used as an internal control. **p < 0.01. e, f SHG-44cells were transfected with RELA or shRELA and paired empty vectors. After 48 h, expression levels of cell and exosomal miR-221 were determined by qRT-PCR and U6 levels were used as internal control, and normalized to the values of control. **p < 0.01, ##p < 0.01
MiR-221 upregulation promotes tumor growth in vivo
To validate the regulatory role of miR-221 in tumor growth, we established the stable miR-221 overexpressing SHG-44 cell line, which were then inoculated in nude mice to initiate human glioma xenografts. As a control, cells transfected with miR-NC were also used for tumor inoculation. qRT-PCR was used to confirm the miR-221 overexpression in miR-221 transduced cells (Fig. 6a). As shown in Fig. 6b, c, miR-221 overexpressing tumors demonstrated a much faster increase in tumor size. Accordingly, the miR-221 overexpressing tumors collected at 20 days after inoculation weighted over two-times higher than the control tumors (Fig. 6d). Therefore, consistent with our observations that miR-221 demonstrated a tumor-promoting role in vitro, miR-221 is also a pro-tumorigenic factor in vivo.

miR-221 promotes tumor growth. The SHG-44 cells were infected with miR-221 or miR-NC lentivirus to establish stable cell lines. a Relative miR-221 expression levels of the stable cells were analyzed by qRT-PCR and normalized to vector group. **p < 0.01. b–d SHG-44/miR-221 and SHG-44/miR-NC cells were dispersed in 100 μL of serum-free 1640 medium and were subcutaneously injected into each side of posterior flank of the nude mice (n = 6). Tumors were measured every 4 days since they were apparently seen and the volumes were calculated using the following formula: volume = 0.5 × length × width2. The tumor was excised and weighed after 20 days with representative pictures of tumors shown (bar 10 mm)
Discussion
While miR-221 may serve a changeable role (anti- or pro-oncogenic) in a variety cancers, here we clarified that miR-221 is a pro-oncogenic factor in glioma. To our knowledge, our study represents the first study that evaluates miR-221 expression clinical glioma samples. We demonstrated that gliomas with higher grades contains a higher level of miR-221. More importantly, we verified that exosomal miR-221 in serum increased with glioma grades. This finding is of critical clinical relevance since miR-221 in circulating exosomes can be analyzed through a simple blood draw, enabling simple and non-invasive evaluation of glioma risk, which is highly desired and patient friendly. Indeed, blood tests for cancer risk-stratification have already been developed based on the miRNA differential expression [16]. The increased level of exosomal miR-221 in serum can enable early glioma detection, accurate risk-stratification and prognosis. This strategy is superior to biopsy, an invasive histopathological analysis, or costly imaging methods, such as computed tomography (CT), magnetic resonance imaging (MRI), etc. Considering the lack of miRNAs as a serum biomarker for glioma diagnosis, the establishment of exosomal miR-221 as a glioma biomarker may potentially lead to a more accurate and efficient clinical management of glioma patients.
Another issue that hinders effective glioma treatment is the frequent drug resistance and subsequent recurrence. Interestingly, we show that miR-221 level is closely correlated with the TMZ resistance of glioma cells. TMZ is an alkylating agent that methylates the O6 position of guanines, which leads to mismatch with thymine during DNA replication and eventually result in cell apoptosis, and is the first-line treatment regimen for glioma patients. However, GBM patients frequently develop TMZ resistance, and current glioma therapy is significantly hurdled by the lack of strategy to predict which patients tend to develop resistance to TMZ and subsequent failure to provide effective and timely treatment. Recently, miRNAs are thought to be associated with acquisition of TMZ resistance [17]. The data in our study suggests that high miR-221 level in glioma patients may predict that patients are likely to develop TMZ resistance. Based on this, evaluation of miR-221 level prior to and during TMZ treatment may be desired so that optimized therapeutic strategy can be employed.
On the other hand, the elevated miR-221 expression in high-grade and TMZ-resistant glioma may potentially justify the use of anti-miR-221 strategy to downregulate miR-221 expression to induce tumor growth retardation and sensitize cells to TMZ treatment. In the present study, anti-miR-221 was transfected, which effectively inhibited proliferation, migration and TMZ resistance of SHG-44 cells. Incorporated with current gene delivery technologies, such as viral and non-viral vehicles [18], inhibition of miR-221 in vivo could be a viable treatment option for GBM patients. This therapeutic strategy also promises to overcome toxicity associated with current chemotherapy drugs. Here we did not investigate how anit-miR-221 exactly modulates TMZ-resistance. Further studies on the interaction of miR-221 and TMZ-resistance factors, such as O6-methylguanine-DNA methyltransferase (MGMT), are warranted [19].
Despite the important role of miR-221 in regulating cancer progression, our understanding about the targets of miR-221 is still little. A number of genes have been proposed being regulated by miR-221, including p27 [20], PUMA [13] and Cx43 [14]. In this study, we found that two putative binding sites for miR-221 exist in DNM3 gene indicating that DNM3 is a target of miR-221. DNM3 is a member of the dynamin family of genes that play an important role in endocytosis and possess mechanochemical properties utilized to tabulate and sever membranes [21]. To date, few studies have investigated the impact of deregulated DNM3 gene in malignant disease. First we demonstrated that DNM3 expression was inversely correlated with the glioma grades. This mirrors previous findings that DNM3 gene is downregulated in hepatocellular cancer [15, 22]. In support of this, upregulating miR-221 in SHG-44 cells downregulated DNM3 expression. More interestingly, ectopic DNM3 overexpression compromised the tumor-promoting effect of miR-221. Indeed, a negative correlation was found between miR-221 and DNM3 expression. This finding can be readily translated to that DNM3 overexpression can be used to sensitize glioma cells to TMZ treatment. Thus, DNM3 may be another useful therapeutic target in glioma.
We also clarified that miR-221 expression is regulated by RELA. RELA is a key protein in the NF-ĸB pathway, which is an oncogenic driver in many cancer sites [23]. Nuclear translocation of RELA leads to binding of RELA to miR-221 promoter, activating miR-221 overexpression. This is verified by ectopic RELA expression, which promoted miR-221 expression, and transfection with shRELA, which inhibited miR-221 expression. Given that miR-221 is a pro-oncogenic gene, this finding is in accordance to the stimulative role of RELA in cancer. Interestingly, it has also been indicated that DNM3 is also regulated by RELA [15], but the precise mechanism of this regulation is unknown. Therefore, our result suggests that miR-221 may “connect the dots” between RELA and DNM3. However, further studies are necessary to validate this hypothesis.
Conclusions
miR-221 is a biomarker that positively correlates with glioma grade and TMZ resistance. DNM3 is a target of miR-221 and miR-221 is regulated by RELA. The miR-221/RELA axis may serve as a novel therapeutic target for glioma patients.
References
Omuro A, DeAngelis LM (2013) Glioblastoma and other malignant gliomas: a clinical review. JAMA 310:1842–1850. doi:10.1001/jama.2013.280319
DeAngelis LM (2001) Brain tumors. N Engl J Med 344:114–123
Louis DN (2006) Molecular pathology of malignant gliomas. Annu Rev Pathol Mech Dis 1:97–117
Calin GA, Croce CM (2006) MicroRNA-cancer connection: the beginning of a new tale. Cancer Res 66:7390–7394
Ciafre S, Galardi S, Mangiola A, Ferracin M, Liu C-G, Sabatino G, Negrini M, Maira G, Croce C, Farace M (2005) Extensive modulation of a set of microRNAs in primary glioblastoma. Biochem Biophys Res Commun 334:1351–1358
Azmi AS, Bao B, Sarkar FH (2013) Exosomes in cancer development, metastasis, and drug resistance: a comprehensive review. Cancer Metastasis Rev 32:623–642
Yang C, Robbins PD (2011) The roles of tumor-derived exosomes in cancer pathogenesis. Clin Dev Immunol 2011:842849
Liu M, Liu J, Wang L, Wu H, Zhou C, Zhu H, Xu N, Xie Y (2014) Association of serum microRNA expression in hepatocellular carcinomas treated with transarterial chemoembolization and patient survival. PloS One 9:e109347
Tao K, Yang J, Guo Z, Hu Y, Sheng H, Gao H, Yu H (2014) Prognostic value of miR-221-3p, miR-342-3p and miR-491-5p expression in colon cancer. Am J Transl Res 6:391–401
Yau T, Wu C, Dong Y, Tang C, Ng S, Chan F, Sung J, Yu J (2014) MicroRNA-221 and microRNA-18a identification in stool as potential biomarkers for the non-invasive diagnosis of colorectal carcinoma. Br J Cancer 111:1765–1771
Zheng Q, Peskoe SB, Ribas J, Rafiqi F, Kudrolli T, Meeker AK, De Marzo AM, Platz EA, Lupold SE (2014) Investigation of miR-21, miR-141, and miR-221 expression levels in prostate adenocarcinoma for associated risk of recurrence after radical prostatectomy. Prostate 74:1655–1662
Chen L, Zhang J, Han L, Zhang A, Zhang C, Zheng Y, Jiang T, Pu P, Jiang C, Kang C (2012) Downregulation of miR-221/222 sensitizes glioma cells to temozolomide by regulating apoptosis independently of p53 status. Oncol Rep 27:854–860. doi:10.3892/or.2011.1535
Zhang C-Z, Zhang J-X, Zhang A-L, Shi Z-D, Han L, Jia Z-F, Yang W-D, Wang G-X, Jiang T, You Y-P (2010) MiR-221 and miR-222 target PUMA to induce cell survival in glioblastoma. Mol Cancer 9:1
Hao J, Zhang C, Zhang A, Wang K, Jia Z, Wang G, Han L, Kang C, Pu P (2012) miR-221/222 is the regulator of Cx43 expression in human glioblastoma cells. Oncol Rep 27:1504
Zhang Z, Chen C, Guo W, Zheng S, Sun Z, Geng X (2016) DNM3 attenuates hepatocellular carcinoma growth by activating P53. Med Sci Monit 22:197–205. doi:10.12659/msm.896545
Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K, Guo J, Zhang Y, Chen J, Guo X (2008) Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res 18:997–1006
Ujifuku K, Mitsutake N, Takakura S, Matsuse M, Saenko V, Suzuki K, Hayashi K, Matsuo T, Kamada K, Nagata I, Yamashita S (2010) miR-195, miR-455-3p and miR-10a(*) are implicated in acquired temozolomide resistance in glioblastoma multiforme cells. Cancer Lett 296:241–248. doi:10.1016/j.canlet.2010.04.013
Pereira DM, Rodrigues PM, Borralho PM, Rodrigues CM (2013) Delivering the promise of miRNA cancer therapeutics. Drug Discov Today 18:282–289
Kohsaka S, Wang L, Yachi K, Mahabir R, Narita T, Itoh T, Tanino M, Kimura T, Nishihara H, Tanaka S (2012) STAT3 inhibition overcomes temozolomide resistance in glioblastoma by downregulating MGMT expression. Mol Cancer Ther 11:1289–1299. doi:10.1158/1535-7163.MCT-11-0801
Zhang C, Kang C, You Y, Pu P, Yang W, Zhao P, Wang G, Zhang A, Jia Z, Han L (2009) Co-suppression of miR-221/222 cluster suppresses human glioma cell growth by targeting p27kipl in vitro and in vivo. Int J Oncol 34:1653
Orth JD, McNiven MA (2003) Dynamin at the actin–membrane interface. Curr Opin Cell Biol 15:31–39
Inokawa Y, Nomoto S, Hishida M, Hayashi M, Kanda M, Nishikawa Y, Takeda S, Fujiwara M, Koike M, Sugimoto H, Fujii T, Nakayama G, Yamada S, Tanaka C, Kobayashi D, Kodera Y (2013) Dynamin 3: a new candidate tumor suppressor gene in hepatocellular carcinoma detected by triple combination array analysis. Onco Targets Ther 6:1417–1424. doi:10.2147/OTT.S51913
Hwang SK, Baker AR, Young MR, Colburn NH (2014) Tumor suppressor PDCD4 inhibits NF-kappaB-dependent transcription in human glioblastoma cells by direct interaction with p65. Carcinogenesis 35:1469–1480. doi:10.1093/carcin/bgu008
Acknowledgements
The authors would like to thank Dr. Li-qun Wang, Dr. Zhong-qiang Lv, and Dr. Zong-mao Zhao for their technical advice.
Funding
This study was supported by the Program of Graduate students innovation fund of Hebei Province.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Ethical approval
The protocols approved by the Committee on the Ethics of Animal Experiments of The Second Hospital of Hebei Medical University.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Yang, JK., Yang, JP., Tong, J. et al. Exosomal miR-221 targets DNM3 to induce tumor progression and temozolomide resistance in glioma. J Neurooncol 131, 255–265 (2017). https://doi.org/10.1007/s11060-016-2308-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11060-016-2308-5