
Abstract
Werner syndrome (WS) is a premature aging disorder characterized by early onset of symptoms related to normal aging and by a high predisposition to various types of cancer, including gliomas. WS is caused by inherited recessive mutations in the WRN gene, which encodes a helicase considered a caretaker of the genome. Aiming to study the role of WRN Cys1367Arg in glioma susceptibility and oncologic prognosis of patients, we investigated the genotype distribution of this single nucleotide polymorphism in 94 glioma patients and 100 healthy subjects. Comparisons of genotype distributions and allele frequencies did not reveal any significant difference between the groups. Overall and disease-free survival rates were calculated, but no statistically significant difference was observed. Our data suggest that WRN Cys1367Arg SNP is not involved either in susceptibility to developing gliomas or in patient survival, at least in the Brazilian population.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Gliomas are the most common tumors of the central nervous system. In spite of the marked advances in the characterization of the molecular pathogenesis of gliomas, these tumors remain incurable and, in most of the cases, resistant to treatments, due to their molecular heterogeneity [1].
Werner syndrome (WS) is a premature aging disorder characterized by early onset of symptoms related to normal aging, including coronary artery disease, atherosclerosis, osteoporosis, and diabetes mellitus [2–4]. WS is caused by inherited recessive mutations in the WRN gene, which encodes a member of the RecQ family of helicases involved in DNA replication and in maintaining the integrity of the genome [5, 6]. Therefore, the cells of WS patients show a high level of chromosomal translocations and deletions, and these patients exhibit an increased predisposition to various types of cancer, including gliomas [7, 8]. However, despite its putative tumor suppressor function, little is known about the contribution of the WRN protein to human sporadic malignancies.
Taking into account that almost all cancers occur in the elderly and that mutations in the WRN gene lead to accelerated aging, it has been reasoned that polymorphisms of the WRN gene might also be associated with age-related pathologies and cancer predisposition. In particular, a single nucleotide polymorphism (SNP) that leads to a cysteine-to-arginine substitution in WRN codon 1367 (Cys1367Arg) has attracted a considerable amount of attention from different research groups [9–16].
Based on a case–control analysis of glioma patients and healthy controls, we investigated, using the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) approach, the role of WRN Cys1367Arg SNP as a potential risk factor and/or prognostic marker of glioma.
Materials and methods
Study population
A total of 94 gliomas were analyzed, which had been surgically resected from previously untreated patients under the care of the Neurosurgery Department of the Pio XII Foundation – Cancer Hospital of Barretos (Barretos, SP, Brazil). These samples were used in other, previously reported genetic studies [17, 18]. Tumor types and stages were determined by two experienced pathologists, according to WHO criteria [19]. The clinical outcome, including length of survival, was obtained from patient records and by contacting each patient’s general practitioner. The mean follow-up period for all patients was 47.62 weeks (range = 0.12–118.14). Blood samples of 100 healthy individuals were collected as controls. Because of the highly heterogeneous ethnic composition of the Brazilian population, the individuals of the control group were randomly selected from the general population of São Paulo State, with no family history of cancer in first-degree relatives. The control sample was matched for gender and mean age with the patient group. The mean age of both patient and control groups was 45 years, and 69.14% of patients and 69% of controls were 40 years old or older. Informed consent was given from all recruited cancer patients and healthy controls. The study was previously approved by the Ethics Committee of the University Hospital of the Ribeirão Preto School of Medicine, University of São Paulo.
DNA extraction and primer design
DNA extraction was performed using proteinase K and phenol–chloroform, according to routine molecular biology protocols. Primers were designed using the Gene Runner 3.05 program (Hasting Software, Inc.) from the WRN Cys1367Arg SNP sequence, obtained in the dbSNP of NCBI (ID# rs1346044). A 119 bp sequence containing the polymorphic site was amplified by PCR using the designed primers: forward 5′-GAC ACG TAC CTT ATC CAC ATG G-3′ and reverse 5′-GAA CAG ATC TCT TCA GAA CCG G-3′.
Genotyping
PCR was carried out in a final volume of 25 μl containing 50 ng of genomic DNA template, 1× PCR buffer with 2 mM MgCl2, 0.4 μM of each primer (Invitrogen), 50 μM dNTPs (Amersham Biosciences), and 0.5 U DNA polymerase (Biotools). The PCR cycling conditions were: 5 min at 94°C, followed by 35 cycles of 30 s at 94°C, 30 s at 58°C, and 30 s at 72°C, and a final elongation cycle at 72°C for 5 min.
For RFLP, the PCR products were digested with PmlI (4 U at 37°C for 4 h) (New England BioLabs). PmlI recognizes a restriction site of the Arg1367 allele (CAC^GTG) and generates two fragments of different sizes (49 and 70 bp, respectively), while the Cys1367 allele is not cleaved and keeps the original PCR product (119 bp). DNA fragments were electrophoresed on a 10% acrylamide:bisacrylamide gel (19:1), and then stained with silver nitrate.
For quality control, the genotypes of 10% of the samples were reassessed to confirm the results. Also, selected PCR products were purified and submitted to bidirectional sequencing, to further confirm the reliability of the genotype analysis. The PCR products were purified with ExoSAP (USB), followed by sequencing with a DYEnamic ET Dye Terminator Kit (Amersham Biosciences), according to the manufacturer’s instructions. Sequencing reactions were performed in a MegaBACE 1000 sequencing system (GE Healthcare).
Statistical analysis
The chi-square test was used to verify whether genotype distributions were in Hardy–Weinberg equilibrium. Observed genotype and allele frequencies in gliomas were compared to controls using the chi-square and Fisher’s exact tests, respectively. Kaplan–Meier curves were constructed to assess overall survival and disease-free survival rates, and differences among groups were analyzed by the log-rank test. Odds ratio (OR) and 95% confidence interval (CI) were calculated using a logistic regression model. Statistical significance was set at P < 0.05. Statistical analyses were performed with softwares S-Plus 2000 (Insightful, Inc.) and GraphPad Prism 4.0 (GraphPad Software, Inc.).
Results
A total of 94 glioma patients and 100 control subjects were included in this study. The patient sample comprised 59 males and 35 females (M/F ratio = 1.69) and the control sample consisted of 63 males and 37 females (M/F ratio = 1.7). Mean age in both patient and control groups was 45 years (range = 1–75 and 18–72, respectively). No significant gender- or age-related differences were observed between the groups (P > 0.05). Eighty-one patients had astrocytomas (11 grade I, 23 grade II, 8 grade III, and 39 grade IV glioblastomas), five had oligodendrogliomas (3 grade II and 2 grade III), and eight had ependymomas (5 grade II and 3 grade III). Forty-two of the 94 gliomas were classified as low-grade (grades I and II) and 52 as high-grade (grades III and IV). No significant difference in stratification by tumor grade was observed between the groups (P = 0.211).
Genotype and allele frequencies were in Hardy–Weinberg equilibrium in all groups. Allele and genotype frequencies of WRN Cys1367Arg SNP did not differ significantly between case and control groups. Arg1367 allele frequency in controls was 0.22, while in patients it was 0.26 (P = 0.514). The frequencies of genotypes Cys/Cys, Cys/Arg, and Arg/Arg in controls were 59%, 38% and 3%, while in patients they were 51.1%, 45.7% and 3.2%, respectively (P = 0.534) (Table 1).
Compared to Cys/Cys, the most common genotype in the study population, both the Cys/Arg and the Arg/Arg genotypes did not influence the risk of glioma development (OR = 1.39; 95% CI, 0.78–2.48; P = 0.332; and OR = 1.23; CI, 0.24–6.37; P = 0.861, respectively). Genotypes Cys/Arg and Arg/Arg were combined here because of the low frequency of Arg/Arg, and showed no influence on the risk of developing gliomas (OR = 1.38; 95% CI, 0.78–2.43; P = 0.334) (Table 2).
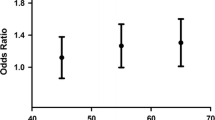
The mean follow-up period for all patients was 47.62 weeks (range = 0.12–118.14). For the 53 patients who survived the follow-up period (censored patients), the mean follow-up time was 62.07 weeks; for the 41 patients who died during the follow-up period, it was 28.93 weeks. The survival curves for overall survival (Fig. 1a) and disease-free survival (Fig. 1b) of the patients were not significantly different among the WRN Cys1367Arg SNP genotypes (P > 0.05). Once all three patients with genotype Arg/Arg died during the follow-up period (Fig. 1a), the disease-free survival curve was obtained only from patients with genotypes Cys/Cys and Cys/Arg (Fig. 1b).

Survival curves show no correlation between survival and the WRN Cys1367Arg SNP genotype. (a) Overall survival (P = 0.396); (b) disease-free survival (P = 0.843)
Discussion
In addition to premature aging, patients with WRN germline mutations develop a broad spectrum of tumors, which is one of the main causes of their death before the age of 50. This is the reason why a tumor suppressor function for WRN has been proposed. This putative role is also supported by a very high rate of loss of heterozygosity at the chromosomal WRN loci at 8p11.2–p12 in many tumor types, including colorectal and breast cancers [20, 21]. Gene WRN encodes a protein with 1,432 amino acids that possesses both 3′ → 5′ DNA helicase and 3′ → 5′ DNA exonuclease activities. DNA helicases are enzymes that unwind the energetically stable double-stranded structure of DNA, to provide a single-stranded template for important cellular processes such as replication, base excision repair, homologous recombination, and telomere maintenance [4, 22, 23]. Once somatic mutations of WRN have not been described in sporadic neoplasms, the analysis of SNPs within the WRN gene represents a promising strategy for association studies between Cys1367Arg and cancer.
In the present case–control study, we investigated the relationship between WRN Cys1367Arg SNP and risk of glioma and patient survival. The genotype and allele frequencies in case and control groups were similar, showing no significant association with glioma (P = 0.534 and P = 0.514, respectively).
Several epidemiological studies have already been carried out on WRN Cys1367Arg SNP. Most of these studies focused on age-related pathologies, including atherosclerosis, osteoporosis, diabetes mellitus, myocardial infarction, coronary artery disease, and Alzheimer’s disease [2, 9, 10, 12, 13, 24–28]. Two Japanese research groups were the first ones to associate WRN Cys1367Arg with a complex disease [24, 25]. They reported an association between the wild-type genotype Cys/Cys and myocardial infarction in the Japanese population, and a protective role for the minor Arg1367 allele (frequency among controls, 0.08 and 0.09). Moreover, the protective role of Arg1367 was supported by the fact that in populations with increased frequencies of this allele, like the Finnish, Mexican, and North American, the risk for myocardial infarction was not observed (Arg1367 among controls, 0.21–0.30) [27]. On the other hand, in Japan, a study reported that the more frequent Cys1367 allele has been associated with a lower frequency of osteoporosis in postmenopausal women [28]. In this study, the Arg1367 allele seems to act as a vulnerability factor (Arg1367 among controls, 0.21–0.30). Another Japanese study found that the minor allele Arg1367 may be associated with a lower risk of type 2 diabetes mellitus [10], whereas in the Netherlands and in Brazil no significant association of WRN Cys1367Arg with age-related morbidity and Alzheimer’s disease was found, possibly reflecting the increased frequency of the minor Arg1367 allele in those populations (Arg1367 among controls, 0.21–0.30) [9, 12, 13].
It is important to notice that all the reports listed above, except one [28], showed protective effects of the Arg1367 allele against the various diseases associated with the WS phenotype. The same held true for our observations on gliomas, once the frequency of the minor allele in the present study (Arg1367 among controls, 0.22) was similar to previous studies reporting its protective role.
Based on the observation that all known WRN mutations identified so far result in truncation of the C-terminal domain of the WRN protein, the binding site of protein TP53 [29], it has been proposed that lack of the C-terminal nuclear localization signal is important in the pathogenesis of WS [28, 30, 31]. Although WRN Cys1367Arg is located only three amino acids apart from the nuclear localization signal motif, previous studies failed to detect any significant difference between Cys1367Arg alleles with respect to their nuclear localization [2], or helicase and helicase-coupled exonuclease activity [32]. However, other possible explanations for the observed association include the allelic difference of WRN Cys1367Arg in the interactions with other proteins, as well as the presence of unknown functionally responsible SNPs that are in linkage disequilibrium with Cys1367Arg in different populations and phenotypes [33]. Certainly, difficulties observed in genotype-phenotype associations are not an exclusive feature of age-related pathologies. With regard to cancer, reports tend to be even more heterogeneous, as described below.
To date, only five published studies have examined the association between WRN Cys1367Arg and cancer, however, none of them with brain tumors. Shen et al. [34] reported an association of the Arg1367 allele with decreased risk of non-Hodgkin lymphoma among women in North America (OR = 0.71; 95% CI, 0.56–0.91; P = 0.007). Recently, based on a case–control study performed in Japan, Nakayama et al. [33] also suggested a protective effect of the Arg1367 allele against bone and soft tissue sarcomas (OR = 0.66; 95% CI, 0.49–0.88; P = 0.005). The association of WRN Cys1367Arg SNP with breast cancer was also analyzed by two independent groups. Wirtenberger et al. [14] suggested that the Arg1367 allele acts as a low-penetrance risk factor for familial breast cancer (OR = 1.28; 95% CI, 1.06–1.54; P = 0.010) and high-risk familial breast cancer (OR = 1.32; 95% CI, 1.06–1.65; P = 0.015) in Germany. However, Ding et al. [15] did not confirm these results in a case–control study in Japan (OR = 1.00; 95% CI, 0.83–1.23; P = 0.97). Finally, Khayat et al. [11] found no evidence of an association between Cys1367Arg and gastric adenocarcinoma in a Brazilian population (P > 0.05).
The present study is the first one to investigate the association between WRN Cys1367Arg and risk and oncologic prognosis of brain tumors, more specifically with regard to glioma. Our data show that neither glioma risk (OR = 1.38; 95% CI, 0.78–2.43; P = 0.334) nor patient survival (overall and disease-free survival, P = 0.396 and P = 0.843, respectively) were associated with variant alleles. Thus, our results corroborate those of Khayat et al. [11] and Ding et al. [15].
In conclusion, our results suggest that there is no association between WRN Cys1367Arg genotypes and susceptibility to glioma or patient survival, at least in the Brazilian population. Nevertheless, future studies on larger populations from other parts of the world are essential for a definitive conclusion.
References
Boudreau CR, Yang I, Liau LM (2005) Gliomas: advances in molecular analysis and characterization. Surg Neurol 64:286–294; discussion 294. doi:10.1016/j.surneu.2005.03.033
Bohr VA, Metter EJ, Harrigan JA et al (2004) Werner syndrome protein 1367 variants and disposition towards coronary artery disease in Caucasian patients. Mech Ageing Dev 125:491–496. doi:10.1016/j.mad.2004.05.001
Shen JC, Loeb LA (2000) The Werner syndrome gene: the molecular basis of RecQ helicase-deficiency diseases. Trends Genet 16:213–220. doi:10.1016/S0168-9525(99)01970-8
Yu CE, Oshima J, Fu YH et al (1996) Positional cloning of the Werner’s syndrome gene. Science 272:258–262. doi:10.1126/science.272.5259.258
Hickson ID (2003) RecQ helicases: caretakers of the genome. Nat Rev Cancer 3:169–178. doi:10.1038/nrc1012
Harrigan JA, Wilson DMIII, Prasad R et al (2006) The Werner syndrome protein operates in base excision repair and cooperates with DNA polymerase beta. Nucleic Acids Res 34:745–754. doi:10.1093/nar/gkj475
Kobayashi S, Gibo H, Sugita K et al (1980) Werner’s syndrome associated with meningioma. Neurosurgery 7:517–520. doi:10.1097/00006123-198011000-00020
Laso FJ, Vasquez G, Pastor I et al (1989) Werner’s syndrome and astrocytoma. Dermatologica 178:118–120
Payao SL, de Labio RW, Gatti LL et al (2004) Werner helicase polymorphism is not associated with Alzheimer’s disease. J Alzheimers Dis 6:591–594; discussion 673–581
Hirai M, Suzuki S, Hinokio Y et al (2005) WRN gene 1367 Arg allele protects against development of type 2 diabetes mellitus. Diabetes Res Clin Pract 69:287–292. doi:10.1016/j.diabres.2005.01.012
Khayat AS, Lobo Gatti L, Moura Lima E et al (2005) Polymorphisms of the TP53 codon 72 and WRN codon 1367 in individuals from Northern Brazil with gastric adenocarcinoma. Clin Exp Med 5:161–168. doi:10.1007/s10238-005-0081-4
Smith MA, Silva MD, Araujo LQ et al (2005) Frequency of Werner helicase 1367 polymorphism and age-related morbidity in an elderly Brazilian population. Braz J Med Biol Res 38:1053–1059. doi:10.1590/S0100-879X2005000700008
Kuningas M, Slagboom PE, Westendorp RG et al (2006) Impact of genetic variations in the WRN gene on age related pathologies and mortality. Mech Ageing Dev 127:307–313. doi:10.1016/j.mad.2005.11.005
Wirtenberger M, Frank B, Hemminki K et al (2006) Interaction of Werner and Bloom syndrome genes with p53 in familial breast cancer. Carcinogenesis 27:1655–1660. doi:10.1093/carcin/bgi374
Ding SL, Yu JC, Chen ST et al (2007) Genetic variation in the premature aging gene WRN: a case-control study on breast cancer susceptibility. Cancer Epidemiol Biomarkers Prev 16:263–269. doi:10.1158/1055-9965.EPI-06-0678
Castro E, Oviedo-Rodriguez V, Angel-Chavez LI (2008) WRN polymorphisms affect expression levels of plasminogen activator inhibitor type 1 in cultured fibroblasts. BMC Cardiovasc Disord 8:5. doi:10.1186/1471-2261-8-5
Pinto GR, Clara CA, Santos MJ et al (2007) Mutation analysis of gene PAX6 in human gliomas. Genet Mol Res 6:1019–1025
Pinto GR, Yoshioka FK, Silva RL et al (2008) Prognostic value of TP53 Pro47Ser and Arg72Pro single nucleotide polymorphisms and the susceptibility to gliomas in individuals from Southeast Brazil. Genet Mol Res 7:207–216
Kleihues P, Louis DN, Scheithauer BW et al (2002) The WHO classification of tumors of the nervous system. J Neuropathol Exp Neurol 61:215; discussion 226–219
Chughtai SA, Crundwell MC, Cruickshank NR et al (1999) Two novel regions of interstitial deletion on chromosome 8p in colorectal cancer. Oncogene 18:657–665. doi:10.1038/sj.onc.1202340
Armes JE, Hammet F, de Silva M et al (2004) Candidate tumor-suppressor genes on chromosome arm 8p in early-onset and high-grade breast cancers. Oncogene 23:5697–5702. doi:10.1038/sj.onc.1207740
Gray MD, Shen JC, Kamath-Loeb AS et al (1997) The Werner syndrome protein is a DNA helicase. Nat Genet 17:100–103. doi:10.1038/ng0997-100
Kamath-Loeb AS, Shen JC, Loeb LA et al (1998) Werner syndrome protein. II. Characterization of the integral 3′ → 5′ DNA exonuclease. J Biol Chem 273:34145–34150. doi:10.1074/jbc.273.51.34145
Ye L, Miki T, Nakura J et al (1997) Association of a polymorphic variant of the Werner helicase gene with myocardial infarction in a Japanese population. Am J Med Genet 68:494–498. doi :10.1002/(SICI)1096-8628(19970211)68:4<494::AID-AJMG30>3.0.CO;2-L
Morita H, Kurihara H, Sugiyama T et al (1999) A polymorphic variant C1367R of the Werner helicase gene and atherosclerotic diseases in the Japanese population. Thromb Haemost 82:160–161
Castro E, Edland SD, Lee L et al (2000) Polymorphisms at the Werner locus: II. 1074Leu/Phe, 1367Cys/Arg, longevity, and atherosclerosis. Am J Med Genet 95:374–380. doi :10.1002/1096-8628(20001211)95:4<374::AID-AJMG14>3.0.CO;2-4
Castro E, Ogburn CE, Hunt KE et al (1999) Polymorphisms at the Werner locus: I. Newly identified polymorphisms, ethnic variability of 1367Cys/Arg, and its stability in a population of Finnish centenarians. Am J Med Genet 82:399–403. doi :10.1002/(SICI)1096-8628(19990219)82:5<399::AID-AJMG8>3.0.CO;2-R
Ogata N, Shiraki M, Hosoi T et al (2001) A polymorphic variant at the Werner helicase (WRN) gene is associated with bone density, but not spondylosis, in postmenopausal women. J Bone Miner Metab 19:296–301. doi:10.1007/s007740170013
Blander G, Kipnis J, Leal JF et al (1999) Physical and functional interaction between p53 and the Werner’s syndrome protein. J Biol Chem 274:29463–29469. doi:10.1074/jbc.274.41.29463
Yu CE, Oshima J, Wijsman EM et al (1997) Mutations in the consensus helicase domains of the Werner syndrome gene. Am J Hum Genet 60:330–341
Matsumoto T, Shimamoto A, Goto M et al (1997) Impaired nuclear localization of defective DNA helicases in Werner’s syndrome. Nat Genet 16:335–336. doi:10.1038/ng0897-335
Kamath-Loeb AS, Welcsh P, Waite M et al (2004) The enzymatic activities of the Werner syndrome protein are disabled by the amino acid polymorphism R834C. J Biol Chem 279:55499–55505. doi:10.1074/jbc.M407128200
Nakayama R, Sato Y, Masutani M et al (2008) Association of a missense single nucleotide polymorphism, Cys1367Arg of the WRN gene, with the risk of bone and soft tissue sarcomas in Japan. Cancer Sci 99:333–339. doi:10.1111/j.1349-7006.2007.00692.x
Shen M, Zheng T, Lan Q et al (2006) Polymorphisms in DNA repair genes and risk of non-Hodgkin lymphoma among women in Connecticut. Hum Genet 119:659–668. doi:10.1007/s00439-006-0177-2
Acknowledgements
We thank Márcio Rogério Penha and Vanderci Massaro de Oliveira for technical support. Research supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Financiadora de Estudos e Projetos (FINEP CT-INFRA/FADESP) (Grant number: 1017-01). R. R. Burbano was the recipient of a PQ-2 fellowship (number 308256/2006-9) from CNPq.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pinto, G.R., Yoshioka, F.K.N., Clara, C.A. et al. WRN Cys1367Arg SNP is not associated with risk and prognosis of gliomas in Southeast Brazil. J Neurooncol 90, 253–258 (2008). https://doi.org/10.1007/s11060-008-9664-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11060-008-9664-8