
Abstract
Climate change models predict an increase in aridity in many parts of the world for the twenty-first century, which is likely to be more intense in the Mediterranean basin than in other regions. This study addresses the potential distribution of three Mediterranean pine species (Pinus pinea L., P. halepensis Mill. and P. pinaster Aiton) in southern Spain in response to the forecast increased aridity. Pines constitute a useful source of various types of raw materials, which has led to their increasing introduction around the world. The study was based on ecological niche modelling using multinomial logistic regression, over an area spanning about 8.7 million ha in the south of Spain. In total, 11 explanatory variables were included, drawing on measurements made at high resolution (200 m). Four different periods were studied: the reference period (1961–2000), early twenty-first century (2011–2040), middle twenty-first century (2041–2070) and late twenty-first century (2071–2100). Future time slices were analysed in three different scenarios: B1, A1b and A2 in the CNCM3 general circulation model. The results predict a wider distribution for stone pine, which could expand its potential area in southern Spain. In contrast, Aleppo pine, and especially cluster pine, would reduce their present distribution, with cluster pine occupying higher altitude sites while low altitude populations diminished. The validation model enables accurate maps to be produced, representing powerful tools for afforestation/reforestation programs in the future.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Projected trends in the context of global climate change suggest an increase in mean annual temperatures in the Mediterranean Basin. This increase is likely to exceed the global mean and lead to augmented drought by the end of the twenty-first century. In addition, annual rainfall is likely to decrease (IPCC 2007; Giorgi and Lionello 2008; García-Ruiz et al. 2011). These arguments reveal uncertainties on the ecological conditions of forest ecosystems to be restored (Jacobs et al. 2015). The probability of more stressful conditions in the Mediterranean Basin in the coming decades, together with the heterogeneous climate in Spain, make this area a suitable scenario for assessing the potential effects of climate change on terrestrial ecosystems (Márquez et al. 2011). This is especially true for southern Spain, which exhibits the largest climate and relief extremes in the Iberian Peninsula. Humans have had a strong impact on flora and vegetation, and are responsible for discontinuities in species distributions, shifts in environmental conditions and consequently habitat loss (Ibáñez et al. 2014). The restoration of degraded areas can be carried out by fast-growing pioneer pine species (Rejmánek and Richardson 1996). The most frequent pines in southern Spain are P. pinea L. (stone pine), P. halepensis Mill. (Aleppo pine) and P. pinaster Aiton (cluster pine), whose excellent performance under harsh climatic and edaphic conditions (Del Campo et al. 2014), facilitates the implementation of afforestation/reforestation programs.
In the nineteenth century, afforestation programs were developed to restore areas which had been anthropically and naturally degraded for centuries (Barbéro et al. 1998). In Spain, a large-scale reforestation program running from 1940 to 1995 led to the planting of approximately 3.5 million ha (Montero 1997), largely with pines (Zavala and Zea 2004). Indeed, in the Sierra Morena—the mountain range stretching the length of northern Andalusia—close to 90 % of all reforestation was carried out with pines (Muñoz Álvarez 2010). Also notable in this context was the cultivation of cluster pine for timber production in the second half of the twentieth century (González-Muñoz et al. 2014).
The naturalness of stone pine has been a matter of debate for centuries (Martínez et al. 2004). It is difficult to ascertain in some populations in the southwest of Spain (Stevenson 1985). Some sceptical authors believe that pines in the Iberian Peninsula are allochthonous and claim that they have never been regarded as phytosociologically natural vegetation (Ceballos and Ruiz de la Torre 1979; Rivas-Martínez 1987; Valle 2004). However, recent studies support a natural origin of these species in the western Mediterranean Basin (Martínez and Montero 2004). The largest stands of stone pine are located in the Iberian Peninsula, which accounts for 60 % of the world distribution (Gómez et al. 2002; Martínez et al. 2004). It is a similar case with cluster pine, the most widespread pine in the Iberian Peninsula, occupying more than 1,200,000 ha—half of which are afforestations (López González 2007). The natural distribution of this pine in the Iberian Peninsula has been established from palaeoecological pollen and archaeological charcoals (Carrion et al. 2000; Carrión and Díez 2004; García-Amorena et al. 2007; Rubiales et al. 2009; López-Sáez et al. 2010). Valle (2004) found an edaphoxerophilous vegetation series of cluster pine on peridotites and dolomites in southern Spain, in addition to an edaphoxerophilous complex of Aleppo pine in south-eastern Spain. These findings have led to stone pine, cluster pine and Aleppo pine being considered autochthonous (Soto et al. 2010). Pine forests were mainly concentrated in sites with poor-developed soils and under harsh climatic conditions (Cabezudo Artero and Pérez Latorre 2004). Largescale reforestation, however, has led to the introduction of pines in potential areas for oaks.
Ecological niche modelling (ENM) is used to correlate the current distribution of a given species with its general abiotic preferences (Olden et al. 2002; Guisan and Thuiller 2005; Araùjo and Guisan 2006; Elith et al. 2006; Mateo et al. 2011; Mellert et al. 2011). Use of the modelling approach to predict potential species distribution has increased in recent years as a useful tool for management, conservation and restoration (Hidalgo et al. 2008; Vessella and Schirone 2013; Bede-Fazekas et al. 2014; López-Tirado and Hidalgo 2014). Many studies using ENM have been carried out in the Iberian Peninsula (Alba-Sánchez et al. 2010; Felicísimo et al. 2012; García-Valdés et al. 2013), including studies on decline and growth as a result of drought (Sánchez-Salguero et al. 2010, 2012). In this work, we used logistic regression (LR), a type of generalized linear model (GLM). Because LR uses presence/absence data, it allows a model with a specific number of randomly chosen absences to be run. The main advantage of LR is the applicability of its formula to the whole territory so that explanatory variables can be related to the probabilities of presence (Sumarga 2011). If the dependent variable has more than two categories, then Multinomial Logistic Regression (MLR) is possible. This type of LR is used to study spatial vegetation dynamics by recording plant species or vegetation types as pixels on a grid (Augustin et al. 2001; Finley et al. 2009). The ensuing MLR formula can be subsequently used in the climate change scenarios for forecasting the suitability of each species in the study area.
The primary aims of this work are: (a) to obtain a high resolution map (200 m) of suitable areas for growth of the three target pine species in the south of Spain from the results of a GLM approach; (b) to implement this method in three different climatic scenarios (B1, A1b and A2) over the course of the twenty-first century; and (c) to predict the potential impact of climate change on the distribution of the target species and assess potential scenarios for future afforestation/reforestation programs.
Materials and methods
Study area
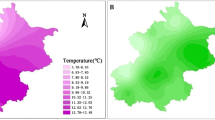
The study area spans ca. 8.7 million ha in the south of Spain (largely in the Andalusian region; Fig. 1) and exhibits the greatest extremes of rainfall and temperature in Spain. Sub-desert zones contrast with the wettest mountainous areas of the study area (De Castro et al. 2005). The region includes two mountain ranges, the Sierra Morena, a belt running across the north, originated by Hercynian folding, with summits reaching around 1300 m a.s.l., and essentially characterized by acid soils. The second is the Baetic range, a more recent limestone formation resulting from Alpine relief, which is located in the east. It is characterised by an extreme relief, with only 35 km from the coastline to the highest summit (3479 m a.s.l.). Thus, mild and harsh conditions prevail in very close proximity.

Location of the study area. An elevation gradient is shown
Between these two ranges, the Guadalquivir depression spans a triangular-shaped area running from the southwest to the northeast. Landscape consists in highly fragmented agricultural land with vestigial patches of original vegetation. The basin has loamy soils, low elevations and smooth terrains.
Target species
Stone pine is distributed across southern Europe and western Asia (López González 2007). Its natural area is difficult to define because the Romans contributed to spreading the trees (Costa Tenorio et al. 2005; Muñoz Álvarez 2010). Stone pine is well distributed in the west part of the study area. The largest pure stands are located in coastal areas of western Andalusia, but pines also occur in the eastern part of the Sierra Morena, mainly interspersed with holm oak (Quercus ilex L.) and cork oak (Quercus suber L.) stands. Stone pine prefers siliceous soils but can grow well on limestone. It is a thermophilous species, not exceeding elevations above 1000 m a.s.l. (Costa Tenorio et al. 2005). The other two species are more typical of the eastern end of the study area, with cluster pine usually growing at higher elevations than Aleppo pine.
Aleppo pine is a thermophilous species which adapts easily to diverse environments (particularly in the eastern xeric areas). The species mainly lives on limestone soils up to 1600 m a.s.l. in Andalusia (López González 2007). It is the most widespread pine species in the Mediterranean basin, where it grows under different climatic conditions (Barbéro et al. 1998; Richardson and Rundel 1998), and so holds significant interest as a forestry species. Main stands are found in North Africa and the Iberian Peninsula (Costa Tenorio et al. 2005).
Cluster pine is restricted to the western Mediterranean basin (López González 2007). Its main stands are in the Iberian Peninsula, where it has been widely planted. In the study area, it usually grows in altitudinal belts below stands of black pine (P. nigra Arnold) and scots pine (P. sylvestris L.) in the Baetic range; however, it can also be found along the Sierra Morena. It thrives on siliceous soils but can also grow on sandy limestone soils (López González 2007).
Dataset origin
The dataset used was developed from environmental data of REDIAM (Red de Información Ambiental de Andalucía) and AMAYA (Agencia de Medio Ambiente y Agua), both under the umbrella of the Andalusian Regional Government (Consejería de Medio Ambiente y Ordenación del Territorio, Junta de Andalucía). The study was carried out at 200 m resolution across four time periods: the reference period (1961–2000), early twenty-first century (2011–2040), middle twenty-first century (2041–2070) and late twenty-first century (2071–2100). The first period was used as a reference to predict potential areas and the others for future projections in the context of climate change. Each period was examined in three different scenarios (B1, A1b and A2). B1 describes a convergent world, undergoing rapid changes in economic structures toward a service and information economy, with a global population that peaks in mid-century. A1b retains the same global population as B1, but experiences less rapid changes in economic structures. It also assumes the rapid introduction of new and more efficient technologies and a balance across all resources. A2 depicts a very heterogeneous world, with high population growth, slow economic development and slow technological change. Based on the recommendations in the 4th Assessment Report of the Intergovernmental Panel on Climate Change (IPCC 2007), data were processed with CNCM3 GCM. Unfortunately, the 5th AR of the IPCC at 200 m resolution is still under construction for the study area.
Many approaches to the final model were carried out. With the aim of trying to avoid correlation and arrive at the best performance, a total of 11 explanatory variables were finally used. Temperature and rainfall are the key factors in global climate change. Temperature variables (MAT, MCT and MWT), as well as annual rainfall (AR), played an important role in demarking the suitability area (for abbreviations see the footnote of Table 1). Other additional variables had to be chosen according to the species or the study area. CI was calculated by subtracting the mean temperatures of the warmest (July) and coldest months (January). This parameter becomes higher the farther inland one travels, and can thus distort the balance of oceanic and continental environments. Moreover, variables such as MCT, MWT, CI, OMB and AAI were chosen with extreme conditions in mind, since the target species were known to easily adapt to diverse climatic conditions (Del Campo et al. 2014). OMB and AAI measure the humidity and aridity in the study area. The latter is especially important for the most arid zone located in the southeast of the study area. WB and PPA were also included because they are usually the subjects of silvicultural management. Water availability to plants is commonly evaluated from WB, which has previously been found to be a performing variable in modelling distribution of tree species (Zhang et al. 2002; Piedallu et al. 2013). Primary production availability is an important variable for species of economic significance, such as pines and other conifers (Acker et al. 2002; Baishya and Barik 2011; Campoe et al. 2013). It estimates the photosynthetic period from the photoperiod, temperature (over 7.5 °C) and positive water balance. The remaining variables (ETo and SR) were included in the ENMs because of their high degree of significance to the distribution of the target pine species. ETo measures the rate of evapotranspiration from a hypothetical crop of green grass of uniform height, actively growing, well-watered and completely shading the ground. Finally, SR measures the amount of solar energy from the sun.
Compilation of the dataset
The dataset was compiled using the software ArcGIS v. 10 (ESRI, 2010). Unsuitable areas for hosting woody tree species (marshes and rice crops, mainly) were excluded. Explanatory variables were recorded in raster files of 200 m resolution and pine occurrences as vector polygons (1:10,000 scale). Additional information was retrieved from three different projects: VEGE10 (detailed digital vegetation maps at the 1:10,000 scale), MUCVA (Map of Uses and Plant Covers of Andalusia at the 1:25,000 scale) and IFN3 (3rd National Forest Inventory). Natural and non-natural stands of the species were considered for this study. The occurrence data for stone pine, Aleppo pine and cluster pine was 42,128, 42,026 and 24,070 polygons respectively. After the conversion the result was 93,849, 101,899 and 47,131 points. The number of used absence points was 129,998. In total, the whole area comprised a grid of about 2 million points.
Development of the model
The database was processed by using the statistical software SPSS v.20 (IBM, 2011). Data for the three pine species were subjected to MLR analysis, using the main effects method, which is insensitive to small changes and provides accurate predictions (Augustin et al. 2001). The default options (PIN = 0.05, POUT = 0.1) were used. All explanatory variables were assumed to be continuous and hence used as “covariates”. The output of the model was the β value of each explanatory variable. The sign of β indicates whether a given variable supports (+) or avoids (−) the presence of the species concerned, and its absolute value is a measure of importance (the closer to 0, the less important the variable). Using β values in the multinomial logistic formula gave the probability for each point on the grid, which ranged from 0 (no probability of presence) to 1 (maximum probability of presence). Finally, probability values were used to construct a raster file by using GIS in order to develop a suitability distribution map at 200 m resolution.
Calibration and validation of the model
Nagelkerke’s R 2 (Nagelkerke 1991) was the selected method to assess the calibration of the model. For validation Akaike Information Criterion (AIC; Akaike 1973) and the Bayesian Information Criterion (BIC; Schwarz 1978) were used.
Results
Table 1 shows the mean and standard deviation, and the minimum and maximum values, for each individual species in the study area over the reference period. Based on temperature-related variables (MAT, MCT and MWT), stone pine was the most thermophilous species, followed by Aleppo pine and cluster pine. The values for stone pine were higher than the mean for the study area, while those for Aleppo pine and stone pine were lower. AR exhibited large differences between minimum and maximum values. Cluster pine was the species with the greatest mean, whereas Aleppo pine had the smallest mean and minimum values. CI mean values were quite similar across the three species, but somewhat lower for stone pine. The OMB values for stone pine and Aleppo pine were very similar to the mean for the study area. The highest OMB mean value was for cluster pine and the lowest for Aleppo pine. Obviously, the highest mean value of AAI was for Aleppo pine. More interestingly, its maximum value was much greater than those for stone pine and cluster pine. Aleppo pine was the species able to grow in the harshest conditions, thus exhibiting the smallest PPA. On the other hand, stone pine and cluster pine clearly exceeded the mean for the study area. WB in cluster pine was twice that of the other two species. By contrast, ETo and SR were similar for the three species and the study area, with stone pine slightly above and the other two species below the mean of the area.
Table 2 compares various future scenarios and periods. An increase in temperature-related variables and a decrease in annual rainfall, the two main factors influencing climate change predictions, can be anticipated.
The statistical results of the model are shown in Table 3. Overall, β was very close to 0, OMB excepted. Four out of the eleven explanatory variables had the same sign in β; specifically, MCT and OMB were both negative, whereas PPA and ETo were positive.
Electronic Supplementary Material shows the predicted probability of occurrence as a percentage, estimated by using a threshold of 0.5. The results reveal some trends for each species in different scenarios and periods of the twenty-first century. Stone pine extends its potential distribution until the late twenty-first century. In contrast, Aleppo pine and cluster pine suffer a significant reduction.
Nagelkerke’s R2 score was 0.8. AIC and BIC had lower values for the full model, 483,737.631 and 484,127.475 respectively (“final” in the model fitting information table of output data) than for the null model, 992,108.052 and 992,140.539 respectively (“intercept only”).
Figure 2 shows the current presence data and potential distribution of each species in the reference period. The potential distribution of suitable areas was wider for Aleppo pine and, especially, stone pine. Cluster pine exhibited less salient predictions relative to its current distribution. Based on these, the most suitable areas for stone pine are mainly in the Guadalquivir depression, running west to northeast. This species is not expected to reach higher elevations in the eastern part or in the northernmost sites in the Sierra Morena. In contrast, Aleppo pine prevails on the eastern end of the study area, matching the Baetic range, and is found largely at low elevations. Interestingly, there is a suitable patch in the centre of the Sierra Morena for which no current occurrence data are available. Cluster pine also has potential areas in the eastern part of the study area, but has a very important stand in the south-central part.

Current presence data (orange) and potential distribution in the reference period (green) of a stone pine, b Aleppo pine and c cluster pine. A suitability gradient from poor (light green) to the most suitable areas (dark green) is shown. White denotes the absence of suitability. (Color figure online)
In response to climate change, the potential distribution area for stone pine is predicted to increase over the course of the future periods studied (Fig. 3). The maps for scenarios B1 and A2 in the early twenty-first century are very similar, with A1b having the largest potential area. This trend is echoed by the predictions for the middle twenty-first century. On the other hand, A2 is predicted to be the best scenario for stone pine in the late twenty-first century. A2 and A1b exhibit a new potential patch in the east of the study area (viz. 450 m a.s.l. in the valley of Almanzora river). In contrast, suitable areas for Aleppo pine and cluster pine are predicted to contract (Figs. 4, 5). An upward migration in both pine species is expected at a similar rate in the three scenarios of the early twenty-first century. In this period, A2 is better for Aleppo pine, whereas all scenarios are similarly favourable for cluster pine. In the middle twenty-first century, the treeline is expected to rise again in both pine species. In this period, B1 is predicted to lead to lesser migration and a higher potential. The least suitable areas are those for cluster pine in A2, A1b and A2 being equally favourable for Aleppo pine. Finally, the upward migration will continue in the late twenty-first century (especially in B1 and, less markedly, in A2). The most drastic reduction in potential area for both species will occur in A2.

Potential distribution of stone pine in three different scenarios during the twenty-first century. For details, see caption to Fig. 2

Potential distribution of Aleppo pine in three different scenarios during the twenty-first century. For details, see caption to Fig. 2

Potential distribution of cluster pine in three different scenarios during the twenty-first century. For details, see caption to Fig. 2
Discussion
The low resolution of ENMs has often reduced the accuracy of their predictions (Ruiz-Labourdette et al. 2012). Studies at very high resolutions have been conducted in recent years on some forest species (Hidalgo et al. 2008; López-Tirado and Hidalgo 2014). This research was carried out at a resolution of 200 m, which led to accurate species distribution and explanatory variables to be identified. MLR involves zoning among the target species. Thus, the analysis of forest trees by ENMs is particularly useful when biotic factors are considered (Coudun et al. 2006; González-Salazar et al. 2013; Vessella et al. 2015). Predicting suitable areas for growth allows afforestation/reforestation programs to be highly reliably designed. The goodness of the model developed here is supported by its Nagelkerke’s R 2 value (viz. 0.4, indicating good calibration; Bässler et al. 2010). Also, the AIC and BIC values are indicative of a high goodness of fit (Burnham and Anderson 2004).
The mean, minimum and maximum values of the temperature-related variables (MAT, MCT and MWT) can be discussed jointly. Stone pine grows in coastal areas and low altitudinal ranges such as the Sierra Morena. Its values (the highest) correlated inversely with elevation by effect of its lapse rate (Jacobson 2005; Ahrens 2006). The β values for MAT and MWT confirm its tendency to seek increased temperatures. In contrast, the β values for MAT in cluster pine and Aleppo pine were both negative. Both species occur mainly in the Baetic range at higher altitudes than the sites occupied by stone pine, cluster pine generally occurring at higher altitudes than Aleppo pine. The negative β value of MCT for all species indicates avoidance of the lowest temperatures. The extreme values obtained for AR can be accounted for by the complex relief and rainfall patterns which are responsible for producing within close proximity sub-desert areas and the wettest area in the Iberian Peninsula (De Castro et al. 2005). Aleppo pine exhibited the smallest minimum value of AR, which confirms its importance in semi-arid zones (Néeman and Trabaud 2000; Derak and Cortina 2014). In contrast, cluster pine receives higher AR due to Föhn’s effect (Strahler and Strahler 1989), while stone pine remains at an intermediate level. Interestingly, it had the lowest standard deviation values, indicating a more regular regime of rainfall from the western Atlantic fronts. At the same time, the β value for AR was negative, which suggests that stone pine could grow in areas with lower AR. Meanwhile, Aleppo pine had a positive β value, indicating preference for areas with increased AR—this species had the lowest mean AR. The decreasing trend in annual rainfall in recent decades has led to a decline in P. halepensis subsp. brutia (Ten.) Holmboe in the eastern Mediterranean basin (Sarris et al. 2007, 2011). Cluster pine was theoretically insensitive to AR. CI suggests that the oceanic environment is suitable for stone pine, a finding which is consistent with the location of the majority of the pure stands on the west coast and the negative β value. CI was slightly greater for Aleppo pine and cluster pine, due to the prevalence of inland stands—hence the positive sign in β. The β value for OMB was farther from 0 than that of the other variables, and thus enjoys slightly greater weight in the model. Its negative sign indicates avoidance of wetter ombrotypes even though cluster pine had the value closest to zero (i.e. it is the species less markedly avoiding wet areas). In fact, this species had the greatest mean value in CI for the same reason as AR (i.e. its growing at higher elevations and in mountainous areas). In contrast, the AAI score for Aleppo pine confirms the harsh conditions where it usually grows—and led to a positive β value for this variable. This trait suggests that populations of this species should be less attractive for economic purposes. Aleppo pine had the lowest PPA and WB values, in fact, these were the closest to zero of all. Only WB for Aleppo pine was negative, the explanation of which is similar to that for AAI. Stone pine and cluster pine had negative AAI values because they grow in wetter areas than Aleppo pine. ETo values exhibited no substantial differences among the species, all three having a positive β value. β was also positive for SR in stone pine. Flat and low altitude sites are generally typical of the distribution of this species in the western area. Stone pine is a shade intolerant species (López González 2007), which resulted in the highest SR mean value, and hence in the highest ETo value. On the other hand, Aleppo pine and cluster pine had a negative β value in SR because the main stands are located on mountainous areas in the Baetic range.
MAT, MCT and MWT were predicted to increase over the twenty-first century (see Table 2). CI, AAI, ETo and SR exhibited a similar trend, whereas AR, OMB, PPA and WB changed in the opposite direction. With respect to the three scenarios, B1 exhibited the most extreme values in the early period. On the other hand, A1b had the highest temperatures in the middle period, although OMB, PPA, WB and AAI were highest in A2. In fact, A2 was the harshest scenario in the late twenty-first century, while B1 was the mildest. Overall, the model predicted a mean increase of 3.08 °C and a decrease of 97.84 mm in AR, with the other variables changing accordingly. Worthy of special note here is the drastic decrease predicted in PPA (591.99) and WB (495.87), i.e. about 31 and 57 %, respectively.
The results show that the largest area is occupied by stone pine, followed by Aleppo pine and cluster pine. Specifically, stone pine is predicted to expand during the twenty-first century, whereas Aleppo pine and cluster pine might exhibit the opposite trend. In the harshest scenario (viz. A2 in the late twenty-first century), there would be no suitable areas exceeding the 0.5 probability threshold for the latter species. A comparison of extreme scenarios suggests that Aleppo pine might undergo a steady reduction in potentiality over a quarter of the area it presently occupies. Conversely, stone pine might increase by about 17 % (see Electronic Supplementary Material).
The thermophilous nature of Aleppo pine will probably hinder migration to higher altitude sites (López González 2007). The centre of the Sierra Morena, which we predicted to be a suitable region for Aleppo pine, would be lost in the worst case scenario for the last period (A2). Stone pine can be reliably expected to occupy suitable sites in this scenario—the best for this species according to Fig. 3. This prediction is consistent with Bede-Fazekas et al. (2014) which stated that most of the Spanish distribution of stone pine will remain viable in the middle twenty-first century. According to these authors, cluster pine populations at the lowest altitude sites could be lost due to migration to higher elevations in the south of the Iberian Peninsula over the period 2011–2040. Our high resolution model enables the identification of a trend in cluster pine towards higher elevations until the end of the twenty-first century. Similar poleward and upward migration trends have also been projected by other authors (Bertrand et al. 2012; Lenoir et al. 2008; Parmesan and Yohen 2003; Rabasa et al. 2013).
Spanish territory is dominated by a small number of species, including pines (García del Barrio et al. 2013). Overall, stone pine would be the most suitable pine species in the western and northeastern zones of the study area. Based on the goodness of fit provided by the model, stone pine could be a key species for afforesting the Guadalquivir depression in restoration programs. These theoretical populations could shelter afforested specimens of holm oak, which is the main natural species (Rivas-Martínez 1987; Valle 2004). Consequently, the formation of natural patches and corridors might reduce fragmentation in this large deforested territory.
On the other hand, Aleppo pine would be the most suitable species in the eastern end. Unfortunately, this is one of the most desertification-prone zones (Fons-Esteve and Páramo 2003). Because stone pine has a good recruitment capacity and adapts well to stressful environments (Manso et al. 2014), it should be considered in future reforestation programs in some of these areas. Cluster pine is predicted to undergo a very marked decrease in suitability. Climate/growth models have predicted a strong decline in basal area increment during the twenty-first century in the northwest of the Iberian Peninsula (González-Muñoz et al. 2014). Higher temperatures in spring and summer could increase water stress and have an adverse impact on cluster pine (Nabais et al. 2014), which might migrate to the highest elevations in response.
In conclusion, the ENM method provides very good fit in predicting and forecasting the evolution of the target Mediterranean pines species. The model is highly accurate and the ensuing maps provide a powerful tool for designing reforestation programs in the context of global climate change and afforestation (Gastón et al. 2014). These results can be considered especially relevant to anthropogenic areas with massive landscape fragmentation. Further studies could be carried out with multiple algorithms addressing the whole distribution so as to achieve a deeper understanding of global changes affecting the species.
References
Acker SA, Halpern CB, Harmon ME, Dyrness CT (2002) Trends in bole biomass accumulation, net primary production and tree mortality in Pseudotsuga menziesii forests of contrasting age. Tree Physiol 22:213–217
Ahrens CD (2006) Meteorology today, 8th edn. Brooks/Cole Publishing, California
Akaike H (1973) Information theory as an extension of the maximum likelihood principle. Second international symposium on information theory. Akademiai Kiado, Budapest, pp 267–281
Alba-Sánchez F, López-Sáez JA, Benito-de Pando B, Linares JC, Nieto-Lugilde D, López-Merino L (2010) Past and present potential distribution of the Iberian Abies species: a phytogeographic approach using fossil pollen data and species distribution models. Divers Distrib 16:214–228
Araùjo MB, Guisan A (2006) Five (or so) challenges for species distribution modeling. J Biogeogr 33:1677–1688
Augustin NH, Cummins RP, French DD (2001) Exploring spatial vegetation dynamics using logistic regression and a multinomial logit model. J Appl Ecol 38(5):991–1006
Baishya R, Barik SK (2011) Estimation of tree biomass, carbon pool and net primary production of an old-growth Pinus kesiya Royle ex. Gordon forest in north-eastern India. Ann For Sci 68:727–736. doi:10.1007/s13595-011-0089-8
Barbéro M, Loisel R, Quézel P, Richardson MD, Romane F (1998) Pines of the Mediterranean basin. In: Richardson DM (ed) Ecology and biogeography of Pinus. Cambridge University Press, Cambridge, pp 153–170
Bässler C, Müller J, Hothorn T, Kneib T, Badeck F, Dziock F (2010) Estimation of the extinction risk for high-montane species as a consequence of global warming and assessment of their suitability as cross-taxon indicators. Ecol Indic 10:341–352. doi:10.1016/j.ecolind.2009.06.014
Bede-Fazekas A, Horvath L, Kocsis M (2014) Impact of climate change on the potential distribution of Mediterranean pines. Idojaras 118:41–52
Bertrand R, Perez V, Gegout JC (2012) Disregarding the edaphic dimension in species distribution models leads to the omission of crucial spatial information under climate change: the case of Quercus pubescens in France. Glob Change Biol 18:2648–2660
Burnham KP, Anderson DR (2004) Multimodel inference—understanding AIC and BIC in model selection. Sociol Meth Res 33:261–304. doi:10.1177/0049124104268644
Cabezudo Artero B, Pérez Latorre AV (2004) Las comunidades vegetales. In: Herrera CM (ed) El monte mediterráneo en Andalucía. Consejería de Medio Ambiente, Junta de Andalucía, Sevilla, pp 29–45
Campoe OC, Stape JL, Albaugh TJ, Allen HL, Fox TR, Rubilar R, Binkley D (2013) Fertilization and irrigation effects on tree level aboveground net primary production, light interception and light use efficiency in a loblolly pine plantation. For Ecol Manage 288:43–48. doi:10.1016/j.foreco.2012.05.026
Carrión JS, Díez MJ (2004) Origen y evolución de la vegetación mediterránea en Andalucía a través del registro fósil. In: Herrera CM (ed) El monte mediterráneo en Andalucía. Consejería de Medio Ambiente, Junta de Andalucía, Sevilla, pp 21–27
Carrion JS, Navarro C, Navarro J, Munuera M (2000) The distribution of cluster pine (Pinus pinaster) in Spain as derived from palaeoecological data: relationships with phytosociological classification. Holocene 10:243–252. doi:10.1191/095968300676937462
Ceballos L, Ruiz de la Torre J (1979) Árboles y arbustos, 1st edn. Escuela Técnica Superior de Ingenieros de Montes, Madrid
Costa Tenorio M, Morla Juaristi C, Sainz Ollero H (eds) (2005) Los bosques ibéricos. Una interpretación geobotánica, Editorial Planeta, Barcelona
Coudun C, Gégout JC, Piedallu C, Rameau JC (2006) Soil nutritional factors improve models of plant species distribution: an illustration with Acer campestre (L.) in France. J Biogeogr 33:1750–1763
De Castro M, Martín-Vide J, Alonso S (2005) El clima de España: pasado, presente y escenarios de clima para el siglo XXI. In: Moreno Rodríguez JM (ed) Evaluación preliminar de los impactos en España por efecto del cambio climático, Ministerio de Medio Ambiente y Universidad de Castilla–La Mancha, pp 1–64
Del Campo AD, Fernandes TJG, Molina AJ (2014) Hydrology-oriented (adaptive) silviculture in a semiarid pine plantation: How much can be modified the water cycle through forest management? Eur J For Res 133:879–894. doi:10.1007/s10342-014-0805-7
Derak M, Cortina J (2014) Multi-criteria participative evaluation of Pinus halepensis plantations in a semiarid area of southeast Spain. Ecol Indic 43:56–68. doi:10.1016/j.ecolind.2014.02.017
Elith J, Graham CH, Anderson RP, Dudík M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton JM, Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberon J, Williams S, Wisz MS, Zimmermann NE (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
Felicísimo AM, Muñoz J, Mateo RG, Villalba CJ (2012) Vulnerabilidad de la flora y vegetación españolas ante el cambio climático. Ecosistemas 21(3):1–6
Finley AO, Banerjee S, McRoberts RE (2009) Hierarchical spatial models for predicting tree species assemblages across large domains. Ann Appl Stat 3:1052–1079. doi:10.1214/09-aoas250
Fons-Esteve J, Páramo F (2003) Mapping sensitivity to desertification (DISMED). European Environmental Agency, European Topic Center, Terrestrial Environment, Internal Report
García del Barrio JM, Aunon F, Sanchez de Ron D, Alia R (2013) Assessing regional species pools for restoration programs in Spain. New For 44:559–576. doi:10.1007/s11056-013-9363-y
García-Amorena I, Manzaneque F, Gómez Rubiales JM, Granja HM, Soares de Carvalho G, Morla C (2007) The late quaternary coastal forests of western Iberia: a study of their macroremains. Palaeogeogr Palaeoclimatol Palaeoecol 254:448–461
García-Ruiz JM, López-Moreno JI, Vicente-Serrano SM, Lasanta T, Beguería S (2011) Mediterranean water resources in a global change scenario. Earth Sci Rev 105:121–139
García-Valdés R, Zavala MA, Araújo MB, Purves DW (2013) Chasing a moving target: projecting climate change-induced shifts in non-equilibrial tree species distributions. J Ecol 101:441–453
Gastón A, García-Viñas JI, Bravo-Fernández AJ, López-Leiva C, Oliet JA, Roig S, Serrada R (2014) Species distribution models applied to plant species selection in forest restoration: are model predictions comparable to expert opinion? New For 45:641–653
Giorgi F, Lionello P (2008) Climate change projections for the Mediterranean region. Glob Planet Change 63:90–104
Gómez A, Aguiriano E, Alía R, Bueno MA (2002) Análisis de los recursos genéticos de Pinus pinea L. en España mediante microsatélites del cloroplasto. Invest Agr Sist Recur For 11:145–154
González-Muñoz N, Linares JC, Castro-Díez P, Sass-Klaassen U (2014) Predicting climate change impacts on native and invasive tree species using radial growth and twenty-first century climate scenarios. Eur J For Res 133:1073–1086. doi:10.1007/s10342-014-0823-5
González-Salazar C, Stephens CR, Marquet PA (2013) Comparing the relative contributions of biotic and abiotic factors as mediators of species’ distributions. Ecol Model 248:57–70
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8:993–1009
Hidalgo PJ, Marin JM, Quijada J, Moreira JM (2008) A spatial distribution model of cork oak (Quercus suber) in southwestern Spain: a suitable tool for reforestation. For Ecol Manage 255:25–34. doi:10.1016/j.foreco.2007.07.012
Ibáñez I, Katz DSW, Peltier D, Wolf SM, Barrie BTC (2014) Assessing the integrated effects of landscape fragmentation on plants and plant communities: the challenge of multiprocess–multiresponse dynamics. J Ecol 102:882–895. doi:10.1111/1365-2745.12223
IPCC (2007) Summary for policymakers climate change 2007: the physical science basis. In: Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change, Cambridge University Press, Cambridge
Jacobs DF, Oliet JA, Aronson J, Bolte A, Bullock JM, Donoso PJ, Landhäusser SM, Madsen P, Peng S, Rey-Benayas JM, Weber JC (2015) Restoring forests: what constitutes success in the twenty-first century? New For 46:601–614. doi:10.1007/s11056-015-9513-5
Jacobson MZ (2005) Fundamentals of atmospheric modeling, 2nd edn. Cambridge University Press, Cambridge
Lenoir J, Gégout JC, Marquet PA, de Ruffray P, Brisse H (2008) A significant upward shift in plant species optimum elevation during the 20th century. Science 320:1768
López González GA (2007) Guía de los árboles y arbustos de la Península Ibérica y Baleares, 3rd edn. Ediciones Mundi-Prensa, Madrid
López-Sáez JA, López-Merino L, Alba-Sánchez F, Pérez-Díaz S, Abel-Schaad D, Carrión JS (2010) Late Holocene ecological history of Pinus pinaster forests in the Sierra de Gredos of central Spain. Plant Ecol 206:195–209. doi:10.1007/s11258-009-9634-z
López-Tirado J, Hidalgo PJ (2014) A high resolution predictive model for relict trees in the Mediterranean-mountain forests (Pinus sylvestris L., P. nigra Arnold and Abies pinsapo Boiss.) from the south of Spain: a reliable management tool for reforestation. For Ecol Manage 330:105–114. doi:10.1016/j.foreco.2014.07.009
Manso R, Pukkala T, Pardos M, Miina J, Calama R (2014) Modelling Pinus pinea forest management to attain natural regeneration under present and future climatic scenarios. Can J Forest Res Rev Can Rech For 44:250–262. doi:10.1139/cjfr-2013-0179
Márquez AL, Real R, Olivero J, Estrada A (2011) Combining climate with other influential factors for modelling the impact of climate change on species distribution. Clim Change 108:135–157. doi:10.1007/s10584-010-0010-8
Martínez F, Montero G (2004) The Pinus pinea woodlands along the coast of Southwestern Spain: data for a new geobotanical interpretation. Plant Ecol 175:1–18
Martínez F, Montero G, Ruiz-Peinado R, Cañellas I, Candela JA (2004) Geobotánica e historia de los Pinares. In: El pino piñonero (Pinus pinea L.) en Andalucía. Ecología, distribución y selvicultura. Junta de Andalucía, Consejería de Medio Ambiente Sevilla, pp 49–112
Mateo RG, Felicisimo AM, Munoz J (2011) Species distributions models: a synthetic revision. Rev Chil Hist Nat 84:217–240
Mellert KH, Fensterer V, Kuechenhoff H, Reger B, Koelling C, Klemmt HJ, Ewald J (2011) Hypothesis-driven species distribution models for tree species in the Bavarian Alps. J Veg Sci 22:635–646. doi:10.1111/j.1654-1103.2011.01274.x
Montero M (1997) Breve descripción del proceso repoblador en España (1940–1995). Legno Celulosa Carta 4:35–42
Muñoz Álvarez JM (ed) (2010) Vegetación de la Reserva de la Biosfera y de los Espacios Naturales de Sierra Morena, Consejería de Medio Ambiente, Junta de Andalucía, Córdoba
Nabais C, Campelo F, Vieira J, Cherubini P (2014) Climatic signals of tree-ring width and intra-annual density fluctuations in Pinus pinaster and Pinus pinea along a latitudinal gradient in Portugal. Forestry 87:598–605. doi:10.1093/forestry/cpu021
Nagelkerke NJD (1991) A note on a general definition of the coefficient of determination. Biometrika 78:691–692
Néeman G, Trabaud L (eds) (2000) Ecology, biogeography and management of Pinus halepensis and Pinus brutia forest ecosystems in the Mediterranean Basin. Backhuys Publication, Leiden
Olden JD, Jackson DA, Peres-Neto PP (2002) Predictive models of fish species distributions: a note on proper validation and chance predictions. Trans Am Fish Soc 131:329–336
Parmesan C, Yohen G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42
Piedallu C, Gegout JC, Perez V, Lebourgeois F (2013) Soil water balance performs better than climatic water variables in tree species distribution modelling. Global Ecol Biogeogr 22:470–482. doi:10.1111/geb.12012
Rabasa SG, Granda E, Benavides R, Kunstler G, Espelta JM, Ogaya R, Peñuelas J, Scherer-Lorenzen M, Gil W, Grodzki W, Ambrozy S, Bergh J, Hódar JA, Zamora R, Valladares F (2013) Disparity in elevational shifts of European trees in response to recent climate warming. Glob Change Biol 19:2490–2499
Rejmánek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77:1655–1661
Richardson DM, Rundel PW (1998) Ecology and biogeography of Pinus: an introduction. In: Richardson DM (ed) Ecology and biogeography of Pinus. Cambridge University Press, Cambridge
Rivas-Martínez S (1987) Memoria del mapa de series de vegetación de España. Ministerio de Agricultura, Pesca y Alimentación, ICONA, Madrid
Rubiales JM, García-Amorena I, García Álvarez S, Morla C (2009) Anthracological evidence suggests naturalness of Pinus pinaster in inland southwestern Iberia. Plant Ecol 200:155–160. doi:10.1007/s11258-008-9439-5
Ruiz-Labourdette D, Nogues-Bravo D, Ollero HS, Schmitz MF, Pineda FD (2012) Forest composition in Mediterranean mountains is projected to shift along the entire elevational gradient under climate change. J Biogeogr 39:162–176. doi:10.1111/j.1365-2699.2011.02592.x
Sánchez-Salguero R, Navarro-Cerrillo RM, Camarero JJ, Fernández-Cancio A (2010) Drought-induced growth decline of Aleppo and maritime pine forests in south-eastern Spain. For Syst 19(3):458–469
Sánchez-Salguero R, Navarro-Cerrillo RM, Camarero JJ, Fernández-Cancio A (2012) Selective drought-induced decline of pine species in southeastern Spain. Clim Change 113:767–785
Sarris D, Christodoulakis D, Koerner C (2007) Recent decline in precipitation and tree growth in the eastern Mediterranean. Glob Change Biol 13:1187–1200
Sarris D, Christodoulakis D, Koerner C (2011) Impact of recent climatic change on growth of low elevation eastern Mediterranean forest trees. Clim Change 106:203–223. doi:10.1007/s10584-010-9901-y
Schwarz G (1978) Estimating the dimension of a model. Ann Stat 6:461–464
Soto A, Robledo-Arnuncio JJ, Gonzalez-Martinez SC, Smouse PE, Alia R (2010) Climatic niche and neutral genetic diversity of the six Iberian pine species: a retrospective and prospective view. Mol Ecol 19:1396–1409. doi:10.1111/j.1365-294X.2010.04571.x
Stevenson AC (1985) Studies in the Vegetational History of S. W. Spain. I. Modern pollen rain in the Doñana National Park, Huelva. J Biogeogr 12(3):243–268
Strahler AN, Strahler AH (1989) Geografía Física. 3rd end, Editorial Omega S.A. Barcelona, p 539
Sumarga E (2011) A comparison of logistic regression, geostatistics and Maxent for distribution modeling of a forest endemic; a pilot study on lobel’s maple at Mt. Pizzalto, Italy. Ph.D. Dissertation, University of Twente, The Netherlands
Valle F (ed) (2004) Modelos de restauración forestal. Consejería de Medio Ambiente, Junta de Andalucía
Vessella F, Schirone B (2013) Predicting potential distribution of Quercus suber in Italy based on ecological niche models: conservation insights and reforestation involvements. For Ecol Manage 304:150–161
Vessella F, Simeone MC, Schirone B (2015) Quercus suber range dynamics by ecological niche modelling: from the Last Interglacial to present time. Quat Sci Rev 119:85–93
Zavala MA, Zea K (2004) Mechanisms maintaining biodiversity in Mediterranean pine-oak forests: insights from a spatial simulation model. Plant Ecol 171:197–207
Zhang L, Walker GR, Dawes WR (2002) Water balance modelling: concepts and applications. In: McVicar TR, Rui L, Walker J, Fitzpatrick RW, Liu C (eds) Regional water and soil assessment for managing sustainable agriculture in China and Australia, ACIAR Monograph 84, Canberra, pp 31–47
Acknowledgments
The authors are grateful to the Council of Economy, Innovation, Science and Employment of the Andalusian Regional Government for supporting this study in the framework of the project ‘‘Modelo espacial de distribución de las quercíneas y otras formaciones forestales de Andalucía: una herramienta para la gestión y la conservación del patrimonio natural” (Code P10-RNM-6013). This is the contribution no. 115 from the CEIMAR Journal Series.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
Additional information on comparative areas between the reference period and each scenario is shown for stone pine. Only cells with a probability over 0.5 (as percentage next to each map) were taken into account. Green corresponds to area gained, purple indicates lost areas and blue shows stable areas. White denotes the absence of suitability for both period and scenario (TIFF 2767 kb)
ESM 2
Additional information on comparative areas between the reference period and each scenario is shown for Aleppo pine. For details, see caption to ESM 1 (TIFF 3540 kb)
ESM 3
Additional information on comparative areas between the reference period and each scenario is shown for cluster pine. For details, see caption to ESM 1 (TIFF 1001 kb)
Rights and permissions
About this article

Cite this article
López-Tirado, J., Hidalgo, P.J. Ecological niche modelling of three Mediterranean pine species in the south of Spain: a tool for afforestation/reforestation programs in the twenty-first century. New Forests 47, 411–429 (2016). https://doi.org/10.1007/s11056-015-9523-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-015-9523-3