
Abstract
Most research on carbon content of trees has focused on temperate species, with less information existing for tropical trees and very little for tropical plantations. This study investigated factors affecting the carbon content of nineteen tropical plantation tree species of ages seven to twelve and compared carbon content of Khaya species from two ecozones in Ghana. For all sample trees, volume of the main stem, wood density, wood carbon (C) concentration and C content were determined. Estimated stem volume for the 12-year-old trees varied widely among species, from 0.01 to 1.04 m3, with main stem C content ranging from 3 to 205 kg. Wood density among species varied from 0.27 to 0.76 g cm−3, with faster growing species exhibiting lower density. Significant differences in wood density also occurred with position along the main stem. Carbon concentration also differed among tree species, ranging from 458 to 498 g kg−1. Differences among species in main stem C content largely reflected differences among species in estimated main stem volume, with values modified somewhat by wood density and C concentration. The use of species-specific wood density values was more important for ensuring accurate conversion of estimated stem volumes to C content than was the use of species-specific C concentrations. Significant differences in wood density did exist between Khaya species from the wet and moist semi-deciduous ecozones, suggesting climatic and site factors may also need to be considered. Wood densities for these plantation grown trees were lower than literature values reported for the same species in natural forests, suggesting that the application of data derived from natural forests could result in overestimation of the biomass and C content of trees of the same species grown in plantations.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Growing concerns about the climatic impacts of increased concentrations of atmospheric greenhouse gases have stimulated discussions about the importance and potential of using forests for carbon sequestration. Due to anthropogenic emissions, the concentration of the major greenhouse gas carbon dioxide (CO2) has increased from around 280 μL L−1 in the late 1800s to the more than 390 μL L−1 in 2013 (Schneider 1990; NOAA 2013). Mean global temperatures increased by 0.74 °C over the same time period (Solomon et al. 2007), and regional temperatures in West Africa may increase by an additional 2–5 °C over the next century as atmospheric CO2 concentration continues to rise (Christensen et al. 2007).
To offset a portion of the escalating CO2 concentrations, afforestation and reforestation systems have been encouraged as means to sequester CO2 in biomass. This concept was formally endorsed by the Kyoto Protocol, which allowed for the opportunity to offset CO2 emissions through collaboration between developed and developing nations in reforestation or afforestation projects (Desanker 2005; ITTO 2006; World Bank 2009). The purposes for promoting plantation development in West African nations, such as Ghana, include restoring degraded forests, providing raw materials for industry and bioenergy, providing non-timber goods for local populations, and potentially creating extra income from carbon credits as a means of value addition (FC 2006; Sandker et al. 2010; Kalame et al. 2011). Under the United Nation’s REDD + program (Reducing Emissions from Deforestation and Forest Degradation, with the “+” adding conservation, sustainable forest management and enhancement of forest carbon stocks), planting of trees for reforestation or canopy shade cover can be eligible for carbon credits. Ghana is among the REDD partner countries, and thus needs to be able to provide authenticated estimates of the nation’s forest carbon stocks (Gibbs et al. 2007).
Quantifying carbon stocks in forests requires accurate estimation of aboveground biomass (Brown et al. 1989; Ketterings et al. 2001; Chave et al. 2005), but many factors can influence tree and forest biomass and C content, including tree species, climate, topography, soil fertility, and water supply (Fearnside 1997; Luizão et al. 2004; Sicard et al. 2006; Slik et al. 2008). Wood density can serve to integrate the effects of these factors and thus is often viewed as a critical parameter needed for deriving biomass estimates from volumes determined using forest inventory data (Brown et al. 1989; Fearnside 1997; Preece et al. 2012). Note, however, that it has been suggested that species-specific wood density may not be needed for estimating biomass of small diameter stems in tropical dry forest (Chaturvedi and Raghumbanshi 2013).
Along the main stem of a tree, wood density varies from the base to the top of the stem, and radially from pith to bark (Henry et al. 2010). Typically, wood density decreases from the stump to half of the total height of the tree, and may increase afterwards towards the top (Espinoza 2004). Wood density also varies with species, age and geographical location in natural tropical forests (Fearnside 1997; Suzuki 1999; Slik et al. 2008; Henry et al. 2010). However, little information exists on wood density for trees growing in plantations of the species recommended for use in Ghana and other sub-Saharan African countries. Available information on the volume, biomass and carbon content for tropical tree species is limited relative to the wide variety of species and regional environments that exist in the tropics, making estimation of the value of many species as carbon sinks difficult to predict. Available data has revealed significant differences among different tree species growing at various sites (Elias and Potvin 2003; Lamlom and Savidge 2006; Bert and Danjon 2006). In addition, many of the existing equations for estimating volume and biomass of tropical trees were derived from native forests (Chave et al. 2005; Henry et al. 2010), potentially limiting their applicability to plantations intended for afforestation or reforestation, which will for many years involve smaller trees grown in full sunlight. This research seeks to help fill the knowledge gaps regarding the carbon content of plantation tree species in the tropics by providing information on wood density, stem volume and biomass, and carbon concentration and content for those that are likely to be used in afforestation and reforestation efforts in the wet ecozone of Ghana and similar areas of West Africa. The study also compared the wood density of mahogany (Khaya) species planted in the moist semi-deciduous and wet evergreen forest ecozones of Ghana.
Specific objectives were to assess: (1) differences among species in volume growth and main stem C content; (2) differences among species and stem locations in wood density and C concentration and the potential errors that may result from using only lower-stem density data to estimate stem biomass; (3) the relative importance of knowing species-specific wood density and C concentration for accurately determining main stem C content; (4) if wood density varies with tree age; (5) if wood density and C content for Khaya species differ among ecozones; (6) if differences in growth rate, wood density or C content are related to a species guild classification; and (7) if wood density values for species growing in plantation are similar to those for the same species growing in a natural forest. We hypothesized that there will be significant differences in carbon content among trees species in the same ecozone and among Khaya planted in the moist semi-deciduous and wet evergreen ecozones, due primarily to differences in wood density. We also hypothesized that fast growing plantation trees will have lower wood density, and thus C content for a given volume, than the same species are known to have in natural forests.
Methods
Study areas
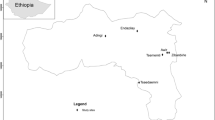
Two study areas were used, the first in the Oda-kotoamso Community Agroforestry Project (OCAP), and the second at Bobiri forest reserve (Fig. 1). Oda-kotoamso is located in the western region of Ghana, about 10 km from Asankraqwa, the district capital of the Wassa Amenfi political district. Geographically, Oda-kotoamso lies between latitude 5°18′ and 5°45′ N and longitude 2°10′ and 2°30′ W. This area falls within the hot humid tropical rainforest of the wet evergreen zone. There are two rainy seasons: a major season from April to July, and a minor season from September to November. Average annual rainfall ranges from 1,750 to 2,000 mm (Hall and Swaine 1981). Average monthly temperature ranges from approximately 24–27 °C at the nearby (about 70 km) Ghana Rubber Estate Plantation (Wauters et al. 2008). The landscape is characterised by undulating stretches of land with hills and flattened mountains. Elevation ranges from about 90 to 400 m above sea level. The soils are acidic with a pH of approximately 3–4 (Hall and Swaine 1981).

Location of the OCAP and Bobiri study areas in Ghana, West Africa
The plantation at OCAP was initiated in 1997 and has a total size of approximately 290 ha. To date, 20 native tropical and 3 exotic species have been successfully planted as either mixed or single species stands, with spacing ranging from 3 × 3 to 4 × 4 m. The plantation was developed and is owned by over eighty outgrower farmers with technical and financial support from Samartex Timber and Plywood company in Samreboi, Ghana.
The second site for the study was Bobiri (6°40′ N, 1°19′ W), about 35 km from Kumasi in the Ashanti region of Ghana. The Bobiri site falls within the moist semi-deciduous forest, which is drier than the wet evergreen forest zone. Average annual rainfall for the moist semi-deciduous forest ranges from 1,200 to 1,800 mm (Hall and Swaine 1981), variation in monthly temperature is similar to that at Oda-kotoamso, and the daily high temperature peaks at about 32 °C in March. Topography of the area is moderate, with elevation ranging from 150 to 600 m (Hall and Swaine 1981). The soils are slightly acidic with pH of approximately 5–6 (Hall and Swaine 1981). The Bobiri plantation consists of single species stands of Khaya ivorensis and Khaya grandifoliola on one-hectare plots.
The plantations at both sites originated from degraded forest lands which were covered largely by Chromolaena odorata (L.) R. King & H. Rob. and other vegetation. During preparation for planting, the vegetation was cut down by manual weeding and the debris was burnt. The next activity was pegging, which involved mapping the location for each seedling before planting one-year old seedlings, from local seed sources, at 3 m × 3 m spacing for 7- and 5-year-old plantations and 4 m × 4 m spacing for the 12-year-old plantations. At both sites, no other silvicultural treatments were performed in the plantations except weeding Chromolaena odorata and other competing vegetations three times per year. An inventory of trees was done in these plantations and unique numbers were marked on each tree. Selection of trees for this study was performed through random selection from the pool of unique numbers for each species.
Tree sampling
A total of sixty nine trees were randomly selected from the plantations for determination of stem volume, wood density and carbon concentration. For the OCAP wet evergreen forest, a total of 41 and 12 trees of ages twelve and seven, respectively, were sampled. Eighteen species of age twelve were sampled at OCAP: Aningeria robusta A.Chev., Antiaris toxicaria Lesch., Cedrela odorata L., Ceiba pentandra (L.) Gaertn., Entandrophragma angolense (Welw.) C.DC., Guarea thompsonii Sprague & Hutch., Heritiera utilis (Sprague) Sprague, Khaya ivorensis A.Chev., Lophira alata Banks ex P.Gaertn., Mammea africana Sabine, Milicia excelsa (Welw.) C.C.Berg, Pycnanthus angolensis (Welw.) Warb., Tectona grandis L.f., Terminalia ivorensis A.Chev., Terminalia superba Engl. & Diels, Tieghemella heckelii (A.Chev.) Roberty, Triplochiton scleroxylon K.Schum., and Turraeanthus africanus (Welw. ex C.DC.) Pellegr. Four species of age 7 were sampled: Antrocaryon micraster A.Chev. & Guill., Heritiera utilis, Khaya ivorensis, and Turraeanthus africanus. Two to three trees per species were sampled within an age class at a plantation. In addition, four 5-year-old trees from each of two species, Khaya ivorensis and Khaya grandifoliola C.DC., were selected from both the wet evergreen zone plantations at OCAP and the moist semi-deciduous forest zone at Bobiri.
Sample trees were cut down and diameter at breast height (dbh, at 1.3 m) and the length of the main stem (stump to first large canopy branch) were measured. The main stem was divided into three sections, and volumes of the base (stump to 1.3 m), middle (1.3 m to midpoint) and top (midpoint to top) portions were computed using Smalian’s model (Avery and Burkhart 2002):
where V is the estimated volume of a stem section in m3, B is the cross-sectional area (m2) at the large end of the stem section, b is the cross-sectional area (m2) at the small end of the section, and L is the length of the stem section in m. The values for b and B were calculated from the diameters of the two ends of the stem section, assuming the log had a circular cross section.
By dividing the main stem into several smaller-length sections for which we measured end diameters and length, we were able to use Smalian’s model to estimate volume for each section and then sum them to determine the total main stem volume. Given the changing taper that can occur as one moves from the base to the top of the main stem, especially in the lower section from the stump to breast height, our use of the three stem sections should provide a better estimate for these trees than any existing diameter based equation would do, applied over the entire length of the main stem.
Discs of about 2 cm in thickness were cut at the base, middle, and top portion of each tree and their diameter outside the bark was measured. Wedge shaped sections of the field-moist wood (i.e. green wood) extending from the center to the edge of the discs, including bark, were removed and preserved by freezing for further analyses. Volumes of these sub-samples were measured by water displacement via suspension in a vessel of water placed on an electronic balance. Sample green volume (cm3) was determined as the increase in balance reading (g) due to the suspended wood sample. These subsamples were soaked in water for approximately 30 min before volume determination to minimize effects of short-term water absorption during displacement measurement on estimated volume. Individual wood samples were stored in plastic bags in a freezer (0 °C) at the Forestry Research Institute of Ghana, until samples could be transported to Michigan Technological University. Wood samples were then oven dried at 70 °C for 48 h and weighed. The 70 °C temperature was used because of the potential for increased loss of volatile organic compounds at higher temperatures, such as 100 °C. Wood density (g/cm3) was calculated as dry mass divided by green volume. This is the appropriate wood density value for converting our estimated main stem green volume to biomass, but may differ slightly from basic wood density, which would have required soaking the samples in water for several days before volume measurement, to ensure they were at maximum volume. The samples were ground to a fine powder using a ball mill (Spex certi-Prep 8000 M) and analyzed for carbon concentration using an elemental analyzer (model NA 1500NC, CE Elantech, Lakewood NJ, USA).
For each stem section from a tree, we multiplied its estimated volume by its wood density to determine the biomass of the stem section. The biomass of each stem section was then multiplied by the stem section’s C concentration to determine its C content. The total biomass (or C content) for the main stem of each sample tree was then estimated by summing the respective values for the tree’s bottom, middle and top stem sections.
Data analysis
Statistical significance for all analyses was accepted at the 0.05 level of probability. Differences among species in estimated main stem volume and carbon content were determined using one-factor analysis of variance (ANOVA), with separate analyses conducted for the age twelve and age seven species. Significant species effects for the age twelve trees in these analyses were further examined through mean separation using Tukey’s HSD test. Two-factor ANOVAs were used to test for significance of the effects of stem position and species on wood density and carbon concentration. Separate analyses were conducted for the seven- and twelve-year old trees. Differences among species and stem positions in the analyses for the age 12 trees were further examined through mean separation using Tukey’s HSD test. For the three species for which both age 7 and age 12 data were available, a three-factor ANOVA was used to test for the effects of species, stem section and age on wood density. For the two Khaya species at age 5, the effect of growth in different ecozones (wet evergreen vs. moist semi-deciduous) on wood density was examined by three-factor (species, stem position, ecozone) ANOVA. Differences in estimated main stem C content between ecozones for the two Khaya species were tested with a two-factor (species, ecozone) ANOVA. For the age 12 trees, we also used a one-factor ANOVA to determine if estimated main stem volume, mean wood density or C content differed among guild classifications for the sixteen indigenous species. Guild classifications used were pioneer, shade bearer, and non-pioneer light demander.
To determine if wood density was related to growth rate, correlation analyses were used to test for linear relationships between average tree wood density and three indicators of main stem growth rate: dbh, main stem height, and estimated main stem volume. This examination was performed across all 53 trees of either age 7 or age 12, regardless of species; the number of trees within species was too small (2 or 3) to allow similar analyses by species.
We used linear regression with a zero intercept to determine if the average wood density we found for a species growing in plantation was similar to that for the same species growing in natural forests. The mean density for a species in our data set was used as the dependent variable, and the average value in the literature for the species as the independent variable. A slope whose 95 % confidence limits did not include 1 would indicate a significant difference in wood density for plantation grown trees compared to those growing in natural forests.
Because dbh data is much more likely be available to those wishing to estimate a tree’s woody volume than are the diameters of several individual stem sections, we felt it was important to also develop relationships between dbh and main stem volume. We used non-linear regression to assess relationships between estimated total main stem volume (V) and dbh using a power relationship:
where a and b are regression coefficients. A similar allometric power function was developed using D2H (dbh squared × stem height) as the predictor variable.
Results
Stem volume and wood density
Average tree main stem volume for species in the 12-year-old plantation ranged from a minimum of 0.01 m3 to a maximum of 1.04 m3 (Fig. 2). The species with the greatest estimated stem volume at age 12 were Ceiba pentandra and Cedrela Odorata, while Guarea thompsonii had the lowest volume. Strong predictive relationships for total main stem volume existed for both dbh and D2H for the combined data from the 12- and 7-year old trees.

Mean main stem volumes of species in a 12 year-old plantation at Oda-kotoamso Community Agroforestry Project in the wet evergreen forest ecozone of Ghana. Mean main stem volume was significantly different for species without common letters (Tukey’s HSD, P < 0.05)
Wood density differed significantly among species within the 12-year-old plantation (P < 0.001; Table 1; Fig. 3), but not for the four species in the 7-year-old plantation (Antrocaryon micraster, Heritiera uitlis, Khaya ivorensis, and Turraeanthus africanus) (Table 2). For the 12-year-old trees, mean density ranged from 0.27 to 0.76 g/cm3 (Table 2). Ceiba pentandra had the lowest density, while Lophira alata had the highest density. Comparison of wood density for the same species from 12- and 7-year-old plantations in the wet ecozone indicated no differences among the ages (P = 0.923 for age effect in 3 factor ANOVA). There was a significant difference in wood density of Khaya spp. of the same age (5 years) planted in different ecozones (Fig. 4), with wood density in trees from the wet evergreen zone being higher than that found in trees of the same species from the moist semi-deciduous zone.

Wood density estimates of trees species from a 12-year-old plantation at Oda-kotoamso Community Agroforestry Project in the wet evergreen forest ecozone of Ghana. Wood density differed significantly (P < 0.001) among tree species and among stem positions. Error bars indicate one standard deviation for a stem zone within a species. Mean wood density was significantly different for species without common letters (Tukey’s HSD, P < 0.05). Across species, mean wood density for the bottom stem position was significantly greater than that for the middle and top stem positions (Tukey’s HSD, P < 0.05)

Comparison of wood density in 5-year-old Khaya from the wet evergreen and moist semi-deciduous ecozones of Ghana. Error bars are one standard deviation. Wood density is significantly lower in the moist semi-deciduous ecozone (P = 0.026)
Density differed significantly with position along the stem of the trees (Fig. 3), with the bottom positions having greater mean wood density (0.526 g cm−3) than the middle and the top (0.444 and 0.439 g cm−3, respectively). No interactions were found between species and the three stem positions for wood density (Table 1), indicating that greater wood density for the bottom stem position occurred consistently for the species studied. Across species, wood density was negatively correlated with indicators of growth rate for 12-year-old trees planted in the wet forest ecozone, with negative linear correlations existing within the 41-tree data set between average tree wood density and dbh at age 12 (r = −0.51, P < 0.001), stem length (r = −0.63, P < 0.001) and estimated main stem volume (r = −0.54, P < 0.001).
Carbon concentration and content
Carbon (C) concentration also differed significantly among species and with position along the stem of the trees (Table 1, Fig. 5). The bottom positions of trees had lower C concentration (470 g kg−1) than the middle and the top, which had mean C concentrations of 478 and 477 g kg−1, respectively (Fig. 5). Species by stem position interactions did not occur for wood C concentration (Table 1, P = 0.066), indicating that lower C concentration in the bottom stem position was generally consistent across species, but that some variability in the pattern occurred (Fig. 5).

Wood C concentration by stem position for trees species from a 12-year-old plantation at OCAP in the wet evergreen forest ecozone of Ghana. Carbon concentration differed significantly among tree species and among stem positions (P < 0.001). Error bars indicate standard deviation for a stem zone within a species. Mean wood C concentration was significantly different for species without common letters (Tukey’s HSD, P < 0.05). Across species, mean wood C concentration for the bottom stem position was significantly lower than that for the middle and top stem positions (Tukey’s HSD, P < 0.05)
The mean stem C content across species was 55 kg C. Guarea thompsoni had the lowest average stem carbon content of 2 kg C per tree whereas Ceiba pentandra sequestered the greatest stem C at age 12 of 179 kg per tree (Fig. 6). There was significant variation in average main stem C content among species in the 12- year-old plantation (P < 0.001). However, there were no C content differences among the more limited set of species from the 7-year-old plantation (P = 0.834). Similarly, stem C content of five-year-old Khaya spp. did not differ between the wet evergreen and moist semi-decidous ecozones.

Mean main stem C content per tree for species growing in a 12-year-old plantation at OCAP in the wet evergreen forest ecozone of Ghana. Mean main stem C content was significantly different for species without common letters (Tukey’s HSD, P < 0.05)
Relationships to guilds classification
Apart from Tectona grandis and Cedrela robusta, which are exotic species, all other (i.e. indigenous) tree species were classified into three guilds: pioneers, non-pioneer light demanders, and shade bearers (Hawthorne 1995). Using this classification enabled us to determine if inherent ecological strategies were related to wood density, growth rate or C content of these tropical tree species when grown in plantations. At age 12, the three guild classifications did not differ in wood density. Pioneers and shade bearers had significantly higher main stem volume, biomass and carbon content than non-pioneer light demanders, but significant differences between pioneers and shade bearers did not exist.
Discussion
Due to the relatively small sample size within species, we remind readers that values for individual species in the study should be viewed with caution, and additional data in the literature for the species should be considered if available. In addition, the estimated main stem volume and biomass for our trees at ages seven and twelve may be specific to the area studied. The fairly low variability within species for many parameters is likely due at least in part to the sampling only a few trees of a species from a small area. Greater variability would be expected for a broader sampling across a range of locations and site conditions. Our strongest findings are those that occurred consistently across all species studied, such as greater wood density and lower C concentration at the bottom stem position, lower wood density for plantation grown trees than for the same species growing in natural forests, and C concentrations that are consistently below 500 g kg−1.
Stem volume and wood density
Knowledge of wood density is needed to convert the volume of forest trees to biomass (Brown et al. 1989; Fearnside 1997), which can then be used to estimate carbon content (Brown and Lugo 1982; Zianis and Mencuccini 2004). Wood density can vary greatly among species (Redondo-Brenes and Montagnini 2006; Henry et al. 2010; Sotelo Montes et al. 2012), as was observed in this study (Table 1; Fig. 3). Wood density was significantly different among the stem positions used in this study and generally increased from top to bottom in the main stem. This observation has been noted in similar studies elsewhere (Espinoza 2004; Nogueira et al. 2005; Weber and Sotelo Montes 2005, 2008). In particular, mean density of trees in Amazon forests is known to decrease from breast height to the top of the bole (Nogueira et al. 2005). The variation in densities of the three positions for the main stem reported here, needs to be taken into account in biomass estimation of tropical trees because these differences could lead to potential errors. Across species, bottom stem density was on average 12 % greater than average wood density across all stem positions, which would cause corresponding overestimates of main stem biomass if only the bottom stem density was used for converting volumes into biomass.
For several species, wood density was similar to previously published results (Table 2). For example, Reyes et al. (1992) reported average densities of 0.26 g cm−3 for Ceiba pentandra and 0.45 g cm−3 for Entandrogma angolense. These values are quite close to the mean values of 0.27 and 0.44 g cm−3, respectively, reported in this study. For other species, we often found slightly lower wood density in the plantation grown trees of this study than previously reported for the same species growing in natural stands (Table 2). Elias and Potvin (2003) similarly found that wood from natural forest individuals had higher C concentrations than that from plantation grown trees of the same species. Overall, a regression of our wood density values versus average values in the literature produces a highly significant linear relationship (r = 0.89, P < 0.001), with a slope of 0.92 (Fig. 7), indicating the tendency for the young plantation grown trees in this study to have lower wood density than trees of the same species growing in natural forests.

Relationship between densities found in this study for plantation grown trees and published values for the same species from native forests. The dark solid line indicates the linear regression, with a zero intercept, for predicting plantation density from literature values from natural forests. The 95 % confidence limits for the slope of this line are lower than 1 (0.862–0.975). The thin solid line is a 1:1 relationship. Published values were not available for two of the eighteen species studied, hence there are only sixteen data points
There is often an inverse relationship between wood density and rate of volume growth (Thomas 1996; Redondo-Brenes and Montagnini 2006, Weber and Sotelo Montes 2008), especially when comparisons are made across species that vary greatly in growth rate. Within species, relationships can be much more variable, with inverse relationships common but neutral and positive relationships also occurring (de Castro et al. 1993; Weber and Sotelo Montes 2005, 2008). In West Africa, a positive within-species relationship was observed between tree growth and density for both Balanites aegyptiaca (L.) Delile and Prosopis africana (Guill. & Perr.) Taub (Sotelo Montes and Weber 2009; Weber and Sotelo Montes 2010).
Across species in our study, we found that negative relationships existed between wood density and measures of growth rate. It is possible that a similar effect of greater growth rate leading to lower wood density may have contributed to our tendency to find slightly lower wood density than reported in the literature for a given species. Individuals in the plantations are not light limited to the degree that similar small trees in a natural forest would be. This could allow for higher growth rates, which might contribute to lower density. Henry et al. (2010) examined several of the same species used in this study in a natural forest, also in the wet evergreen ecozone in Ghana, and found higher wood density values. In their study, the smallest Tieghemella heckelii and Heritiera utilis examined were similar in diameter to the plantation trees we studied, and had densities of 0.58–0.78 g cm−3, which are 34–47 % greater than densities we measured for the same species growing in plantations. The age of the trees in Henry et al.’s study is not known, but it is likely that they are older than the trees in our study, as it would typically take individuals a longer time to achieve a given size when grown under an existing canopy than when grown in the open. In trees from natural forests, there is a tendency for the smallest trees of these species to have the highest density (Henry et al. 2010). This may reflect slower growth rate experienced by shaded individuals in the understory, leading to higher wood density than we found in similar sized, but faster growing trees in the high light environment of a young plantation. Overall, it appears that the wood density of small trees from natural tropical forests is not indicative of the wood density that would occur in similar sized trees of the same species in plantations. In fact, the published values for larger trees in natural forests (e.g. Henry et al. 2010) seem more suitable.
Densities of Khaya species at age five grown in the moist semi-deciduous ecozone were lower than those for Khaya grown in the wet evergreen forest area. It is known that wood density can vary from one geographic location to another (Wiemann and Williamson 2002; Baker et al. 2004) and may be positively associated with precipitation (Maharjan et al. 2011). This could be responsible for higher density for the wetter forest zone (Wiemann and Williamson 2002). However, in contrast to these findings, Steege and Hammond (2001) reported that wood density is not correlated with precipitation or soil fertility, and Sotelo Montes and Weber (2009) found a negative relationship between wood density and precipitation. Additional research on this topic is needed, as the 23 % lower density we found for Khaya in the moist semi-deciduous ecozone indicates that the application of wet evergreen ecozone values to plantations planned for nearby ecozones in Ghana (both moist semi-deciduous and dry semidecidous) might lead to substantial overestimations of biomass and C sequestration.
Carbon content and guilds classification
Tree carbon content was significantly different among species in the 12-year-old plantation but did not differ among the guilds classification (pioneers, non- pioneer light demanders, and shade bearers) in relationship to the light environment each is presumed to prefer. Ceiba pentandra had the greatest carbon content, belonged to the pioneer category, and had the highest volume growth rate and lowest density. However, other pioneer species such as Lophira alata and Triplochiton scleroxylon had fairly low volume, biomass and carbon content at age 12, and mean values for pioneers did not differ from mean values for shade bearers. These results are thus only in partial agreement with Henry et al. (2010), who did find significantly lower wood density for pioneers than shadebearers, and Redondo-Brenes and Montagnini (2006) who showed that fast growing trees contain extremely high aboveground biomass, despite often having lower density. In natural forests, the shadebearers may have spent their entire lifespan in a low light environment, leading to slow growth and higher wood density. As stated earlier, the high light environment experienced by shadebearers in the young plantations of our study may have led to faster growth and lower wood density, eliminating the differences among guilds found in natural forests.
The faster growing species in this study (Ceiba pentandra, Cedrela odorata, and Turraeanthus africanus) accumulated the highest biomass and C content despite their tendency to have lower wood density. Estimated stem volume at age 12 was the greatest contributor to C content in this study, but species differences in wood density and C concentration also were important factors. Of these factors, wood density was much more variable among the tree species (0.27–0.76 g cm−3), and thus had a greater influence on the C content differences among species than did C concentration. Carbon concentration for all species was less than the commonly assumed value of 50 % (500 g C kg−1), and thus it is important that species-specific values be used when available. Our samples were oven-dried, and Martin and Thomas (2011) have recently reported that oven drying can cause the loss of volatile carbon compounds from tropical wood samples. Based on their results, our estimates may be low by as much as 12 g C kg−1. Even with this adjustment, our values are virtually all still less than 500 g C kg−1.
Plantation development in Ghana has focused on promoting mixtures of fast growing species primarily to provide raw material for industry. The development of these plantations also can play a useful environmental role in C sequestration and storage (Winjum and Schroeder 1997; Schroeder 1992; Nair et al. 2009). However, as shown here, some slower growing species also might have good potential to contribute to carbon sequestration, due to high wood density.
Conclusions
This research was undertaken to provide information needed to aid the assessment of the C content of tropical trees grown in forest plantations in Ghana and the surrounding region, as well as other areas of the world having similar tropical climates. Species differences in the parameters necessary for estimating C content, such as wood density, C concentration and volume were investigated. The results of this study revealed significant differences among tree species in wood density and carbon concentration, with both also varying significantly among the bottom, middle and top positions of the main stem. The common assumption of half of wood biomass being C did not hold, as an average value of 476 g C kg−1 was observed across species, with no individual species having an average value of 500 g C kg−1. There was wide variability in wood density among species, and it had a stronger influence on species differences in stem C content than did C concentration. In addition, it appears that wood density for young plantation grown trees is often lower than that for the same species growing in natural forests. Densities of Khaya spp. grown in the moist semi-deciduous ecozone were lower than those of trees grown in wet evergreen forest area. To clarify the cause of the differences related to ecozone, further research is needed to examine the environmental and edaphic factors influencing wood density in Ghana and similar tropical areas with widely varying ecozones. In particular, efforts should be geared towards scrutinizing the effects of soil fertility, rainfall and elevation on wood density. We feel general values for wood density should not be applied across all species when calculated volumes are converted to biomass.
References
Avery TE, Burkhart HE (2002) Forest measurements. McGraw Hill, New York
Baker TR, Phillips OL, Malhi Y, Almeida S, Arroyo L, Di Fiore A, Erwin T, Killeen TJ, Laurance SG, Laurance WF, Lewis SL, Lloyd J, Monteagudo A, Neill DA, Patino S, Pitman NCA, Silva JNM, Vasquez Martinez R (2004) Variation in wood density determines spatial patterns in Amazonian forest biomass. Global Change Biol 10:545–562
Bert D, Danjon F (2006) Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.). For Ecol Manage 222:279–295
Bolza E, Keating WG (1972) African timbers: the properties, uses and characteristics of 700 species. Division of Building Research, CSIRO, Melbourne
Brown S, Lugo AE (1982) The storage and production of organic-matter in tropical forests and their role in the global carbon-cycle. Biotropica 14:161–187
Brown S, Gillespie AJR, Lugo AE (1989) Biomass estimation methods for tropical forests with applications to forest inventory data. For Sci 35:881–902
Chaturvedi RK, Raghumbanshi AS (2013) Aboveground biomass estimation of small diameter woody species of tropical dry forest. New Forest 44:509–519
Chave J, Andalo C, Brown S, Cairns MA, Chambers JQ, Eamus D, Fölster H, Fromard F, Higuchi N, Kira T, Lescure J-P, Nelson BW, Ogawa H, Puig P, Riéra B, Yamakura T (2005) Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145:87–99
Christensen JH, Hewitson B, Busuioc A, Chen A, Gao X, Held I, Jones R, Kolli RK, Kwon W-T, Laprise R, Magaña Rueda V, Mearns L, Menéndez CG, Räisänen J, Rinke A, Sarr A, Whetton P (2007) Regional climate projections. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL (eds) Climate change 2007: the physical science basis. Contribution of working group I to the fourth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
de Castro F, Williamson GB, de Jesus RM (1993) Radial variation in the wood specific gravity of Joannesia princep: the roles of age and diameter. Biotropica 25:176–182
Desanker PV (2005) The Kyoto Protocol and the CDM in Africa: a good idea but … Unasylva 222:24–26
Elias M, Potvin C (2003) Assessing inter- and intra-specific variation in trunk carbon concentration for 32 neotropical tree species. Can J For Res 33:1039–1045
Espinoza JA (2004) Within-tree density gradients in Gmelina arborea in Venezuela. New Forest 28:309–317
FC (2006) National forest plantation development programme: the modified Taugya system and private developers. In: Annual report for 2008, Forestry Commission, Government of Ghana
Fearnside PM (1997) Wood density for estimating forest biomass in Brazilian Amazonia. For Ecol Manage 90:59–87
Gibbs HK, Brown S, Niles JO, Foley JA (2007) Monitoring and estimating tropical forest carbon stocks: making REDD a reality. Environ Res Lett 2:045023
Hall JB, Swaine MD (1981) Distribution and ecology of vascular plants in a tropical rain forest: forest vegetation in Ghana. Dr W Junk, The Hague, the Netherlands
Hawthorne WD (1995) Ecological profiles of Ghanaian forest trees. Oxford Forestry Institute, Department of Plant Sciences, University of Oxford, Oxford, UK
Henry M, Besnard A, Asante WA, Eshun J, Adu-Bredu S, Valentini R, Bernoux M, Saint-Andre L (2010) Wood density, phytomass variations within and among trees, and allometric equations in a tropical rainforest of Africa. For Ecol Manage 260:1375–1388
ITTO (2006) Guide book for the formulation of afforestation and reforestation projects under the clean development mechanism. In: Pearson T, Walker S, Brown S (eds) Technical series 25. International Tropical Timber Organization, Yokohama, p 53
Kalame FB, Aicloo R, Nkem J, Ajayie OC, Kanninen M, Luukkanen O, Idinoba M (2011) Modified taungya system in Ghana: a win–win practice for forestry and adaptation to climate change? Environ Sci Policy 14:519–530
Ketterings QM, Coe R, van Noordwijk M, Ambagau Y, Palm CA (2001) Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. For Ecol Manage 146:199–209
Lamlom SH, Savidge RA (2006) Carbon content variation in boles of mature sugar maple and giant sequoia. Tree Physiol 26:459–468
Luizão RCC, Luizão FJ, Paiva RQ, Monteiro TF, Sousa LS, Kruijt B (2004) Variation of carbon and nitrogen cycling processes along a topographic gradient in a central Amazonian forest. Global Change Biol 10:592–600
Maharjan SK, Poorter L, Holmgren M, Bongers F, Wiering JJ, Hawthorne WD (2011) Plant functional traits and the distribution of west African rain forest trees along the rainfall gradient. Biotropica 43:552–561
Martin AR, Thomas SC (2011) A reassessment of carbon content in tropical trees. PLoS ONE 6(8):e23533
Nair PKR, Kumar BM, Nair VD (2009) Agroforestry as a strategy for carbon sequestration. Z Pflanzenernähr Bodenk 172:10–23
NOAA (2013) Trends in atmospheric carbon dioxide. US National Oceanic and Atmospheric Administration. http://www.esrl.noaa.gov/gmd/ccgg/trends/global.html. Accessed 5 April 2013
Nogueira EM, Nelson BW, Fearnside PM (2005) Wood density in dense forest in central Amazonia, Brazil. For Ecol Manage 208:261–286
Preece ND, Crowley GM, Lawes MJ, van Oosterzee P (2012) Comparing above-ground biomass among forest types in the wet tropics: small stems and plantation types matter in carbon accounting. For Ecol Manage 264:228–237
Redondo-Brenes A, Montagnini F (2006) Growth, productivity, aboveground biomass, and carbon sequestration of pure and mixed native tree plantations in the Caribbean lowlands of Costa Rica. For Ecol Manage 232:168–178
Reyes G, Brown S, Chapman J, Lugo AE (1992) Wood densities of tropical tree species. Gen Tech Rep SO-88. US Dept of Agriculture, Forest Service, Southern Forest Experiment Station, New Orleans, LA, Gen Tech Rep SO-88, 15 p
Sandker M, Nyame SK, Forster J, Collier N, Shepherd G, Yeboah D, Ezzine-de Blas D, Machwitz M, Vaatainen S, Garedew E, Etoga G, Ehringhaus C, Anati J, Quarm ODK, Campbell BM (2010) REDD payments as incentive for reducing forest loss. Conserv Lett 3:114–121
Schneider SH (1990) The global warming debate heats up: an analysis and perspective. Bull Am Meteor Soc 71:1292–1304
Schroeder P (1992) Carbon storage potential of short rotation tropical tree plantations. For Ecol Manage 50:31–41
Sicard C, Saint-Andre L, Gelhaye D, Ranger J (2006) Effect of initial fertilisation on biomass and nutrient content of Norway spruce and Douglas-fir plantations at the same site. Trees Struct Funct 20:229–246
Slik JWF, Bernard CS, Breman FC, van Beek M, Salim A, Sheil D (2008) Wood density as a conservation tool: quantification of disturbance and identification of conservation-priority areas in tropical forests. Conserv Biol 22:1299–1308
Solomon S, Qin D, Manning M, Alley RB, Berntsen T, Bindoff NL, Chen Z, Chidthaisong A, Gregory JM, Hegerl GC, Heimann M, Hewitson B, Hoskins BJ, Joos F, Jouzel J, Kattsov V, Lohmann U, Matsuno T, Molina M, Nicholls N, Overpeck J, Raga G, Ramaswamy V, Ren J, Rusticucci M, Somerville R, Stocker TF, Whetton P, Wood RA, Wratt D (2007) Technical summary. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL (eds) Climate change 2007: the physical science basis. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
Sotelo Montes C, Weber JC (2009) Genetic variation in wood density and correlations with tree growth in Prosopis africana from Burkina Faso and Niger. Ann For Sci 66:713
Sotelo Montes C, Weber JC, Silva DA, Andrade C, Muñiz GIB, Garcia RA, Kalinganire A (2012) Effects of region, soil, land use and terrain type on fuelwood properties of five tree/shrub species in the Sahelian and Sudanian ecozones of Mali. Ann For Sci 69:747–756
Steege HT, Hammond DS (2001) Character convergence, diversity, and disturbance in tropical rain forest in Guyana. Ecology 82:3197–3212
Suzuki E (1999) Diversity in specific gravity and water content of wood among Bornean tropical rain forest trees. Ecol Res 14:211–224
Thomas SC (1996) Asymptotic height as a predictor of growth and allometric characteristics in Malaysian rain forest trees. Am J Bot 83:556–566
Wauters JB, Coudert S, Grallien E, Jonard A, Ponette Q (2008) Carbon stock in rubber tree plantations in Western Ghana and Mato Grosso (Brazil). For Ecol Manage 255:2347–2361
Weber JC, Sotelo Montes C (2005) Variation and correlations among stem growth and wood traits of Calycophyllum spruceanum Benth. from the Peruvian Amazon. Silvae Genetica 54:31–41
Weber JC, Sotelo Montes C (2008) Geographic variation in tree growth and wood density of Guazuma crinita Mart. in the Peruvian Amazon. New Forest 36:29–52
Weber JC, Sotelo Montes C (2010) Correlations and clines in tree growth and wood density of Balanites aegyptiaca (L.) Delile provenances in Niger. New Forest 39:39–49
Wiemann MC, Williamson GB (2002) Geographic variation in wood specific gravity: effects of latitude, temperature, and precipitation. Wood Fiber Sci 34:96–107
Winjum JK, Schroeder PE (1997) Forest plantations of the world: their extent, ecological attributes, and carbon storage. Agric For Meteor 84:153–167
World Bank (2009) Carbon finance guide for communities. In: Maryanne G, Muyeye C, Barry K, Thais C (eds) Working paper volume 1. World Bank, Washington, DC
Zianis D, Mencuccini M (2004) On simplifying allometric analyses of forest biomass. For Ecol Manage 187:311–332
Acknowledgments
We would like to express our sincere appreciation to SamartexTimber and Plywood Limited (Samartex) for providing study locations and logistical support during sampling. We particularly acknowledge Richard Nsenkyire (General Manager), KKF Ghartey (Forestry Consultant) for their support at Samartex. We thank Crispin Suglo, Nelson Amelodze, G.K.A Acolatse, Lord Amenyaw, Sandra Owusu, and Millicent Obeng, field technicians and students at both the Forestry Research of Ghana and Samartex for their help in field data collection and lab support in Ghana. Jennifer Eikenberry and Emmanuel Ebanyenle of Michigan Technological University provided lab assistance and editorial support, respectively. The study was funded by the International Tropical Timber Organization, the Forestry Research Institute of Ghana, the Ecosystem Science Center at Michigan Technological University, and Michigan Technological University through the graduate research assistance training program.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yeboah, D., Burton, A.J., Storer, A.J. et al. Variation in wood density and carbon content of tropical plantation tree species from Ghana. New Forests 45, 35–52 (2014). https://doi.org/10.1007/s11056-013-9390-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-013-9390-8