
Abstract
Fall nutrient loading of deciduous forest nursery seedlings is of special interest because of foliage abscission and varied translocation patterns. For non-deciduous seedlings in the nursery, fall fertilization typically can reverse nutrient dilution and possibly increase nutrient reserves; however, this technique has received little attention with deciduous conifer trees that translocate nutrients before abscising foliage. We evaluated how fall nitrogen (N) fertilization affected N storage and translocation in the deciduous conifer Olga Bay larch (Larix olgensis Henry) seedlings during the hardening period. Seedlings were supplied with 25 mg N seedling−1 for 15 weeks before hardening and fall fertilization treatments began with a three week application period of K15NO3 at 0, 5, 10 and 15 mg N seedling−1. During the hardening period, fall N fertilization had little effect on seedling morphology. The N concentration and content of needles decreased dramatically as needles abscised, while that of stems and roots increased. Six weeks after fall N fertilization ceased, all seedlings translocated similar net N from their needles. For the control seedlings, this accounted for 84 % of the N stored in stems and roots. For fall fertilized seedlings, however, the proportion of N stored in stems and roots translocated from needles accounted for only 41–61 % of the total because of absorption of fall fertilizer that was translocated directly to stems and roots. Six weeks after fall fertilization, the distribution pattern of N concentration and content in seedlings was found in this order: stems > fine roots > coarse roots > needles. Our results suggest that providing deciduous conifer seedlings N during hardening, in this case Olga Bay larch, is a way to promote nutrient loading during nursery production.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Fast-growing and high-yielding forest plantations play an important role in effectively addressing the imbalance between supply and demand in China’s timber market. In northeast China, Olga Bay larch (Larix olgensis Henry) is a popular timber species because of its rapid early growth rate and use in a wide variety of products. Because of the poor natural regeneration of this species (Wang and Zhang 1992; Liu 1997), efforts to reforest large areas with Olga Bay larch have focused on outplanting seedlings from nurseries. In China, more than half of the remaining sites in need of afforestation are difficult to establish, and will require seedlings of high quality (Liu et al. 2012). The current national standard specifies that only 1 + 1 transplants be used in plantations to ensure seedling survival (State Forestry Administration 2003); although, 1 + 0 seedlings are also outplanted in some regions because of their low cost. Li et al. (2011) and Duan et al. (2013) have shown the feasibility of using large 1 + 0 Olga Bay larch for afforestation. Thus, if 1 + 0 seedlings of high quality could be produced, land managers may obtain seedlings with lower costs in less time, as well as obtain more successful plantation establishment.
Increasing the preplanting nutrient status of seedlings by fall fertilization has been shown to be an optional method to enhance the growth of newly outplanted seedlings. Irwin et al. (1998) found foliar nitrogen (N) of Pinus elliottii var. elliottii Engelm. seedlings increased significantly with fall fertilizer treatment, and first year survival and heights were 15 % greater than unfertilized seedlings, and Boivin et al. (2004) demonstrated that fall fertilization of Picea mariana (Mill.) BSP. increased N uptake 44–167 % during hardening and biomass 115 % 13 weeks after outplanting.
Supply of extra nutrients during the hardening stage (fall fertilization) can prevent nutrient dilution and consistently stimulate N accumulation in seedlings (Timmer 1997; Sung et al. 1997; Rikala et al. 2004), but the most effective dose of fall fertilizer to enhance nutrient accumulation in the nursery and promote outplanting response may not be the highest one (Boivin et al. 2004). Many studies have shown that foliage is the major N sink in evergreen seedlings (van den Driessche 1985; Millard and Grelet 2010). Thus, it could be that the benefit of fall fertilizer on evergreen seedling quality, particularly enhancement of foliage nutrient reserves, may be less important, or uniquely different, for deciduous seedlings because they translocate most of their foliar nutrients into shoot and root tissues prior to leaf senescence at the end of growing season (Duchesne et al. 2001; Cheng and Fuchigami 2002). Despite this, nutrient loading does show promise as it has been observed to increase internal nutrient reserves and improve the quality and field performance of Quercus rubra seedlings (Salifu and Jacobs 2006; Salifu et al. 2009a).
Most fall fertilization studies have been conducted on evergreen species (e.g., Picea abies (L.) Karst (Rikala et al. 2004), P. mariana (Malik and Timmer 1996; McAlister and Timmer 1998; Boivin et al. 2004), Pinus taeda L. (Sung et al. 1997), and Pseudotsuga menziesii (Mirb.) Franco (van den Driessche 1985)) with few papers addressing fall fertilization of deciduous seedlings (Salifu and Jacobs 2006; Salifu et al. 2008, 2009a; Li et al. 2012). Therefore, we initiated a study to examine the effects on N translocation and storage for the deciduous conifer species Olga Bay larch using 15N to observe N movement within seedlings, similar to the technique used on the deciduous hardwood, Q. rubra (Salifu et al. 2008). We hypothesized that (1) movement of N from foliage to stems and roots during hardening would compensate for any N dilution effect in the seedling after needle senescence; (2) tissue N contents and concentrations would increase with fall-applied N fertilizer; (3) fertilizer use efficiency would decrease as the amount of fall-applied N fertilizer increased; and (4) increasing the rate of fall-applied N fertilizer would reduce the relative contribution of N translocated from needles to final stem and root N contents.
Materials and methods
Plant material and cultural conditions
Larix olgensis seeds, originally from Shanxi, China, were obtained from Lawyer Nursery (Plains, MT, USA). Seeds were soaked 48 h in running water and then stratified 28 days at 2–3 °C. On 23 May 2011, we sowed seeds into 5 trays each containing 98 Ray Leach Super Cells (164 cm3, 3.8 cm top diameter, 21 cm depth, and 528 seedlings m−2; Stuewe and Sons, Inc, Tangent, OR, USA) filled with a peat:vermiculite (1:1, v/v) medium (Sunshine Special Blend Forestry #1, Sun Gro Horticulture, Bellevue, WA, USA). Three weeks after sowing, we thinned seedlings to one per cell. Seedlings were cultured in a greenhouse (USDA Forest Service, Rocky Mountain Research Station, Moscow, ID, USA; 46.7232°N, 117.0029°W) under natural day-length with day: night temperatures averaging 28:16 °C.
Fertilization commenced on 22 June and continued for 10 weeks. Irrigation or fertigation (irrigation with soluble fertilizer added) frequency was determined gravimetrically and applied when container mass reached 65 % of field capacity. The proportion of nutrients was constant: 50 N (15NO3 −; 19NH4 +; 16urea): 44P:48K: 26Ca: 13Mg: 4S: 1.39Fe: 0.15Mn: 0.13Zn: 0.03Cu: 0.02B (Peters Professional, The Scotts Company, Marysville, OH, USA). We supplied seedlings with a total of 25 mg N seedling−1 before hardening. The hardening treatment began on 2 September, with a two-week short-day treatment (8 h light) to promote terminal bud formation. Hardening continued with a return to natural day-length and a gradual lowering of greenhouse temperatures (18:7 °C day:night). Cells were rotated biweekly within trays to minimize edge effects.
After short day exposure, we surrounded each seedling with an open-topped white netting bag to facilitate collection of abscised needles. A 4 nitrogen rate × 5 replication complete randomized design was used to test fall application levels of N:0, 5, 10, and 15 mg N seedling−1. N was supplied as KNO3 enriched to 95 atom % 15N (Sigma Aldrich, WI, USA). Three split applications were conducted on October 5, 12, and 19. To each seedling on each application date, we applied 15 mL of the appropriate N treatment (N concentration rates: 0, 0.11, 0.22 and 0.33 mg N mL−1). To reduce the amount of leaching, seedlings were irrigated less than the container capacity. During the N application period and until the end of December 2011 when the experiment terminated, greenhouse temperatures averaged 15:5 °C day: night.
Morphology and N measurement
We randomly sampled 5 seedlings from each replication (25 per treatment) before fall fertilization, and two other groups of 5 seedlings from each treatment and replicate combination were sampled at 2 and 6 weeks after fall fertilization. The timing of our fall fertilization applications coincided with the typical hardening period for 1 + 0 bareroot Olga Bay larch (Li 2009). We chose our sample dates to examine potential acute responses to fall fertilization (2 weeks) as well as the response after most needles were expected to have senesced (6 weeks). Individual seedlings were washed free of growing media and rinsed in deionized water, measured for height (root collar to tip of terminal bud) and root-collar diameter (RCD), partitioned into needles, stems, fine roots (<2 mm diameter) and coarse roots (>2 mm diameter), then oven dried at 65 °C for 72 h and weighed. We portioned roots into fine and coarse fractions because fine roots are regarded as the most important for water and nutrient uptake (Gordon and Jackson 2000). For each tissue fraction, the 5 seedlings within each fertilizer-replication combination were bulked and ground. The ratio of 15N to 14N (δ15N ‰) in each tissue were expressed relative to that of air and determined with an Elemental Analyzer (NC2500, CE Instruments, Milan, Italy) at the University of Idaho Stable Isotope Laboratory (Moscow, ID, USA).
Calculations and statistical analysis
We measured the change in N content of each tissue (net N translocation amount, NTA, mg) as follows:
where A is N content (mg) at 6 weeks after the last fall fertilization application, B is the N content prior to fall fertilization, D is the nutrient content of dead tissue between the times before and after fall fertilization 6 weeks, Le is the amount of nutrient leached. D + Le is assumed to be negligible in this short-term trial as noted by others (Millard 1976; Helmisaari 1992).
We measured the percentage change in N content of each tissue (net N translocation ratio, NTR, %) as follows:
where A and B are as described above.
We assumed that all mobile N in senescing needles would be stored in stem and roots. The percentage of needle net 15N translocation amount (NNTA) contributing to the final amount of 15N stored in the stem and roots (NNS) was calculated as follows:
where, S was the total increase of 15N (mg) in stem or roots 6 weeks after fall fertilization and NNTA was the net needle 15N translocation amount (mg).
Fall fertilizer use efficiency (FUE) was calculated with the following equation:
where T was the total 15N content (mg) of stem and roots in fertilized seedlings, C was the average 15N content (mg) in control seedlings, and E was the total amount of 15N applied (mg) 6 weeks after fall fertilization when most needles had senesced.
Data were analyzed using SPSS 16.0 software (Chicago, IL, USA). For the samples collected 2 and 6 weeks after fall fertilization, treatment means and standard errors computed from 5 individual seedlings for each replicate were used for height, RCD, and tissue biomass. We used a univariate analysis in a general linear model (GLM) by a linear mixed model to test the N concentration and content, as well as the ratio of 15N to total N by tissue type (needle, stem, and roots). The two-way analysis of variance was employed to test the effects of fixed variables (N rate and tissue type) on these dependent variables: biomass, and N concentrations and contents. For the 6-week sample, we also analyzed NTA, NTR, NNS, and FUE among treatments. For each analysis, when the effects of treatment or tissue type were significant, means were ranked according to Tukey’s studentized range test at α = 0.05.
Results
Plant growth response
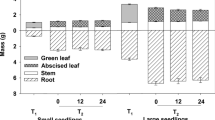
At the onset of fall N applications, the average height, RCD, and needle, stem, and root biomass of seedlings averaged 22.6 ± 1.8 cm, 2.80 ± 0.22 mm, and 0.79 ± 0.13, 0.35 ± 0.07, and 0.37 ± 0.79 g, respectively. In addition, the N concentrations of needles, stems, and roots were 11.45, 6.24 and 7.39 g kg−1, respectively, and subsequent N contents of those were 9.04, 2.21, and 2.79 mg seedling−1, respectively (Fig. 1).

The nitrogen (N) content and concentration of Olga Bay larch seedlings sampled before fall N fertilization (0) and 2 and 6 weeks after fall N fertilization at four rates: 0, 5, 10, 15 mg N seedling−1
Two weeks after cessation of fall N applications, N rate had no effect on either height (P = 0.716) or RCD (P = 0.348) (Table 1). Biomass was unaffected by the interaction of fall N application and tissue type, but was significantly affected by tissue type (Table 2). Most biomass was associated with needles, which accounted for 36 % of the total biomass, significantly more than stems and roots having diameter <2 mm (hereafter R < 2) (27 % each). Biomass of stems and R < 2 were not significantly different, but were both significantly heavier than roots having diameter >2 mm (hereafter R > 2). R > 2 accounted for 10 % of total biomass (Table 3).
Fall N rate and tissue type significantly interacted to affect N concentration and N content (Tables 2, 3; Fig. 1). N concentration of needles, regardless of N rate, decreased to about 4.76 g kg−1 (Fig. 1), whereas N concentration of stems and roots increased with increasing N application rate. The tissue N content was similar to the pattern observed for concentration.
Compared with the initial status of the seedlings, two weeks after the final N application, stem N concentration increased 161 % and N content rose to 7.44 mg, the highest amount of N in the seedling (Table 3). The next highest N content was 6.58 mg in fine roots (R < 2). N concentration in fine roots was significantly higher than the coarse roots (R > 2); all the N contents followed the same pattern. We detected no difference in N concentration of stem and roots between the 10 and 15 mg N seedling−1 treatments (Fig. 1). Needle N concentrations were similar among all N fertilization levels. The N concentrations of roots and stem in control seedlings did not show any dilution, but rather increased.
Six weeks after treatments, fall-applied N had no effect on height, needle biomass, and stems biomass, but significantly affected RCD and root biomass (Table 1). Fertilized seedlings were not significantly different from each other, but had 3.3–8.5 % more RCD than their control cohorts (P = 0.049). Biomass of R < 2 and R > 2 were affected by fall fertilization, but the average values were 0.42 and 0.16 g, respectively, similar to the control (Table 1).
Although not tested statistically, the average values of stem and root growth increased 37 and 19 %, respectively from onset of fall fertilization through 2 weeks after fall fertilization, and then appeared static (Table 3), whereas needle biomass decreased 27 and 55 % at the two sample dates after fall fertilization, compared with the value before treatment.
Except for needles, the N concentration in all the tissue types increased (Fig. 1; Table 3). Fall fertilization rate increased seedling total stem and root N content by 26–59 % (P < 0.001) and 27–69 % (P < 0.001) relative to the control. Seedling N concentration and N content differed significantly between tissues (Table 3). The interactions of the effects of tissue type and fertilization treatment were significant for N concentration and N content. The distribution pattern of N concentration and content in seedlings was found in this order: stems > fine roots > coarse roots > needles.
N translocation and utilization
Although the concentration dynamics of 15N in tissues followed the same pattern as N concentration (data not shown), we observed that the ratio of 15N to total N (hereafter 15N:N) in needles increased significantly as fall N rate increased (Fig. 2). The 15N:N remained constant in control seedlings regardless of tissue type and sample time. Six weeks after fall fertilization ceased, the averaged value of 15N:N in treated seedling roots was significantly higher than that of stems (+8.5 %) and needles (+38 %) (P < 0.001). Fall N fertilization of 15 mg yielded the highest 15N:N value, and was 21 % greater than that for seedlings receiving the lowest rate (5 mg N).

The ratio of 15N to total N in Olga Bay larch seedling tissues sampled before fall N fertilization (0), and 2 and 6 weeks after fall N fertilization at four rates: 0, 5, 10, 15 mg N seedling−1
Except for needles, net translocation amount (NTA) increased as fall N rates increased (Table 4). Fall N rate had no significant effect on the amount of N that moved out of needles (P = 0.282); the average amount across all treatments was 7.00 ± 0.75 mg N seedling−1 (Table 4). For stems, the two lowest rates (control and 5 mg N) and the two highest rates (10 and 15 mg N) were significantly different (P < 0.001); the two highest rates resulted in 50 % more translocation into stems than the two lowest rates (Table 4). The trend was similar for roots, although the control was significantly lower than the 5 and 10 mg N rates, which were not significantly different; the 15 mg N rate had significantly more translocated N than all other rates (P < 0.001). Adding 5 or 10 mg N increased the amount of N translocated to roots by 44 % compared to the control; adding 15 mg N resulted in an additional increase of 37 % compared to the lower fall fertilization rates.
Results for N translocation ratio (NTR) were similar to those for NTA. For needles, NTR was, on average, about −78 % and unaffected by N rate. In the control, translocation from needles accounted for 84 % of the N moving to stems and roots, increasing the N content of those tissues by 204 and 160 %, respectively (Table 4). Increasing amounts of N generally yielded higher stem NTR than that of needles and roots (P < 0.001). Although roots had the highest NTA, it was the stems of seedlings receiving the 10 and 15 mg N applications that had the highest positive NTR, which were 59 % greater than the average of the control and 5 mg N rates. Seedlings given fall N fertilization had 44–96 % greater NTR than control seedlings.
The percentage of N contributed by needles (NNS) to the roots and stems was significantly affected by fall N rates (P < 0.001). In the control seedlings, most of the N translocated to roots and stems (84 %) originated in the needles (Table 4). Adding 5 mg N significantly reduced this value to 61 %, and adding either 10 or 15 mg N significantly reduced it further, to about 45 %.
Fall fertilization use efficiency (FUE) decreased as N rate increased (Table 4). Although for every 5 mg N increase, FUE decreased 8.5 %, only the 5 mg and 15 mg rates were significantly different (Table 4).
Discussion
During the hardening of conifer seedlings, N dilution is a common occurrence for seedlings that are still growing (increasing biomass) in the absence of N additions (Imo and Timmer 1992; Boivin et al. 2002; Jackson et al. 2012). Our results for control seedlings show that the net loss of N from senescing needles provided 84 % of the N used for N storage, which offset dilution in those tissue, and was consistent with the 50–90 % leaf nutrient translocation into shoot and root tissues prior to senescence that was reported by Duchesne et al. (2001) and Cheng and Fuchigami (2002). Plants depend on fine roots (diameter < 2 mm) for water and mineral uptake (Gordon and Jackson 2000), indicating higher nutrient uptake activity in finer roots. Thus, we accept our first hypothesis that translocation of N from needles to stems and fine roots prior to abscission negated overall N dilution in the seedling at the end of the growing season.
As observed in other studies involving fall fertilization of evergreen seedlings (Irwin et al. 1998; Rikala et al. 2004; Islam et al. 2009), our data show that fall additions of N to the deciduous Olga Bay larch also increased tissue N concentration and content of the seedlings. Unlike evergreen seedlings where foliage, stem, and root tissues show an increase in N concentration and content, we observed reduced N in foliage concurrent with an increase of N in stems and roots. Salifu and Jacobs (2006) speculated this would occur with deciduous seedlings, and deciduous trees are known to translocate resources before leaf abscission (Aerts 1996); therefore, this result is not surprising. Our data show that Olga Bay larch seedlings translocate N from senescing needles while concurrently absorbing new N from fall fertilization (Table 2; Fig. 1); consequently, we accept our second hypothesis that tissue N concentrations and contents of this deciduous species would increase as a result of fall additions of N.
Many studies in bareroot and container nurseries have shown that FUE usually decreases with increasing rates of fertilization (Dumroese et al. 2005; Salifu et al. 2009b), especially with evergreen conifer seedlings (Timmer and Parton 1984; Timmer and Armstrong 1987). In general, only when nutrients are applied at rates to match seedling demand can fertilization enhance the nutrient uptake efficiency. Our results with deciduous larch were similar to those observed with evergreen species: FUE decreased as N rate increased (Table 4). Thus, we accept our third hypothesis.
Fall fertilized seedlings of evergreen species store most of the fall applied N in their shoots (leaves and stem) (Islam et al. 2009; Boivin et al. 2002, 2004). Much of this N is stored in photosynthetic proteins, especially RuBisCo, required for photosynthesis (Millard et al. 2007). In our study, the deciduous Olga Bay larch seedlings, because of their different foliage habit, switched at onset of needle senescence from having most N stored in their needles to having the stem and fine roots (root diameter <2 mm) become the main N sink (Fig. 1).
Indeed, we saw no evidence that our applications of N had any effect on needle N content or concentration two or six weeks after fall N applications, suggesting that all fall applied N absorbed by Olga Bay larch seedlings was stored in tissues other than needles. Further, our data showed that the N content (Fig. 1) and net N translocation ratio (NTR, Table 4) of needles six weeks after fall fertilization were similar among treatments, indicating that all larch needles decreased to a similar N content prior to abscission regardless of fall N application rate. Concurrently, stems and roots had increases in N concentration and content. These increases were more than what could be expected from the decrease of needle N, which indicates Olga Bay larch seedlings absorbed fall applied N, similar to results with Acer pseudoplatanus L. (Millard and Proe 1991).
Comparing the ratio of 15N and total N content of tissues in seedlings receiving fall applied N with those of control seedlings, the fall fertilized seedlings absorbed more 15N as application rate increased (Fig. 2). Within each fall-applied N level, the ratio of 15N and total N content in roots was significant higher than that of stems or needles, suggesting that larch roots acquire more new N in relation to the other tissue types, similar to the results of fertilizer uptake of peach and pear trees in autumn (Sanchez et al. 1991; Munoz et al. 1993). Because the needle N translocation amount of all treatments was similar, we accept our fourth hypothesis that increasing the rates of fall applied N would decreased the relative contribution of N translocation from needles to stems and roots.
Fall fertilization may be a useful method during larch seedling culture to produce high-quality seedlings for field planting. Our research shows that 84 % of the N stored in stems and roots of control seedlings after needle senescence was translocated there from the needles before they abscised. The amount of N stored in stems and roots can further be augmented when N is applied in fall immediately before onset of needle senescence in larch, and can be done so without affecting seedling morphology. Fall N fertilization significantly increased the N stature of one year old Olga Bay larch seedlings, similar to the effects of exponential fertilization applied to 1 + 0 seedlings (Duan et al. 2013). Enhanced N status has been shown to improve the outplanting performance of evergreen seedlings. This has led to an increased attention on deciduous forest seedling morphology and nutrient translocation reaction to nutrient-loading (Salifu et al. 2009a, b; Li et al. 2012). Further investigation is required, however, on nutrient carbohydrate and outplanting performance of fall fertilized deciduous conifer species such as larch, particularly in cold areas such as northeast China.
References
Aerts R (1996) Nutrient resorption from senescing leaves of perennials: are there general patterns? J Ecol 84:597–608
Boivin JR, Miller BD, Timmer VR (2002) Late-season fertilization of Picea mariana seedlings under greenhouse culture: biomass and nutrient dynamics. Ann For Sci 59:255–264
Boivin JR, Salifu KF, Timmer VR (2004) Late-season fertilization of Picea mariana seedlings: intensive loading and outplanting response on greenhouse bioassays. Ann For Sci 61:737–745
Cheng L, Fuchigami LH (2002) Growth of young apple trees in relation to reserve nitrogen and carbohydrates. Tree Physiol 22:1297–1303
Duan J, Xu C, Jacobs DF, Ma L, Wei H, Jiang L, Ren J (2013) Exponential nutrient loading shortens the cultural period of Larix olgensis seedlings. Scand J Forest Res. doi:10.1080/02827581.2013.778328
Duchesne L, Ouimet R, Camire C, Houle D (2001) Seasonal nutrient transfers by foliar resorption, leaching, and litter fall in a northern hardwood forest at Lake Clair Watershed, Quebec, Canada. Can J For Res 31:333–344
Dumroese RK, Page-Dumroese DS, Salifu KF, Jacobs DF (2005) Exponential fertilization of Pinus monticola seedlings: nutrient uptake efficiency, leaching fractions, and early outplanting performance. Can J For Res 35:2961–2967
Gordon WS, Jackson RB (2000) Nutrient concentrations in fine roots. Ecology 81:275–280
Helmisaari H (1992) Nutrient retranslocation within the foliage of Pinus sylvestris. Tree Physiol 10:45–58
Imo M, Timmer VR (1992) Nitrogen uptake of mesquite seedlings at conventional and exponential fertilization schedules. Soil Sci Soc Am J 56:927–934
Irwin KM, Duryea ML, Stone EL (1998) Fall-applied nitrogen improves performance of 1–0 slash pine nursery seedlings after outplanting. South J Appl For 22:111–116
Islam MA, Apostol KG, Jacobs DF, Dumroese RK (2009) Fall fertilization of Pinus resinosa seedlings: nutrient uptake, cold hardiness, and morphological development. Ann For Sci 66:704–713
Jackson DP, Dumroese RK, Barnett JP (2012) Nursery response of container Pinus palustris seedlings to nitrogen supply and subsequent effects on outplanting performance. For Ecol Manage 265:1–12
Li GL (2009) Studies on techniques of irrigating, fertilizing, and grading Larix olgensis seedlings [D]. Beijing Forestry University, Beijing
Li GL, Liu Y, Zhu Y, Yang J, Sun HY, Jia ZK, Ma LY (2011) Influence of initial age and size on the field performance of Larix olgensis seedlings. New For 42:215–226
Li GL, Liu Y, Zhu Y, Li QM, Dumroese RK (2012) Effect of fall-applied nitrogen on growth, nitrogen storage, and frost hardiness of bareroot Larix olgensis seedlings. Silva Fennica 46:345–354
Liu QJ (1997) Structure and dynamics of the subalpine coniferous forest on Changbai Mountain, China. Plant Ecol 132:97–105
Liu Y, Bai SL, Zhu Y, Li GL, Jiang P (2012) Promoting seedling stress resistance through nursery techniques in China. New For 43:639–649
Malik V, Timmer VR (1996) Growth, nutrient dynamics, and interspecific competition of nutrient-loaded black spruce seedlings on a boreal mixedwood site. Can J For Res 26:1651–1659
McAlister JA, Timmer VR (1998) Nutrient enrichment of white spruce seedlings during nursery culture and initial plantation establishment. Tree Physiol 18:195–202
Millard P (1976) Effect of nitrogen supply on nutrient in litter fall and crown leaching in a stand of Corsican pine. J Appl Ecol 13:233–256
Millard P, Grelet G (2010) Nitrogen storage and remobilization by trees: ecophysiological relevance in a changing world. Tree Physiol 30:1083–1095
Millard P, Proe MF (1991) Leaf demography and the seasonal internal cycling of nitrogen in sycamore (Acer pseudoplatanus L.) seedlings in relation to nitrogen supply. New Phytol 117:587–596
Millard P, Sommerkorn M, Grelet G-A (2007) Environmental change and carbon limitation in trees: a biochemical, ecophysiological and ecosystem appraisal. New Phytol 175:11–28
Munoz N, Guerri J, Legaz F, Primo-Millo E (1993) Seasonal uptake of 15N-nitrate and distribution of absorbed nitrogen in peach trees. Plant Soil 150:263–269
Rikala R, Heiskanen J, Lahti M (2004) Autumn fertilization in the nursery affects growth of Picea abies container seedlings after transplanting. Scand J For Res 19:409–414
Salifu KF, Jacobs DF (2006) Characterizing fertility targets and multi-element interactions for exponential nutrient loading of Quercus rubra seedlings. Ann For Sci 63:231–237
Salifu KF, Apostol KG, Jacobs DF, Islam MA (2008) Growth, physiology, and nutrient retranslocation in nitrogen-15 fertilized Quercus rubra seedlings. Ann For Sci 65(101):1–8
Salifu KF, Jacobs DF, Birge ZKD (2009a) Nursery nitrogen loading improves field performance of bareroot oak seedlings planted on abandoned mine land. Restor Ecol 17:339–349
Salifu KF, Islam MA, Jacobs DF (2009b) Retranslocation, plant and soil recovery of nitrogen-15 applied to bareroot black walnut seedling. Commun Soil Sci Plan 40:1408–1417
Sanchez EE, Righetti TL, Sugar D, Lombard PB (1991) Recycling of nitrogen in field-grown ‘Comice’ pears. J Hort Sci 66:479–486
State Forestry Administration (2003) The national compilation of technical standards for silviculture—volume of seeds and seedlings. In: Tree seedling quality grading of major species for afforestation. Chinese Standard Press, Beijing, pp 387–405
Sung SS, Black CC, Kormanik TL, Zarnoch SJ, Kormanik PP, Counce PA (1997) Fall nitrogen fertilization and the biology of Pinus taeda seedling development. Can J For Res 27:406–1412
Timmer VR (1997) Exponential nutrient loading: a new fertilization technique to improve seedling performance on competitive sites. New For 13(1–3):279–299
Timmer VR, Armstrong G (1987) Growth and nutrition of containerized Pinus resinosa at exponentially increasing nutrient additions. Can J For Res 17:644–647
Timmer VR, Parton WJ (1984) Optimum nutrient levels in container growing medium determined by a saturated aqueous extract. Commun Soil Sci Plant Anal 15:607–618
van den Driessche R (1985) Late-season fertilization, mineral nutrient reserves, and retranslocation in planted Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) seedlings. Forest Sci 31:485–496
Wang Z, Zhang SY (1992) Larch forests in China. Forestry Publication House in China, Beijing, pp 185–186 In Chinese
Acknowledgments
This study was funded by the Fundamental Research Funds for the Central Universities (Contract TD2011-08, JD2011-3) and the USDA Forest Service Rocky Mountain Research Station and the National Center for Reforestation, Nurseries, and Genetics Resources. We sincerely appreciate assistance from Dr. John D. Marshall and Margaret Ward at the University of Idaho Stable Isotope Laboratory; Dr. Deborah Page-Dumroese and Joanne Tirocke at the USDA Forest Service, Rocky Mountain Research Station. We especially thank Associate Editor Dr. Barbara J. Hawkins for her efficient work and valuable suggestions, Editor-in-Chief Dr. Douglass F. Jacobs for the insightful comments, and the anonymous reviewers for valuable comments that improved this manuscript.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Zhu, Y., Dumroese, R.K., Pinto, J.R. et al. Fall fertilization enhanced nitrogen storage and translocation in Larix olgensis seedlings. New Forests 44, 849–861 (2013). https://doi.org/10.1007/s11056-013-9370-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-013-9370-z