
Abstract
Panama has the highest rate of change in the area of primary forests within Central America. However, to meet growing timber demands, it became popular over the last decades to establish plantations made up of foreign species such as Tectona grandis or Pinus spp. In the majority of the cases the species used are well known; their characteristics such as growth performance have been reviewed intensively and can be accessed in numerous publications. Characteristics of Panama’s native tree species of commercial relevance such as Hieronyma alchorneoides, Swietenia macrophylla and Terminalia amazonia are largely unknown and have been investigated within the study at hand. Using valuation methods of financial mathematics, the competitive position of these three indigenous species was assessed, the results compared to those of T. grandis stands in the same area. Land costs and taxes were not considered, as they would be the same for all species. Financial estimates for indigenous species will enlarge their acceptance for use in reforestation and plantation projects. Using the NPV method and applying the standard scenario, the profitability of T. grandis is lower than that of T. amazonia and S. macrophylla and lies only slightly above the profitability calculated for H. alchorneoides. This result clearly indicates that the investigated native tree species are comparable with T. grandis regarding their economic profitability. Besides its ecological impact, growing native tree species is now also economically legitimate. By calculating land expectation values for all tree species, ideal rotation lengths could be determined. For these species, considerable flexibility exists regarding the optimal rotation length.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the countries of Central America, traditional culling of tropical timber from primary forests is expected to come to a standstill within the next years, due to the depletion of remaining stands (Solorzano-Soto 1995). Especially in Latin America and the Caribbean, featuring high population densities, the timber trade forms an important economic factor and an important source of income for the rural population, just as it is in temperate regions. Growing difficulties of timber supply as well as increasing demand for land have led lead to rapid deforestation in these countries, according to FAO (2005).
As the country with the highest rates of change within its existing primary forest the study at hand subjects Panama to an exemplary closer inspection:
In Panama there are 1.2 Mha of land suitable and available for plantation establishment (Boyd 1998). Because of ongoing deforestation, this area is expected to increase. In 1990, Panama, with a total land surface of 7.5 Mha, still had 3.7 Mha of primary forests. In 2005, only 3.0 Mha were left. According to the FAO (Food and Agriculture Organisation of the United States) (2005) the annual deforestation rate is 1.23%. Accordingly, Panama’s primary forests are heavily declining. To protect the remaining tropical primary forests, mainly two alternatives are discussed:
-
(1)
Establish plantation forests (Günter et al. 2008; Cubbage et al. 2007; Pandey and Ball 1998; Lamprecht 1989),
-
(2)
Sustainably manage remaining natural forests (Günter et al. 2008; Cubbage et al. 2007; Finegan 1992; Quesada Mateo 1990).
The driving force for all activities in forest management, environmental protection and investment is cost-effectiveness. Comparing the management of natural forests with plantations financially, plantations often seem to be the better option. Natural forests tend to show a lower productivity, which leads to a lower cost-effectiveness, while plantations tend to show a higher cost-effectiveness (Cubbage et al. 2007). The establishment of a plantation however, also requires substantial initial investment. Alternative financing models can be a solution. Investment opportunities that offer competitive rates of return while also showing low risks are in demand. With growing environmental awareness, considerations of long term stability and real investment versus pure monetary values come to the forefront more and more. Many large-scale investors already are aware of this situation. The UBS AG (Union Bank of Switzerland), the world’s largest asset manager, also is one of the largest forest owners worldwide (Kollmansberger 2006). In spite of growing interest in non market values, so called Ecosystem Services, the return of investment still ranks first for investors. Estimates of rates of return are often based on optimistic assumptions to spark interest in forest plantations as an investment opportunity. Furthermore, in the majority of cases, operations are limited to monocultures of exotic species. Often, even for timber of indigenous tree species with a market, no published financial optimization in terms of forest management exist (Nichols et al. 2006).
In Panama, Tectona grandis is frequently planted (Simmons et al. 2002). The following hypothesis is therefore to be tested with a particular focus on this tree species for Panama:
H 1 :
The profitability of Teak plantations cannot be equalled by planting native tree species.
Indigenous tree species have some advantages compared to fast-growing exotic species. Native species are adapted to the site conditions and are therefore also more tolerant towards natural risks, while achieving comparable growth rates (Piotto et al. 2004a). Many native species can be grown on a broader scale of sites than exotic species. All these features may lead to a competitive edge for the less well known native species over the exotic Tectona grandis. But there are also risks in growing native species. Besides testing the hypothesis above on the basis of the available data, possible risks as well as chances related to growing indigenous tree species are to be discussed. In addition to Tectona grandis being considered for the reforestation in the investigation area, indigenous species have also been used. Alongside the species Anacardium exelcium (Espavé), Cordia aliodora (Laurel), Xantoxilum sp. (Tachuelo), Sterculia apetala (Panamá), Calicophylum candidisimun (Madroño), Didimopanax morototori (Pava), Cedrela odorata (Cedro), Miconia gobulifera (Pipi), Cassia mochata (Cañafistula) and Byrsonima crasifolia (Nance), that mainly answer the purpose of enriching biodiversity, are these three indigenous species of commercial value: Hieronyma alchorneoides, Swietenia macrophylla and Terminalia amazonia.
Many studies also show that by varying rotation lengths, the productivity of forest plantations can be considerably improved (e.g. Brazee and Mendelsohn 1988). Using the data available for the study at hand, a second hypothesis is therefore to be verified:
H 2 :
By varying rotation lengths, financial productivity can be increased.
Valuing plantations of native forest species
Establishment of forest plantations generally increases the contribution of forestry to the national economy (Alam et al. 2010). By establishing plantation forests on degraded areas that are abandoned such as former cattle ground, timber can be provided for the market and the impact of exploitation of natural forests can be mitigated. Furthermore, environmental conditions in the country are positively influenced and therefore upgraded (Gutierrez and Diaz 1999).
In the year 1990 forest plantations covered 10,000 ha of Panama’s land area, of which 7,000 ha where established using Pinus caribaea (INRENARE (Institute for the Management of Renewable Natural Resources) 1990). In 1997 plantations already covered around 30,000 ha, of which 14,000 ha consisted of Tectona grandis and another 10,500 ha of Pinus caribaea both in monoculture. But choosing different tree species for the species composition in a forest plantation project is an important silvicultural instrument that will later heavily influence the susceptibility for risks. By homogenising ecosystems to gain short term benefits, negative effects on biodiversity are generated, which often lead to a diminished financial robustness of the ecosystem, especially if natural risks exist (Knoke 2008). In this connection, Knoke and Hahn (2007, p. 312) assert:
(···) against the background of different production risks of tree species, comparison with a portfolio of shares stands to reason. In a forest stand that is made up of various tree species, effects that are subsumed as «diversification of risk» or «balancing of portfolio» may occur (···)
Planting a combination of tree species that are adapted to site conditions therefore is useful for ecological and financial aspects as well as for mitigating risks. But to fully embrace portfolio aspects in mixed species stands, prospective indigenous species have to be economically assessed.
Existing literature about the economy of plantation forestry in the tropics concentrates on economically productive species, most usually exotics, such as Pinus sp., T. grandis or Eukalyptus sp. (Cubbage et al. 2007). Tectona grandis is a well known species, whose characteristics such as growth performance have been reviewed intensively and can be accessed in numerous publications. But the characteristics of Panama’s native tree species of commercial relevance like Hieronyma alchorneoides, Swietenia macrophylla and Terminalia amazonia, are largely unknown. Regarding native species, we can at the most and if anything at all, find information about growth behaviour (Lamb 1966; Piotto et al. 2004b; Redondo-Brenes and Montagnini 2006). A comprehensive case study that measures mixed stands and projects growth performance over entire production periods and delivers an economic evaluation is initially provided by the present study.
Materials and methods
The study area
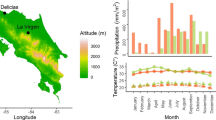
The study was carried out on the Pacific coast of the Central American republic of Panama. In Las Lajas, Province of Chiriquí (81°53’ W, 8°15’ N) at an elevation of about 8–50 m above sea level. The region is part of the tropics with an average annual precipitation of 3,000–3,500 mm and average annual temperature of 26.7°C. The 3–4 month dry season lasts from January to April (Worldwide Bioclimatic Classification System 1996–2009).
In 1995 the first plantation was established. It was planted on 23.5 ha former cattle ground. In 1996 and 1999 the plantation was complemented by adjacent and nearby areas, leading to a total plantation area of about 100 ha managed under the same concept.
The history of the research site being former cattle ground makes the site an ideal example, as equivalent sites are typical for future reforestation.
The main criteria for allocation of the different tree species within the area were the pre-conditions, such as varying soil conditions, of each microsite. Experience in managing the species as well as growth potential and commercial relevance of the species were taken into consideration just as well. The tree species were planted in different mixtures. Areas in which valuable tree species are discretely admixed are spread over the area, tessellated to gain a small-area mixture.
Before planting, the pre-existing vegetation was cleared. Remaining long-standing or valuable trees are mainly of the species Cassia grandis, C. moschata, Enterolobium cyclocarpum and Byrsonima crassifolia. According to the existing laws, 15 m adjacent to river banks were left unplanted. On these riparian strips and in other areas that are inappropriate for plantation establishment, like gullies, natural vegetation was kept. These areas are thus considered designated sanctuaries (Fig. 1).

Plantation layout at the study site
The seeds were obtained regarding to the provenance recommendations of CATIE (Centro Agronómico Tropical de Investigación y Enseñanza) in Costa Rica. For both Tectona grandis (Teak) and Hieronyma alchorneoides (Zapatero) the provenience “Pérez Zeledon” was suitable, delivered by the seed trader “Coopeagri”. For the species Swietenia macrophylla (Caoba) a Colombian provenience sold by “Semicol” was chosen, for Terminalia amazonia (Amarillo) the Panamanian provenience “Carta Vieja”, sold by “Particular”. Suitable seeds were sown in a nursery close to the plantation area, the seedlings raised for 6 month to a year before being hand planted into a planting pit of 60 × 45 cm. The top soil that accrues during this process is mixed with an organic fertilizer and put back into the planting pit. For 3 years after planting, the organic fertilizer “Bokashi” is deployed around the seedlings. The components of this fertilizer are fermented bird faeces, calcium, rice pellets, saw dust and ash. Furthermore, during the first 6 years after planting, the site was kept free of weeds by manual cutting six times a year. In the following years, natural regeneration between the planted rows was only cut if it grows directly into the rows of trees.
However, the most important management technique is pruning. Pruning was done during dry season; initially after the trees have reached a height of 4 m, a dbh of at least 10 cm and a branch diameter of more than 3 cm. Branches were removed up to a height of 50% of individual tree height. Thinnings were carried out at age 8 and 10; rotation period has been scheduled to 25 years.
The prescribed approaches to plantation establishment as well as management comply with the actions proposed by Lamb (1998) for the protection of biodiversity in plantations. The reforestation is an approach to sustainable and commercial plantation operation. Therefore, the plantation offers a suitable research area, to evaluate effects of ecologically managed plantations towards the potential of natural regeneration. A study regarding this has been taken in the form of a thesis by Paul (2008).
Financial analysis
To evaluate the economic performance of Tectona grandis as well as of the three native species of financial importance, Hieronyma alchorneoides (Zapatero), Swietenia macrophylla (Caoba) and Terminalia amazonia (Amarillo), all management activities from plantation establishment until final harvest have to be collected and financially valued. Costs for purchase of land and taxes were omitted as they would not affect the decision as to choice of species. All expenses arising for plantation establishment and forest management activities were documented by the plantation management and have been used as data basis for all subsequent calculations. Additionally, all expenses arising for the thinning taken out on the plantation in 2005 have been documented and are used in the following calculations. The income gained by timber sales had to be reconstructed.
Costs for plantation establishment
For plantation establishment, the first step was to prepare the site. In year 0 therefore expenses arise for clearing pre-existing vegetation, digging out planting pits, as well as for buying seeds and necessary tools etc. In the following years, expenses arise from management actions like pruning, weed-control, tools and material, fertiliser and herbicides. The necessary management actions were comprehensively described in the section titled “The Study Area” and have been accounted for by the plantation management. An overview of these expenses arising from year 0 to 5 after plantation establishment can be found in the following Table 1.
For the following years 6–25, for which no expenses were documented, an average of the previous years was used for valuation. Following common conventions, annual inflation rates have been disregarded (cp. Sagl 1995) rather all valuations used actual costs and prices.
Costs for thinning and final harvest
All expenses arising from the thinning in 2005 have been documented in detail. For the first thinning, accomplished in 2003, this information was not available. Therefore its financial data was calculated upon the assumption that the expenses are equal to the costs of the later thinning, as was done for annual spending for management activities and material above. Particular expenses arose for activities to keep the chain of custody complete and to later assign the harvested timber to certain parcels. In detail, these are hours of work for applying number tags as well as material costs for the tags themselves. Furthermore, the costs of forwarding the logs three kilometres to the wood yard by tractor are included. Total expenses for the thinnings taken out at age 8 and age 10 are made up of costs for transporting timber to the wood yard, arising costs for harvesting by chainsaw, costs for safety equipment, material and tools, as well as costs for general workings, that form the largest entry. In consideration of the fact that the thinnings are not commercial thinnings in the classical meaning, the expenses per hectare of around US $300 are relatively high.
Expenses for working time during final harvest at age 25 are calculated using the formula developed by Cruz Madariaga (2003):
T = Working time for logging [sec.], dbh = diameter at breast height [cm].
Using the diameter at breast height of the mean basal area tree at age 25 and the number of trees per hectare the working time for logging is calculated. The expenses for wages are set at US $10.14 per day (8 working hours), following average wages paid by the plantation management in 2006 including social security. Additionally, the costs for skidding and material according to the costs arising during thinnings are considered. Total costs for final harvest operations at age 25 are around US $1,300/ha, whereas the costs slightly vary between the tree species due to differences in timber dimensions and numbers of trees per hectare.
Income gained by timber sales
In Panama, the timber price realised for teak logs in 2007 was, according to the timber market report of ITTO, close to the worldwide midrange. For the other three tree species that are grown in the research area no timber market data was available. The timber obtained during thinnings in the research area could be sold irrespective of the species for a uniform price of US $200/m³ off the wood yard. Considering the growing demand for certified tropical timber and the possible development of market premiums paid for it (Kollert and Lagan 2006), it is assumed that in the following years equal prices will be achieved for all tree species investigated in this study. This view is also supported by personal communication with employees of the plantation management. In view of growing diameters and strong dimensional effects, future timber sales revenues (Table 2) are assumed, according to ITTO timber market reports (ITTO Tropical Timber Market 1998–2010).
It is assumed, that logs of stem wood of all four species, according to the assortment of the thinning in former years, can be sold at one standard price. This assumption is fortified by the comparable physical characteristics of the timber of all four species, shown in Table 3.
Furthermore, a 60% stem wood proportion out of total timber harvested is assumed. For fuel wood a net price of zero US $ is set. This means that the costs for primary conversion of fuel wood are just covered by the attainable revenue for this particular assortment.
Yield projections
The information on growth performance presented below provides the basis for all economic calculations. For the inventories taken out annually by the plantation management, permanent monitoring plots for all tree species were established. Each plot measures 20 × 20 m, and is located within the plantation by random selection. In total there are 16 plots of 400 m² each. Because the spacing between trees is different for each species, the number of trees per plot is between 10 and 48 individuals. The measured data was later converted to analogous values for hectares.
To evaluate the economic performance of T. grandis and S. macrophylla data from the permanent inventory plots, yield tables and information from literature was used. Both tree species have frequent appearances in the literature. Because of this advantage, a rather high validity of the results can be assumed. For H. alchorneoides and T. amazonia only scarce additional information regarding their growth was available from literature (Piotto et al. 2003, Redondo-Brenes and Montagnini 2006). Regarding the reliability of the results it has to be noted that at the research site all four tree species investigated were not planted randomly, but according to their site requirements after a soil sampling was carried out. This leads to systematic differences in site qualities between the tree species. For the growth behavior of the tree species in reality, slight divergences therefore have to be anticipated, that are covered by the optimistic and pessimistic scenarios.
The first thinning was carried out at age 8. During this thinning the number of trees per hectare of each species fell by an amount related to the number of trees planted. In T. grandis stands 41% of the trees were cut, for H. alchorneoides, the number of trees per hectare fell off by an average of 22%, the average number of S. macrophylla trees per hectare was reduced by 6% and in T. amazonia stands 40% of the trees were cut.
During the second thinning at age 10 another 14% of T. grandis trees, 4% of H. alchorneoides, 2% of S. macrophylla and 25% of T. amazonia were harvested. A detailed overview over yield surveys taken out at the study site and expected future growth can be found in the appendix (Tables 6, 7, 8, and 9).
To classify growth performance at the study site and to extrapolate it into the future beginning with age 11, appropriate yield tables were analyzed. For T. grandis, from comparative data by Pérez and Kanninen (2005), expected diameter and height growth at the study site as well as a form factor of 0.45 for round wood calculation was derived.
To forecast further development of growth of the natural forest species H. alchorneoides, we had to revert to a study by Redondo-Brenes and Montagnini (2006) and another study by Piotto et al. (2003). Both studies deal with growth performance of the species H. alchorneoides and T. amazonia. It is assumed that increments at the study site evolutes comparably. To avoid growth prognoses becoming too optimistic, the lower increments for tree height and diameter determined in each of the two studies were used. For calculating the volume of merchantable H. alchorneoides timber, a form factor of 0.45 enters into the calculation. This form factor is according to that used for internal calculations by the management and emanates empirical value (Camacho 2008).Footnote 1
According to the approach used for the yield estimation of T. grandis, for S. macrophylla yield tables from comparable sites were consulted to classify growth performance. For calculating the stem volume of S. macrophylla a form factor of 0.65 was used, as determined by Mora-Chacón et al. (2002).
To project growth performance of T. amazonia according to the approach used for H. alchorneoides results gained by Redondo-Brenes and Montagnini (2006) and Piotto et al. (2003) were used. An overestimate of the growth performance is obviated by using the lower increments determined by Redondo-Brenes and Montagnini (2006). A form factor of 0.45 was used, also taken from the calculations used by the forest enterprise and is based on the experience of the on-site forest engineers (Camacho 2008).
Comparative analysis of yield
For years 11–25 for all tree species no further thinnings were planned. The stem number determined for each species in the research area takes into account an annual mortality of 0.5% of the standing trees per hectare in years 11–25 (Camacho 2008). Comprehensively a loss of 20% of the commercial volume is considered, to factor in bark and losses during harvesting to calculate felling value (Camacho 2008). As mentioned before, all potentially commercial timber (80% of standing timber over bark) is then split into 60% stem wood and 40% fuel wood.
In the research area, Tectona grandis—compared with Hieronyma alchorneoides, Swietenia macrophylla and Terminalia amazonia—gained the highest growth performance until age 10. With advancing age, T. amazonia and S. macrophylla turn out to be more productive. For S. macrophylla this traces back to the advantageous stem form and higher form factor arising from it. Whether it is at all possible to grow S. macrophylla in plantations for equivalent rotation lengths continues to be debated. In the literature, many reports about failures in S. macrophylla stands starting with age 10 exist. In the discussion section of this paper, this problem will be taken up again in detail. For this work, it will be assumed that the species will reach age 25.
Terminalia amazonia displays high increment in other studies as well (Piotto et al. 2003; Redondo-Brenes and Montagnini 2006) a very good growth performance is strongly related to the site quality though (Calvo-Alvarado et al. 2007). A comparative overview is given in Fig. 2.

Total increment of the four tree species in the research area
Ranking method
For Panama, Benitez et al. (2007) estimated risk adjusted discount rates of around 9.9%. International investors aiming at forest investments to diversify existing portfolios come to a much lower receivable interest. Private landowners have very individual receivable interest. As current asset portfolios of private landowners are unknown, a range is necessary to display all possible situations. In South American Tectona grandis plantations, Cubbage et al. (2007) assessed rates of return of 5–13%. These values were used as an indication for general assumptions regarding the profitability of plantations as well as to allow general assessment of forest plantations.
Net present value method
The economic performance of the different species is described with an indicator that is derived from capital budgeting. The net present value (NPV) method is a discounted cash-flow method and counts among the methods of capital budgeting. By discounting payments that are made at different points in time to the start date of an investment, payments made at different moments become comparable using the NPV method. The NPV of an investment is the sum of all present values, thus all cash flows, both inflows and outflows of cash that are caused by the investment. It differentiates between absolutely profitable investments, with a NPV greater than or equal to zero and relatively profitable investments with a NPV greater than or equal to the NPV of an alternative investment. The first case answers the question whether or not an investment generally is to be made; the second case answers the question of which of the alternatives is more profitable. It is the latter case that concerns us here.
For calculating the NPVs the following equation according to Thommen and Achleitner (2009) is used:
NPV0 = Net present value, T = years between stand establishment and final harvest, I n = Cash inflow in year n, O n = Cash outflow in year n, i = discount rate (in hundredth), n = time after stand establishment in years.
The terms cash inflow and cash outflow that are common in financial mathematics correspond with the terms incoming payments and out payments. To calculate the NPV an interest factor or discount rate i is needed. i can be the rate of return required by an investor, the rate of return that can be achieved with an alternative capital asset, or it can be chosen as a target rate of return, including all influencing factors. In many cases i is chosen as the rate of return an alternative investment offers. Because of the controversial opinion regarding suitable interest rates for forest enterprises and forests, for calculating the NPV, interest rates between 0 and 15% have been used in this study.
Considering uncertainties and optimization
Uncertainty and sensitivity analysis
Damages to a forest stand which can result from insect outbreaks, volatile timber markets and other factors influence possible revenues. Also, it is impossible to forecast all relevant factors. For the paper at hand, the future risk situation in form of environmental circumstances or damages to the stand is deemed to be unknown by the decision maker. One way to factor these risks into the analysis is to perform a sensitivity analysis.
Within a sensitivity analysis, the marginal values of the results are elaborated. One or more parameters influencing the investment are changed (Heidingsfelder and Knoke 2004). It is tested at what point the target return values are affected (for example the NPV). By this approach, the sensitivity of the investment towards the change of influencing values like timber market development in form of timber prices, total increment, and others can be tested systematically. For this purpose, various input parameters were changed by posing a range of alternatives for the incoming payments.
Varying rotation lengths
The land expectation value (LEV) represents the present value of annuity of a perennial periodic annuity made up of the sum of the future felling value and all cash inflows compounded to the moment of final harvest, minus all compounded cash outflows for tending, material and other costs (Faustmann 1849). The sum is then diminished by the perennial annuity of administrative expenses. With the LEV it is possible to consider effects of different durations of investments, such as rotation, which is one of the most important control factors in forest management. Even for well investigated species, the optimal rotation length is still being discussed. For many tropical species a rotation period of 25 years is assumed to be ideal (Evans and Turnbull 2004). By investigating the development of LEVs under varying rotation lengths, production periods can be optimised. Furthermore the calculation of LEVs is necessary to assure that longer rotation periods are not estimated too optimistically, as well as to achieve comparability with other studies. Therefore the LEVs of all four investigated tree species were calculated for rotation periods between 10 and 30 years and a regression analysis was carried out with the ascertained values to smooth the curves relative to time.
Results
Results of the ranking methods
The description of the valuation results first goes into cash in- and outflows as well as felling values and other performance indicators. In a first step, a situation without uncertainty is implied.
Cash inflow and outflow
The observations made in sections “Income gained by timber sales” and “Yield projections” result in annual cash in- and outflows of varying amounts for Tectona grandis as shown in Fig. 3. The cash outflows are displayed as hanging columns because of their negative algebraic sign. The cash inflows from thinnings in year 8 and 10, as well as cash inflows from final harvest in year 25 are displayed as standing columns. In all other years, no cash inflows are being set.

Cash in- and out-flows Tectona grandis
The cash outflows for the tree species H. alchorneoides, S. macrophylla and T. amazonia only marginally differ from the cash outflows for T. grandis and are therefore not displayed in detail.
Net present value method
Considering the change of the NPV depending on the required rate of return, the criteria of relative profitability introduced in section “Net preent value method” is taken as basis. Figure 4 illustrates how the NPV for all four tree species changes depending on the discount rate used.

Net present value function of all four tree species
At a required discount rate of 0%, the NPV is equivalent to the total sum of all in- and outflows. The graph of the NPV always intersects with the axis of the discount rate at the internal rate of return. In the example displayed in Fig. 4, the tree species achieve internal rates of return of up to 15%. The internal rates of return for the investigated tree species appear in italics in Table 4.
Furthermore, the dependency of the required final stand value in year 25 and the internal rate of return can be displayed as in the following graph (Fig. 5).

Felling value year 25 necessary for a NPV = 0
The graph displays the felling value the investment must attain in year 25 to still reach a NPV of 0 and therefore to still be considered profitable. This graph can be used as a simple tool to assess the profitability of an investment, if a rough idea of the future felling values, that also holds true in practice, exists.
Because cash outflows are equivalent for all tree species, H. alchorneoides, S. macrophylla and T. amazonia follow as for the displayed species T. grandis.
For a required rate of return of 15% a felling value of around US $120.000/ha would be necessary to avoid a negative NPV. Therefore, such a high internal rate of return appears to be unlikely.
Considering uncertainties: a sensitivity analysis of net present value and internal rate of return
The sensitivity analysis gives an overview of the reactions of the NPVs and internal rates of return on changes in the input variables.
Varying initial cash outflow
A change in the initial cash outflow—meaning the costs of stand establishment—at the beginning of the period under consideration results in a parallel downwards shift of the NPV graph for all tree species. Any such change can for example evolve from rising costs for seedling or rising wages. If the initial cash outflow rises for a certain amount, the NPV declines by the same amount, irrespective of the used rate of return, and vice versa.
This effect is originated by using a discount rate (1 + i)0 at the beginning of the period under consideration. The parallel shift furthermore results in a change of the point of intersection of the graph with the X-axis is moving to the left, which means that the internal rate of return declines as shown by the example of T. grandis in Fig. 6.

Changes in the graph of the net present value function with rising initial cash outflows for stand establishment of T. grandis
The changes in the payments made for stand establishment by US $2,500, and US $5,000, respectively, are deliberately chosen to be very high to clearly point out the parallel shift of the graph. For the sensitivity analysis itself, additional 25, 50, 75, and 100% are charged for the initial cash outflow. A possible decline of the costs for stand establishment is not being considered, as an initial cash outflow of around US $1,000/ha including all plants is already set rather low. Detailed results for all four species investigated can be found in the appendix, Tables 10, 11, 12, and 13.
Even for changes of up to 100%, the internal rate of return for all tree species only varies in a range of less than 1%. Even a considerable increase of the costs for stand establishment has only very little influence on the NPVs. Furthermore it is to be tested how the NPV changes over all requested rates of return if the cash inflow earned by final harvest varies.
Varying cash inflow from final harvest
Varying earnings from final harvest can for example be caused by declining timber prices. Also, lower increments—for example caused by changing environmental conditions—that lead to lower dimensions in the produced timber and lower standing volumes, can influence the earnings gained during final harvest. Vice versa, timber prices, saw wood proportions and many other factors can also change in a positive way. To asses financial consequences of all possible future developments, the cash inflow by final harvest in year 25 for the four tree species considered was varied from −50% through +50%, resulting in a change of the NPVs. The possible range of the NPV at higher, and lower incomes gained by final harvest is shown for T. grandis in Fig. 7.

Changes in the graph of the net present value function with rising and declining cash inflows from final harvest, T. grandis
The NPV at lower cash inflows from final harvest is below the NPV of the standard scenario. Likewise, the NPV gained with a higher final cash inflow as a basis are above the standard scenario. However, in both cases the difference between the standard scenario and the other scenarios becomes smaller and approaches zero with rising discount rate. This is due to the fact that the discount factor used for the cash inflow after final harvest ((1 + i)−25) quickly becomes very small due to rising interest rates and the discounting period of 25 years,.
Pessimistic and optimistic combinations
Within a framework of pessimistic and optimistic combinations, the paper at hand investigates the fluctuation margin of the NPVs for the investigated tree species, if rising cash outflows for stand establishment occur along with sinking cash inflows after final harvest and vice versa. To consider a pessimistic development of the NPV, initial cash outflows are increased by 50% whilst simultaneously decreasing the cash inflow after final harvest by 50%. To consider an optimistic development of the NPV, decreased cash outflows for stand establishment by 50% are combined with 50% higher cash inflows gained by final harvest. The results for all four tree species can be seen in Table 5.
If the internal rate of return is considered, the following ranking results: If a pessimistic development occurs, T. amazonia still reaches the highest internal rates of return. Already at a slight increase of the cash inflows, T. amazonia can compete with the results of the other three species in the situation of an optimistic development.
T. grandis turns out to be least susceptible to the alternative scenarios, whilst H. alchorneoides in a pessimistic case scores lowest and is therefore rather susceptible towards changes.
Varying rotation lengths
By calculating LEVs under varying rotation lengths, production periods can be optimized. Therefore the LEVs of all four investigated tree species are calculated for rotation periods between 10 and 30 years and a regression analysis was carried out with the ascertained values.
The development of the LEV of T. grandis is exemplary displayed in Fig. 8, the graphs for the three native species can be found in the appendix (Figs. 9, 10, and 11). Whereas the smoothed curve is pictured as a bold line, the “real” values are pictured as a dashed line. The partially remarkable leaps of the LEV graphs originate from the variation in incomes from timber due sales due to increasing log sizes.

LEV development for T. grandis under varying rotation lengths
For all four species a rotation period of 25 years turns out to be ideal at a discount rate of 3%. At discount rates larger than 3%, the ideal rotation decreases, but the differences of the LEVs with age however are comparatively low. Overall, the trend line added to Fig. 8 clearly shows the bell-shaped development which is typical for corresponding graphs, whereas the assumed optimal rotation length of 25 years is close to the maximum value at all times. The explicit increase of the LEVs between a 22 year and a 23 year rotation length rests on the rising incoming payments for timber sales due to larger dimensions. Furthermore, the flat run of the graph that occurs in all four species points out a wide range of management options regarding ideal market situations for harvesting operations.
Discussion
Reassessing the hypothesis
Calculating the net present values (NPV) for a standard scenario at a rotation length of 25 years led to the result that T. grandis reaches internal rates of return of approximately 11–12%. The natural forest species H. alchorneoides reaches values of 10–11%, S. macrophylla reaches internal rates of return of approximately 12–13% and the third natural forest species T. amazonia even reaches internal rates of return of >15%. The calculations of the NPVs were carried out for the standard scenario without considering the costs for purchase of land, costs for maintaining an administration, taxes or a distribution system. If all these costs are considered, the results will shift towards the results of the pessimistic development scenario. For plantations growing selected indigenous tree species, Cubbage et al. (2007) calculated rates of return between 5 and 13%. An equivalent result is aimed for at the research site mixing the four species T. grandis, H. alchorneoides, S. macrophylla and T. amazonia.
The rates of return calculated here do turn out to be comparatively low. As predictions regarding timber markets at the rotation age are difficult to make, it has to be mentioned that with increasing volumes of wood becoming available from maturing plantations (Clark 2001), price expectations may not necessarily be realized, even though a market premium for certified logs may be fetched in the future (Kollert and Lagan 2006). The pessimistic scenarios considered here however, display future performances at price declines of up to 50%.
Furthermore there exist large uncertainties regarding the survival probability for the natural forest species. As mentioned before, especially for S. macrophylla there are indications that a rotation period of 25 years in a plantation cannot be assumed offhand. One of the largest known problems with S. macrophylla in plantations are calamities of the insect pest Hypsipyla spp. which can be found in all of Central America (Mayhew and Newton 1998). An infestation results in a damage of the main shoot and therefore forked trees and a considerable decrease of increments. At the research site, single appearances of H. grandella were observed during inventories.
On our assumtions, native species appear competitive with T. grandis in this area. The comparison of the NPVs shows that the profitability of growing T. grandis is below the profitability of T. amazonia as well as S. macrophylla and only slightly higher than the profitability of growing H. alchorneoides if the standard scenario is used. However, for S. macrophylla high uncertainties regarding future outcomes have to be considered, leading to the assumption that results will strongly tend towards the pessimistic scenario if the stands of this species are able to reach the rotation age. Furthermore, the timber may turn out to be mainly sapwood, unless a much longer rotation is used. These uncertainties also raise the question if S. macrophylla should be planted at all. If trees of this species are planted in small groups or as single tree admixtures the risk of insect losses can be minimized. If the trees of S. macrophylla should then fail to reach rotation age, surrounding trees of other species will be able to fill in the gaps. We therefore recommend understanding the admixture of S. macrophylla as an investment that offers certain chances at a high risk and should therefore be planted in proportions smaller than 5% only.
For H. alchorneoides it is likely that the revenues from timber sales will in reality turn out to be higher than the revenues used in the paper at hand, as according to Piotto et al. (2003), an increasing demand for saw timber of this species is to be expected. In this case, the small advantage of T. grandis towards H. alchorneoides would wear off in the occurrence of an according scenario. Our hypothesis—the profitability of Teak plantations cannot be equalled by planting native tree species—is therefore refuted, provided that all assumptions turn out to be true in the future.
Conclusions
This paper was able to show how the investigated species behave regarding their financial performance. To our knowledge it is the first paper delivering actual data for financial comparison of the three investigated native species with Tectona grandis, as so far very little is known about the financial competitiveness of natives. Regarding the NPV of all four tree species, the profitability of T. grandis drops below the profitability of T. amazonia and S. macrophylla applying the standard scenario already and lies only slightly above the profitability calculated for H. alchorneoides. This result clearly indicates that the investigated native tree species are comparable with T. grandis regarding their economic profitability assuming that in the future the wood can be sold at comparable prices. Besides its ecological impact (Evans and Turnbull 2004; Hartley 2002) growing native tree species now also obtains economic legitimacy and should therefore be considered to be used alongside Tectona grandis in plantation establishments. Reconciliation between ecology and economics is made possible as the only obstacle so far was the lack of knowledge existing in this field of research.
An increase in the revenues from final harvest in year 25 has a great influence on the total profitability of the investment. Alongside the timber prices, which cannot be influenced by small enterprises, especially quality and dimension of the grown stem wood plays an important role for increasing revenues. Special attention should therefore be paid to the tending of such timber.
The information provided here makes a substantial contribution to the acceptance of native tree species within commercial forestry in Central America. The information regarding their profitability and growth also provides a basis for further calculations. However, it has to be noted that the extrapolation of growth from year 10 to 25 is very long and therefore holds uncertainties. For more reliable results, future evaluation of the profitability and economy of the investigated species to ages beyond age 10 has to be carried out. By integrating risk, as done for by Heidingsfelder and Knoke (2004) in a comparable study, the insecurity of an investment decision can be quantified. Such risk integration can for example consider the uncertainty regarding the future revenues gained by future timber sales. Furthermore, the tendency to risk taking by the decision maker can be integrated into evaluating the profitability of the investment. The relevance of integrating risks was determined by Knoke and Wurm (2006). Assessments of the diversification that results by growing the four different tree species would be reasonable as well. Diversification effects were first described by Markowitz (1952) in his Portfolio-Theory. An investment made up of a combination of different capital assets can—compared to a single investment—lower the risk at equivalent rates of return. Also, the rate of return can be positively influenced by combining different capital assets. Knoke et al. (2005) devolved this approach on forest economics, determining mixed species plantations to be equivalent to a mixed portfolio. They observed that the ecologic concept of “mixed forests” leads to considerable economic benefits. To assess the diversification effects for the four tree species investigated would further advance the clarification of the economy and productivity of T. grandis, H. alchorneoides, S. macrophylla and T. amazonia. To increase the accuracy of assessing such forest investments there is also further need for information about tree species interaction in mixed stands, according to the research carried out by Petit and Montagnini (2006).
Notes
Yaels Camacho, Forest engineer at the research site.
References
Alam M, Furukawa Y, Harada K (2010) Agroforestry as a sustainable landuse option in degraded tropical forests: a study from Bangladesh. Environ Dev Sustain 12:147–158
Benitez PC, McCallum I, Obersteiner M, Yamagata Y (2007) Global potential for carbon sequestration: Geographical distribution, country risk and policy implications. Ecol Econ 60:572–583
Boyd E (1998) A compilation of forest statistics for selected African and Latin American countries. Oxford Forestry Institute, United Kingdom
Brazee R, Mendelsohn R (1988) Timber harvesting with fluctuating prices. For Sci 34:359–372
Calvo-Alvarado JC, Arias D, Richter DD (2007) Early growth performance of native and introduced fast growing tree species in wet to sub-humid climates of the Southern region of Costa Rica. For Ecol Manag 242:227–235
Camacho Y (2008) Oral communication
Clark (now Ajani) J (2001) The global wood market, prices and plantation investment: an examination drawing on the Australiaan experience. Environ Conserv 23:58–64
Cruz Madariaga GE (2003) Untersuchungen zur Überführung von südchilenischen Nothofagus-Pumilio-Naturwäldern in naturnahe Wirtschaftswälder. Dissertation, Universität Freiburg
Cubbage F, Mac Donagh P, Sawinsk J, Rubilar R, Donoso P, Ferreira A, Hoeflich V, Morales Olmos V, Ferreira G, Balmelli G, Siry J, Noemi Báez M, Alvarez J (2007) Investment returns for selected plantation and native forests in South America and the Southern United States. New For 33:237–255
Evans J, Turnbull J (2004) Plantation forestry in the Tropics. 3. Ausgabe. Oxford University Press, New York
FAO (Food and Agriculture Organisation of the United States) (2005) Countries—forest facts by country (www.fao.org/forestry/site/18307/en) Rome
Faustmann M (1849) Calculation of the value which forestland and immature stands possess for forestry. J For Econ 1(1):7–44 (1995)
Finegan B (1992) The management potential of neotropical secondary lowland rain forest. For Ecol Manag 47:295–322
Günter S, Weber M, Aguirre N, Stimm B, Mosandl R (2008) Reforestation of abandoned pastures: silvicultural means to accelerate forest recovery and biodiversity. In: Beck E (ed) Gradients in a tropical mountain ecosystem of ecuador. Springer, Berlin, pp 431–441
Gutierrez RR, Diaz HI (1999) Memoria de las estadísticas de los recursos forestales de Panama. Workshop for Forest Resources Assessment Programme
Hartley M (2002) Rationale and methods for conserving biodiversity in plantation forests. For Ecol Manag 155:81–95
Heidingsfelder A, Knoke T (2004) Douglasie versus Fichte. Ein betriebswirtschaftlicher Leistungsvergleich auf der Grundlage des Provenienzversuches Kaiserslautern. J.D. Sauerländer Verlag, Frankfurt am Main
INRENARE (Institute for the Management of Renewable Natural Resources) (1990) Plan de accion forestal de Panama, Documento principal. Instituto Nacional de Recursos Naturales Renovables
ITTO Tropical Timber Market (TTM) Report (1998–2010) Output of the ITTO Market Information Service (MIS), Japan. http://www.itto.int
Knoke T (2008) Mixed forests and finance–Methodological approaches. Ecol Econ 65:590–601
Knoke T, Hahn A (2007) Baumartenvielfalt und Produktionsrisiken: Ein Forschungseinblick- und Ausblick. Schweiz Z Forstwesen 158:312–322
Knoke T, Wurm J (2006) Mixed forests and a flexible harvest policy: a problem for conventional risk analysis? Eur J For Res 125:303–315
Knoke T, Stimm B, Ammer C, Moog M (2005) Mixed forests reconsidered: a forest economics contribution on an ecological concept. For Ecol Manag 213:102–116
Kollert W, Lagan P (2006) Do certified tropical logs fetch a market premium? A comparative price analysis from Sabah, Malaysia. For Policy Econ 9:862–868
Kollmansberger P (2006) Forest investment in Panama – Long term investment with real values. (www.sustainablebusiness.com/progressiveinvestor, #39)
Lamb FB (1966) Mahagony of tropical america. It’s ecology and management. The University of Michigan Press, Ann Arbor, p p 220
Lamb D (1998) Large-scale ecological restoration of degraded tropical forest lands: the potential role of timber plantations. Restor Ecol 6:271–279
Lamprecht H (1989) Waldbau in den Tropen. Die tropischen Waldökosysteme und ihre Baumarten; Möglichkeiten und Methoden zu ihrer nachhaltigen Nutzung. Hamburg und Berlin, Verlag Paul Parey
Markowitz H (1952) Portfolio selection. J Finance 7:77–91
Mayhew JE, Newton AC (1998) The silviculture of mahogany. CABI Publishing, Oxon
Mora-Chacón FA, Gutierrez-Leitón M, Orozco C, Brenes, R (2002) Ecosistemas forestales de bosque seco tropical: investigaciones y resultados en Mesoamérica. Editorial Heredia, Universidad Nacional/INISEFOR, CR, pp 87–100
Nichols JD, Bristow M, Vanclay JK (2006) Mixed-species plantations: prospects and challenges. For Ecol Manag 233:383–390
Pandey D, Ball J (1998) The role of industrial plantations in future global fiber supplies. Unasylva (FAO) 49:37–43
Paul C (2008) Vergleichende Untersuchungen zur Diversität des Unterstandes in Teak- (Tectona grandis) und Mischplantagen einheimischer Baumarten in West-Panama. Diploma thesis, Technische Universität München, Germany
Pérez D, Kanninen M (2005) Effect of thinning on stem form and wood characteristics of teak (Tectona grandis) in a humid tropical site in Costa Rica. Silva Fennica 39:217–225
Petit B, Montagnini F (2006) Growth in pure and mixed plantations of tree species used in reforesting rural areas of the humid region of Costa Rica, Central America. For Ecol Manag 233:338–343
Piotto D, Montagnini F, Ugalde L, Kanninen M (2003) Performance of forest plantations in small and medium-sized farms in the Atlantic lowlands of Costa Rica. For Ecol Manag 175:195–204
Piotto D, Víquez E, Montagnini F, Kanninen M (2004a) Pure and mixed forest plantations with native species of the dry tropics of Costa Rica: a comparison of growth and productivity. For Ecol Manag 190:359–372
Piotto D, Montagnini F, Kanninen M, Ugalde L, Viquez E (2004b) Forest plantations in Costa Rica and Nicaragua: performance of species and preferences of farmers. J Sustain For 18(4):59–77
Posch B, Wegener G, Grosser D, Wagner L (2004) Physikalische und mechanische Untersuchungen an Teakholz (Tectona grandis L.f.) aus Plantagen in Panama. Eur J Wood Wood Prod 62:31–35
Quesada Mateo CA (1990) Estrategia de conservación para el desarrollo sostenible de Costa Rica. Ministerio de Recursos Naturales, Energía y Minas. Servicios Litográficos San José (Costa Rica)
Redondo-Brenes A, Montagnini F (2006) Growth, productivity, aboveground biomass, and carbon sequestration of pure and mixed native tree plantations in the Caribbean lowlands of Costa Rica. For Ecol Manag 232:168–178
Rijsjdijk JF, Laming PB (1994) Physical and related properties of 145 timbers. Kluwer, Dordrecht
Sagl W (1995) Bewertung in Forstbetrieben. Blackwell Wissenschaftsverlag. Wien, Berlin
Simmons C, Walker R, Wood C (2002) Tree planting by small producers in the tropics: a comparative study of Brazil and Panama. Agrofor Syst 56:89–105
Solorzano-Soto R (1995) Breve diagnóstico y soluciones de corto plazo al sector forestal. Notas Técnicas y Económicas 7: 10. Centro Científico Tropical, CR
Thommen J-P, Achleitner A-K (2009) Allgemeine Betriebswirtschaftslehre–Umfassende Einführung aus managementorientierter Sicht, 6th edn. Gabler, Wiesbaden
USDA Forest Service, Center For Wood Anatomy Research (2010) Technology transfer, Fact sheets. http://www.fpl.fs.fed.us/research/centers/woodanatomy/index.php
Worldwide Bioclimatic Classification System (1996–2009) S. Rivas-Martinez, S. Rivas-Saenz, Phytosociological Research Center, Spain. http://www.globalbioclimatics.org
Acknowledgments
The presented study is part of the project “Bioeconomic modelling and optimization of forest stands: Towards silvicultural economics” KN 586/7-1 funded by the German Research Foundation (DFG). The authors wish to thank Mrs. Kristin Dzurella for the manuscript language editing and three anonymous reviewers for valuable suggestions.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
See Tables 6, 7, 8, 9, 10, 11, 12, and 13; Figs. 9, 10, and 11.

LEV development for H. alchorneoides under varying rotation lengths

LEV development for S. macrophylla under varying rotation lengths

LEV development for T. amazonia under varying rotation lengths
Rights and permissions
About this article
Cite this article
Griess, V.C., Knoke, T. Can native tree species plantations in Panama compete with Teak plantations? An economic estimation. New Forests 41, 13–39 (2011). https://doi.org/10.1007/s11056-010-9207-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-010-9207-y