
Abstract
To determine the effect of shade on morphology, growth and biomass allocation in Picea sitchensis, Larix × eurolepis and Thuja plicata, seedlings were grown in the open or under shadehouses providing 25%, 50% and 75% reductions of full-light for two growing seasons. For most of the characteristics assessed there was no significant interaction between species and shade indicating that the morphological responses to changing shade treatments were not species-dependent. After two growing seasons the mean height increment for the three species was significantly greater in 25% (76.1 cm) and 50% shade (74.9 cm) than in the open (69.5 cm). Root collar diameter increment, shoot, root and total biomass declined significantly with increasing shade while the opposite was true for the height:diameter ratio. In both western red cedar and hybrid larch the shoot:root ratio was significantly greater in the shade while in Sitka spruce this characteristic was not influenced by shade. While all species had significantly greater specific shoot areas in 75% shade than in 0% shade, this trend was particularly pronounced in hybrid larch. In hybrid larch and western red cedar, the normalised specific projected shoot area increased significantly with increasing shade. The opposite trend was observed for Sitka spruce. We conclude that in the main the species studied demonstrated similar shade acclimation responses despite their reported differences in shade tolerance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In Ireland, the majority of forests are managed under the clearfell system. However, Ireland’s commitment to sustainable forest management and society’s desire for a broader range of forest management options has led to increased interest in continuous cover forestry and hence in alternative silvicultural systems to clearfelling. The alternative systems currently being considered in Ireland include the shelterwood systems and the group selection system. A common feature of these systems is that a young stand develops under the canopy of an older stand which influences the amount of light available to the understorey species. To date, the process of converting forests currently managed under the clearfell system in Ireland to continuous cover forestry has only commenced in a limited number of stands. One of the main difficulties that forest managers face in choosing which system to implement and what silvicultural management to subsequently undertake is the lack of information on how some of the commonly planted tree species in Ireland will grow in the shaded conditions that prevail in silvicultural systems that deliver continuous cover forestry.
The shade tolerance of a tree species is a function of a species’ ability to efficiently capture and use limiting light resources and thereby optimise the whole-plant carbon balance in shade (Khan et al. 2000). This is achieved through a combination of morphological and physiological adaptations. Thus tree species characterised as being shade-tolerant or light-demanding exhibit differing morphological and physiological responses along a light availability gradient (Chen et al. 1996) and the responses to shade may be indicative of how a species copes and survives as light becomes limiting (Van Hees and Clerkx 2003). Plants usually respond to decreasing light availability by reducing growth (Givnish 1988; Wang et al. 1994). Height increment, for example, decreases with shade, but the rate of decline is usually less in shade-intolerant species than in shade-tolerant species (Walters et al. 1993; Chen et al. 1996). This has been attributed in part to the tendency of some shade-tolerant species to sacrifice height growth in the shade in favour of branch elongation (Kimmins 1987). The strategy inherent in shade-intolerant species is to increase height growth in an attempt to out-compete neighbouring plants and improve their chance of light interception in the open. However, in the shade, this strategy is costly, as it requires more energy in an already light limited environment to maintain the increasing proportion of non-photosynthetic tissues (Messier et al. 1999; Claveau et al. 2002).
Specialisation for functioning in shade has also been reported to include an increase in shoot:root ratios (S:R ratios) and height:diameter ratios (H:D ratios), since as light decreases plants allocate more biomass to the above-ground structures especially leaves, to increase photosynthetic leaf area (Wang et al. 1994; Chen 1997; Van Hees and Clerkx 2003; Robakowski et al. 2004). Changes to the structure of leaves with changing light availability have also been noted. Many studies have found that shade-tolerant species exhibit a greater capacity to change leaf structure than shade-intolerant species (Chen et al. 1996). These changes in leaf structure can involve the development of greater leaf area per leaf dry mass (specific leaf area) by decreasing the number and size of the palisade cells (i.e. leaf thickness) and decreasing the leaf stomata density (Chen and Klinka 1996; Fownes and Harrington 2004; Robakowski et al. 2004).
The objective of this study was to examine under controlled conditions the impact of shade on morphology, growth and biomass allocation in Sitka spruce (Picea sitchensis (Bong.) Carr.), hybrid larch (Larix × eurolepis, Henry) and western red cedar (Thuja plicata, Donn ex D. Donn). In these controlled conditions seedlings of the three species were exposed to different shade levels but other resources such as water and nutrients were kept at almost the same levels for all treatments. In a forest understorey the response to changing light levels is influenced by the availability of other resources (Kimmins 1987). In this study the aim was to separate the effects of these confounding factors and focus only on the response to changing light conditions.
The species chosen for study are, or will be, among the conifer species most commonly planted in Ireland and they vary in their reported shade tolerance. In Ireland, the forest estate is dominated by Sitka spruce, which accounts for 60% of the annual afforestation programme. This species has been variably classed as light demanding (Savill 1992; Horgan et al. 2003); as intermediate in its light requirements (Mason et al. 1999) or as shade-tolerant (Wenger 1984). Hybrid larch, which is described as light demanding (Savill 1992; Wenger 1984; Mason et al. 1999; Horgan et al. 2003), has accounted for a significant component of the planting programme in recent years, where it is mainly used for landscape and amenity purposes. Western red cedar, which is considered to be shade-tolerant (Wenger 1984; Mason et al. 1999; Horgan et al. 2003), is currently only a minor species in Irish forestry but its importance is set to increase. These classifications are based, in most instances, on the traditional categorisation of tree species into shade tolerance classes based on the silvics of the species rather than on specific research trials evaluating the response of the species to varying shade levels. Such trials are few especially in the case of hybrid larch and Sitka spruce. This controlled experiment is the first stage of a larger study involving field experiments examining the growth of the species under various levels of shade.
Materials and methods
The shadehouses
Shadehouses (2 m(h) × 2.5 m(w) × 5 m(l)) were constructed on a 0.2-ha section of an open field at the Thornfield Research Complex, located on the Belfield Campus of University College Dublin. Green polypropylene shade fabrics of different mesh gauges were erected on frames to simulate three shade treatments: 25%, 50% and 75%. Frames without shade fabric served as controls (i.e. 0% shade). The shadehouses were placed on Mypex ground cover in the open, 2 m apart to prevent treatment overlap. The experimental area was divided into three blocks to account for any shade effect from adjacent buildings (all of which were one storey tall and at least 20 m away from the shadehouses).
The plants
In April 2003, containerised Sitka spruce (Irish provenance, origin Queen Charlotte Islands, Canada), hybrid larch (British provenance, origin unknown) and western red cedar seedlings (provenance and origin Hoquaim Washington) were purchased from a container nursery in Aughrim, Co. Wicklow. The seedlings had been grown in this operational nursery according to the following standard nursery practice. The Sitka spruce seedlings were derived from seed that had been sown in early spring 2002 in containers in a polyhouse in the nursery where they remained until June of that year. The seedlings were then moved outdoors until the following spring. The seed of both hybrid larch and western red cedar were sown in the same nursery in late spring 2001 in containers in a polyhouse where they also remained until June. The seedlings were moved outdoors in the spring of 2003. The mean seedling heights were 42 cm (s.e. 0.53 cm) for hybrid larch, 30 cm (s.e. 0.30 cm) for Sitka spruce and 39 cm (s.e. 0.41 cm) for western red cedar. The mean root collar diameters were 5.8 mm (s.e. 0.092 mm) for hybrid larch, 4.5 mm (s.e. 0.047 mm) for Sitka spruce and 5.2 mm (s.e. 0.077 mm) for western red cedar. The heights and diameters of the seedlings are typical for seedlings used in forest establishment in Ireland. At the experimental site the seedlings were put into 3-l pots with potting compost. They were then placed in a glasshouse and watered for one month. During this period, weeding took place once.
In May 2003, 1,008 (336 × 3 species) seedlings were taken from the glasshouse and placed in the shadehouses. A total of 28 potted seedlings of each species were placed in each level of shade. The inner 10 seedlings served as sample seedlings while the outer 18 acted as buffer seedlings. The total number of trees in each block was 336. The hybrid larch, which tends to have soft shoots, was staked with 0.5-m split canes to reduce overlapping and shoot breakage. This support was removed when the seedlings were repotted for the second growing season. The pot sizes used allowed the roots to grow freely at all stages during the experiment. All seedlings were watered to field capacity on a twice-weekly basis during the summer months and on a weekly basis otherwise. One litre of Kemira Feed (167 mg/l N; 46 mg/l P; 291 mg/l K; 140 mg/l Ca; 35 mg/l Mg; 64 mg/l S; 0.5 mg/l B; 0.6 mg/l Mn; 0.2 mg/l Cu; 0.4 mg/l Zn; 0.05 mg/l Mo) was applied to each pot every 8 weeks to ensure that lack of nutrients did not become a confounding factor over the duration of the study. Weeding was carried out when required.
Morphological measurements
At the beginning and at the end of the first growing season, the heights (cm) and the root collar diameters (mm) of all sample seedlings were measured. The height and root collar diameter increments at the end of one growing season were the differences in the two sets of values. At the end of the first growing season, three seedlings of each species (two sample seedlings and one buffer seedling) were removed from each shade treatment per block for a total of 108 seedlings. The remaining seedlings were re-potted into 10-l pots. The heights of the shadehouses were increased to 2 m at this time to allow for further unrestricted height growth.
The selected seedlings were carefully removed from the pots and most of the soil was removed from the roots by gentle shaking. Any remaining soil was removed by washing the roots with tapwater on site. The shoots and roots of each of the seedlings were separated by excising the stem at root collar after which they were dried in an oven for 24 h at 105°C. The shoot and root dry weights of each plant were determined after drying.
At the end of the second growing season the heights and root collar diameters of the remaining sample seedlings were measured and the increments calculated for two growing seasons. At this stage the remaining 288 sample seedlings were harvested and biomass data collected following the procedure outlined earlier.
Assessments were made of two aspects of leaf morphology, namely specific shoot area (SSA) and the normalised specific projected shoot area at the end of the second growing season. Measurements were only taken in three shade treatments (i.e. 0%, 50% and 75% shade) as they formed part of another section of the study examining the physiological responses of the seedlings to shade (e.g. carbon isotope discrimination) (Kennedy et al. submitted). The measurement of these physiological characteristics is time consuming and costly thus it was limited to only three shade treatments. To estimate SSA, sections of branch and foliage, 8 cm long including the tip, were removed from the second whorl down from the top of the plant in three randomly chosen seedlings in the three shade treatments. The sections were placed on a scanner and the one-sided projected shoot and leaf area was measured using Scion Imaging Software (Beta 4.0.1, Scion Corporation, Maryland, USA). They were then placed in an oven at 80°C and dried for 48 h. The ratio of projected leaf and stem area to leaf and stem dry mass, i.e. SSA (cm2/g) was then calculated.
To make inter-specific comparisons of the amount of light interception per unit area for an individual shoot, additional 6 cm long sections of shoot tip were taken from the same whorl as had been used for measuring SSA. These were then trimmed so as not to extend beyond 6 cm in width and images were obtained using the flat bed scanner and Scion Imaging Software to calculate the normalised specific projected shoot area, i.e. the ratio of projected leaf and stem area (cm2) to a standard area of 36 cm2.
Experimental design and data analysis
The experimental design was a split plot. The four shade treatments (i.e. the main treatments) were randomly assigned within three blocks and within each shade treatment the groups of 28 seedlings of each tree species (i.e. the subtreatments) were assigned at random to one of three positions.
Tests for normality and homogeneity of variance were undertaken on the data to ensure that the assumptions of ANOVA were satisfied. The data were then analysed using the ANOVA procedure on the GENSTAT statistical package (Payne 2002) with light levels, species and their interaction as model parameters. When the species × shade interaction was not significant (α = 0.05) the main effects of species and shade were tested further using Fisher’s protected least significant difference procedure. When the interaction was significant, shade treatments were compared separately for each species.
A preliminary analysis of the data showed that the mean initial heights and root collar diameters of the species differed significantly. Thus the height increment and root collar diameter increment data were initially converted to relative height growth and relative root collar diameter growth as per the formulae below and an ANOVA undertaken:
where H 1 = initial height (cm), H 2 = height at end of first growing season (cm), D 1 = initial diameter (mm) and D 2 = diameter at end of first growing season (mm).
The results of the ANOVA of both the relative height growth data and relative root collar diameter growth data were similar to those of the actual increment data so the results that are presented are those for actual increment. It was not possible to undertake a similar test for the biomass data because there were too few plants to carry out destructive tests at the beginning of the period of study. However, biomass relative growth rate (RGR) between the first and second growing seasons was calculated (using a similar formula to that used above for height and root collar diameter growth); these data would provide useful information as the seedlings should have acclimated to the shade treatments by the beginning of the second year. Thus the biomass data presented includes the actual biomass values (g dry weight) at the end of both the first and second growing seasons as well as the biomass RGR during this period.
Results
Survival, height increment and root collar diameter increment
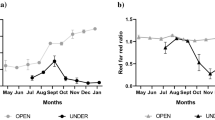
Over the entire study period, i.e. 2 years, only two fatalities were recorded. These occurred in the hybrid larch in 0% shade and were due to disturbance by foxes. Height increment and root collar diameter increment were significantly influenced by shade in both growing seasons (Table 1). At the end of the first growing season, the mean height increment of seedlings of the three species was greater in 25%, 50% and 75% shade than in 0% shade (Table 3). In contrast, the root collar diameter increment was lower in 75% shade than in all other shade treatments (Table 3). As expected, height increment and root collar diameter increment differed significantly between species regardless of shade treatment at the end of the first growing season (Table 1). The greatest height and root collar diameter increment was recorded in hybrid larch while the lowest was found in Sitka spruce (Table 3). While there was no significant interaction between shade and species with respect to height increment and root collar diameter increment this was not true for the H:D ratio (Table 1, Fig. 1). In both hybrid larch and Sitka spruce the H:D ratio increased with increasing shade with the ratio in the most shaded plots being 30% higher for both species than the ratio in the unshaded plots. In contrast, the relative increases in the H:D ratio in western red cedar with increasing shade were much lower.

H:D ratios (at the end of one growing season) and S:R ratios (at the end of two growing seasons) of hybrid larch, Sitka spruce and western red cedar in four levels of shade (means followed by different letters are significantly different at the α ≤ 0.05 level for within-species comparisons). Vertical lines on bars are standard errors
By the end of the second growing season, height increment in the most shaded plots was significantly lower than in all other shade treatments; however, height increment remained significantly higher in 25% and 50% shade than in the open (Table 4). In contrast, the root collar diameter increment was significantly lower in 50% and 75% shade than in the open (Table 4). At this stage the mean height and root collar diameter increments of western red cedar and hybrid larch were similar and significantly greater than those of Sitka spruce. There was no significant interaction between species and shade for the H:D ratio at the end of the second growing season (Table 1) with this ratio greater for all species in 25%, 50% and 75% shade than in 0% shade (Table 4). The smallest H:D ratio was found in Sitka spruce while the ratio was greatest in hybrid larch (Table 4).
Total biomass, shoot and root biomass and S:R ratios
Shade did not have a significant effect on total biomass, shoot biomass, root biomass or the S:R ratio in the three species at the end of the first growing season (Table 2). As expected, the biomass characteristics did differ between species with all mean values for the biomass characteristics in western red cedar significantly greater than those in the two other species (Table 3). The trend with regards to shade differed at the end of the second growing season (Table 2). At this stage total biomass declined significantly with increasing shade (Table 4), but there was no statistically significant interaction between species and shade. A similar trend was noted in shoot and root biomass as well as biomass RGR. Western red cedar continued to have greater total, root and shoot biomass than the two other species while there was no significant difference between species with respect to biomass RGR. However, a species-specific response to increasing shade was noted in the S:R ratios (Table 2, Fig. 1). The S:R ratio of western red cedar consistently increased with shade with the ratio 74% higher in the most shaded plots than in the unshaded plots. In hybrid larch, the S:R ratio in 75% shade was significantly greater than in 0% shade but only by 42%. In contrast, there was no significant difference in the S:R ratios of Sitka spruce in the four shade treatments.
SSA and normalised specific projected shoot area
The trend in SSA among species over the three shade treatments differed significantly as indicated by the significant interaction between species and shade (Table 5, Fig. 2). While all species had significantly greater SSAs in 75% shade than in 0% shade, this trend was particularly pronounced in hybrid larch. There was a significant interaction between species and shade for the normalised specific projected shoot area (Table 5, Figs. 2 and 3). In both hybrid larch and western red cedar, the normalised specific projected shoot area increased significantly with increasing shade albeit to a much greater extent in the latter species. The opposite trend was observed for Sitka spruce.

SSA and the normalised specific projected shoot area of Sitka spruce, hybrid larch and western red cedar grown in three shade treatments for two years (means followed by different letters are significantly different at the α ≤ 0.05 level for within-species comparisons). Vertical lines on bars are standard errors

Images of terminal shoot sections from branches of the three different species grown in 0%, 50 % and 75 % shade for two growing seasons
Discussion
The species chosen for this study were selected to represent a range of shade tolerances with hybrid larch classed as a light demanding species, Sitka spruce variably described as light-demanding, shade-tolerant or intermediate in its light requirements while western red cedar is considered to be shade-tolerant. Some morphological and structural characteristics of these three species were compared in a range of shade levels.
Survival, height increment and root collar diameter increment
In this study, survival rates of the three species were not influenced by shade with the only two fatalities attributed to disturbance by foxes. Greater mortality in the less shade-tolerant species was expected in the more shaded plots as lower survival rates than those of shade-tolerant species have been noted for these species previously in both natural (Daniel et al. 1979; Pacala et al. 1994; Chen 1997; Kaelke et al. 2001) and controlled environmental conditions (Fairbairn and Neustein 1970; Walters and Reich 1996). Height increment has also been shown to decline in shade, with the rate of decline usually less in shade-intolerant species than in shade-tolerant species. Beaudet and Messier (1998) found that the decline in height increment in yellow birch (Betula alleghaniensis Britton) with increasing shade was less than that recorded for beech (Fagus grandifolia Ehrh). Similarly the root collar diameter growth of tree species is expected to decline as shade increases. Pacala et al. (1994) noted a higher diameter growth in shade-tolerant temperate conifer and broadleaved species than in the shade-intolerant species in low light conditions. However, the results from our study do not agree with these previous findings. The first growing season’s data show clearly that height increment in the three species was higher in all shade levels than in the open contrary to expectation. This trend was also evident at the end of the second growing season although height increment in the most shaded plots (75% shade) was lower than in the open. This would suggest that the expected decline in height growth in the shade may be time dependent, especially in seedlings that have already been in unshaded conditions in a nursery for a number of growing seasons before being exposed to shade. Our analysis also showed that there was no significant interaction between species and shade for both height increment and root collar diameter increment. While all three species showed the classic decline in root collar diameter increment with increasing shade, there was no evidence of a significant difference in the relative decline in root collar diameter between the three species.
The ratio of tree height to diameter is a characteristic that has been shown in some species to be influenced by shade. Chen (1997) found that the H:D ratio in both Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) and Englemann spruce (Picea engelmannii Parry ex Engelm) increased significantly with increasing shade. However, the same author found that shade did not significantly influence the H:D ratio in ponderosa pine (Pinus ponderosa Dougl.). In our study there was evidence of a species-specific response to shade in relation to the H:D ratio at the end of the first growing season only. This was due largely to the lower H:D ratio in western red cedar in 50% compared to 25% shade while in the two other species the H:D ratio increased with increasing shade. At the end of the second growing season the response of the H:D ratio to shade was not species-dependent.
Biomass
Most researchers agree that total biomass production in shaded tree seedlings is lower than that in tree seedlings growing in higher light levels (Wang et al. 1994; Mitchell and Arnott 1995; Chen 1997; Khan et al. 2000). However, there is conflicting evidence as to how the shade-tolerance of a tree species influences this trend. Walters et al. (1993) in their study of northern hardwood species and Kitajima (1994) in his study of 13 tropical species found that shade-tolerant species show a greater reduction in total biomass in the shade than shade-intolerant species. However, Khan et al. (2000) found no significant difference in the rate of decline in total biomass with shade in a number of temperate coniferous species including western red cedar. In our study the total, shoot and root biomass as well as the biomass RGR of western red cedar, hybrid larch and Sitka spruce declined significantly with increasing shade at the end of the second growing season and there was no evidence of a species-dependent response. Only in the case of the S:R ratio data was a significant interaction between species and shade found. However, the reaction of the species to increasing shade was not as expected with the shade-tolerant western red cedar increasing its S:R ratio as shade increased to a greater extent that the shade-intolerant hybrid larch. Sitka spruce demonstrated the characteristics of a shade-tolerant species by decreasing its total biomass in the shade yet maintaining a similar S:R ratio at all shade levels.
Foliar characteristics
Specific leaf area describes the efficiency with which a leaf captures light relative to the biomass invested in leaf tissue (Marshall and Monserud 2003). In this study SSA was assessed in order to include all the foliage on the shoot as it was not possible to separate the foliage and the shoot in the case of western red cedar. SSA is effectively specific leaf area albeit at a different organisational scale. In our study, the shade-intolerant hybrid larch showed the greatest response to shade. The SSA of this species was 100% greater in the most shaded plots compared to the open, whereas in Sitka spruce and western red cedar it was only 58% and 60%, respectively. Groninger et al. (1996) also found that shade-intolerant species exhibited a greater increase in SLA in the shade compared with shade-tolerant species although Chen et al. (1996) found the opposite. It may be, as Groninger et al. (1996) suggest, that SLA may be a shade response “symptom” rather than an acclimation to a low light intensity environment. Furthermore, while a high SLA may allow seedlings to harvest light more efficiently in the shade, it makes the seedlings more vulnerable to water stress (Jones and McLeod 1990) and less resistant to herbivory (Kitajima 1994). Therefore, it is likely that a low SLA in shade-tolerant species, combined with longer-lived leaves, is a successful strategy for survival in shaded environments (De Lucia et al. 1998; Sack and Grubb 2002).
The analysis of the normalised specific projected shoot area showed a substantial inter-species response to changing shade levels. In western red cedar, the normalised specific projected shoot area increased significantly with increasing shade. Furthermore, over the study period, the authors observed that the angular orientation of the leaves of this species became more planar in the shade whereas in the unshaded plots it was erect. Thus the surface area exposed to light is reduced in the open in western red cedar suggesting a light avoidance strategy while in the shade the area available for photosynthesis is increased. In contrast, Sitka spruce exhibited a shade avoidance strategy by reducing its normalised specific projected shoot area in the shade. Hybrid larch exhibited only small increases with increasing shade.
Conclusion
For many of the morphological characteristics examined in this study the average response of the three species to the changing shade levels was as expected. However, the rate at which many of the characteristics changed with increasing shade did not differ between the species. This latter finding was not expected given the reported difference in the shade tolerance ranking of the three species. This may be partly explained by the fact that the study was undertaken in an environment where water and nutrient resources were not limiting. In contrast, the shade tolerance rankings of the species used in this study are derived from field observations using concepts that do not separate light from the confounding effects of microclimate, edaphic factors and competition (Kimmins 1987). Thus the reported variation in response in tree species to shade according to their shade tolerance ranking may only emerge where other resources such as water also change with shade (Black et al. 2005). The species-specific responses that emerged in the S:R ratio and leaf morphology data in general support the categorisation of hybrid larch as light-demanding. However, in the case of western red cedar and Sitka spruce there appears to be contradictory evidence from these characteristics as to their shade tolerance ranking. Analysis of some physiological adaptations of the species to increasing shade may help to resolve these contradictions (Kennedy et al. submitted).
References
Beaudet M, Messier C (1998) Growth and morphological responses of yellow birch, sugar maple, and beech seedlings growing under a natural light gradient. Can J For Res 28:1007–1015
Black K, Davis P, McGrath J, Doherty P, Osborne B (2005) Interactive effects of irradiance and water availability on the photosynthetic performance of Picea sitchensis seedlings: implications for seedling establishment under different management practices. Ann For Sci 62:413–422
Chen HYH (1997) Interspecific responses of planted seedlings to light availability in interior British Columbia: survival, growth, allometric patterns, and specific leaf area. Can J For Res 27:1383–1393
Chen HYH, Klinka K (1996) Light availability and photosynthesis of Pseudotsuga menziesii seedlings grown in the open and in the forest understory. Tree Physiol 17:23–29
Chen HYH, Klinka K, Kayahara GJ (1996) Effects of light on growth, crown architecture, and specific leaf area for naturally established Pinus contorta var. latifolia and Pseudotsuga menziesii var. glauca saplings. Can J For Res 26:1149–1157
Claveau Y, Messiers C, Comeau PG, Coates KD (2002) Growth and crown morphological responses of boreal conifer seedlings and saplings with contrasting shade tolerance to a gradient of light and height. Can J For Res 32:458–468
Daniel TW, Helms JA, Baker FS (1979) Principles of silviculture, 2nd edn. McGraw-Hill, New York
De Lucia EH, Sipe TW, Herrick J, Maherali H (1998) Sapling biomass allocation and growth in the understory of a deciduous hardwood forest. Am J Bot 85:955–963
Fairbairn WA, Neustein SA (1970) Study of responses of certain coniferous species to light intensity. Forestry 43:57–71
Fownes JH, Harrington RA (2004) Seedling responses to gaps: separating effects of light and nitrogen. For Ecol Manage 203:297–310
Givnish TJ (1988) Adaptation to sun and shade: a whole-plant perspective. Aust J Plant Physiol 15:63–92
Groninger JW, Seiler JR, Peterson JA, Kreh RE (1996) Growth and photosynthetic responses of four Virginia Piedmont tree species to shade. Tree Physiol 16:773–778
Horgan T, Keane M, McCarthy R, Lally M, Thompson D (2003) A guide to forest tree species selection and silviculture in Ireland. COFORD, National Council for Forest Research and Development, Agriculture Building, Belfield, Dublin 4, Ireland
Hunt R (1982) Plant growth curves: the functional approach to plant growth analysis. University Park Press, Baltimore, MD
Jones RH, McLeod KW (1990) Growth and photosynthetic responses to a range of light environments in Chinese tallow tree and Carolina ash seedlings. For Sci 36:851–862
Kaelke CM, Kruger EL, Reich PB (2001) Trade-offs in seedling survival, growth, and physiology among hardwood species of contrasting successional status along a light-availability gradient. Can J For Res 31:1602–1616
Khan SR, Rose R, Haase DL, Sabin TE (2000) Effects of shade on morphology, chlorophyll concentration and chlorophyll fluorescence of four Pacific Northwest conifer species. New Forests 19:171–186
Kimmins JP (1987) Forest ecology. Macmillan Publishing Co., New York
Kitajima K (1994) Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees. Oecologia 98:419–428
Marshall JD, Monserud RA (2003) Foliage height influences specific leaf area of three conifer species. Can J For Res 33:164–170
Mason B, Kerr G, Simpson J (1999) What is continuous cover forestry? Forestry Commission Information Note 29. Forestry Commission, Edinburgh
Messier C, Doucet R, Ruel JC, Claveau Y, Kelly C, Lechowicz MJ (1999) Functional ecology of advance regeneration in relation to light in boreal forests. Can J For Res 29:812–823
Mitchell AK, Arnott JT (1995) Effects of shade on the morphology and physiology of amabilis fir and western hemlock seedlings. New Forests 10:79–98
Pacala SW, Canham CD, Silander JA, Kobe RK (1994) Sapling growth as a function of resources in a north temperate forest. Can J For Res 24:2172–2183
Payne RW (ed) (2002) The guide to Genstat: syntax and data management Release 6.1. Lawes Agricultural Trust (Rothamsted Experimental Station)
Robakowski P, Wyka T, Samardakiewicz S, Kierzkowski D (2004) Growth, photosynthesis, and needle structure of silver fir (Abies alba Mill) seedlings under different canopies. For Ecol Manage 201:211–227
Sack L, Grubb PJ (2002) The combined impact of deep shade and drought on the growth and biomass allocation of shade-tolerant woody seedlings. Oecologia 131:174–185
Savill PS (1992) The silviculture of trees used in British forestry. CAB International, Oxford
Van Hees AFM, Clerkx APPM (2003) Shading and root-shoot relations in saplings of silver birch, pedunculate oak and beech. For Ecol Manage 176:439–448
Walters MB, Reich PB (1996) Are shade tolerance, survival, and growth linked? Low light and nitrogen effects on hardwood seedlings. Ecology 77:841–853
Walters MB, Kruger EL, Reich PB (1993) Growth, biomass distribution and CO2 exchange of northern hardwood seedlings in high and low light:relationships with successional status and shade tolerance. Oecologia 94:7–16
Wang GG, Qian H, Klinka K (1994) Growth of Thuja plicata seedlings along a light gradient. Can J Bot 72:1749–1757
Wenger KF (1984) Forestry handbook, 2nd edn. Society of American Foresters. John Wiley & Sons, Inc., New York
Acknowledgements
We would like to thank Tom Moore, Ray O’Haire and Denis Coghlan (School of Biology and Environmental Science, University College Dublin) and Donal O’Hare (Forestry Consultant) for their technical assistance in setting up the experiment and sampling of seedlings. The National Council for Forest Research and Development (COFORD) provided funding for this research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kennedy, S., Black, K., O’Reilly, C. et al. The impact of shade on morphology, growth and biomass allocation in Picea sitchensis, Larix × eurolepis and Thuja plicata . New Forests 33, 139–153 (2007). https://doi.org/10.1007/s11056-006-9019-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-006-9019-2