
Abstract
Knowledge of natural ecology is essential for a better understanding of pathogenicity and opportunism in black yeast-like fungi. Although etiological agents of diseases caused by these fungi are supposed to originate from the environment, their isolation from nature is difficult. This is probably due to their oligotrophic nature, low competitive ability, and, overall, insufficient data on their natural habitat. We obtained environmental samples from mangrove areas where mortalities by lethargic crab disease (LCD) are reported and areas without disease recorded. Isolation of chaetothyrialean black yeasts and relatives was performed using a highly selective protocol. Species-specific primers were used to determine if these isolates represented Exophiala cancerae or Fonsecaea brasiliensis, two proven agents of LCD, in order to test hypotheses about the origin of the disease. Isolates, identified by morphology as Fonsecaea- or Exophiala-like, were tested specific diagnostic markers for the fungi associated with LCD. Although several black fungi were isolated, the main causative agent of the LCD, E. cancerae, was not found. Molecular markers for F. brasiliensis revealed 10 positive bands for isolates from biofilms on mangrove leaves, branches, and aerial roots, of which four were confirmed by ITS sequencing. The absence of E. cancerae in environmental samples suggests that the species is dependent on the crab, as a genuine pathogen, different from F. brasiliensis, which is probably not dependent on the host species, U. cordatus. However, we did not attempt isolation from the marine water, which may represent the pathway of dispersion of the black yeast species between neighbor mangroves.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The mangrove land crab, Ucides cordatus (Brachyura/Ocypodidae), plays a crucial role in a variety of ecosystem processes in its environment, being involved in the decomposition of organic materials [1, 2]. In Brazil, the animal is an important component in the economy of several underprivileged communities that depend on it for their subsistence.
Since the year 1997, an extensive epizootic causes high mortalities of U. cordatus along the Brazilian coast [1], causing a reduction of up to 85 % of the collection rates in some regions in the northeastern region. Affected crabs share several symptoms, such as lethargy, poor motor control, and inability to return to the upright position when turned upside down. The disease was termed Lethargic Crab Disease (LCD) by Boeger et al. [1] because of clinical signs of ataxia observed in moribund organisms [1, 2]. Evidence from a variety of sources (light and electron microscopy, behavioral and experimental tests [1], molecular phylogenetics [1], and experimental infection [3]) indicates that LCD signs are associated with the presence of a black yeast-like fungus, Exophiala cancerae recently described by de Hoog et al. [4]. However, occasional co-infection may take place by another black yeast-like fungus, viz. a species of Fonsecaea recently described by Vicente et al. [5] as F. brasiliensis. Since the first record of mortalities likely associated with LCD in Goiana, Pernambuco State, Brazil, the disease has spread from Northern to Southern estuaries along to South Atlantic Coast in a wave-like pattern [6]. Currently, LCD is apparently absent from the majority of estuaries where it was reported in the past, having remained solely in the mangroves of Espirito Santo State.
Knowledge of natural ecology and evolution is essential for a better understanding of pathogenicity and opportunism of black yeast-like fungi. Although etiological agents of most diseases caused by black yeast-like fungi are supposed to originate from the environment, their isolation from nature is difficult. This is probably due to their oligotrophic nature, low competitive ability, and in general insufficient data on their natural habitat. Several selective techniques have been developed to enable recovery of these fungi [7–12]. Several studies indicate that opportunism of these fungi might be explained from the perspective of their unusual niches [13–15]. Recurrent and consistent infections caused by many representatives of the order Chaetothyriales indicate a possible specialization of the fungi to the animal hosts.
In this study, we obtained environmental samples from mangrove areas in which mortalities by LCD are reported and areas where the disease is not recorded. Isolation of chaetothyrialean black yeasts and relatives was performed using a highly selective protocol. Species-specific diagnostic primers were used to determine if these isolates represented E. cancerae or Fonsecaea brasiliensis, in order to monitor the spread of agents of LCD along the Brazilian coast.
Materials and Methods
Hypothesis Testing
The sampling design of this study is focused on the hypothesis that the fungi associated with LCD are opportunistic, that is, emerging from the environment. The null hypothesis is accepted if the fungus species is found in the environment, at least in areas where LCD outbreaks occur. The alternative hypothesis suggests that the fungus is a real pathogen. This hypothesis will be supported if the fungus consistently associated with LCD cannot be isolated from environmental samples in mangroves where LCD is prevalent.
Fungal Strains
Strains analyzed (52) are listed in Table 1. Stock cultures were maintained on 2 % malt extract agar (MEA, Difco) and oatmeal agar (OA) slants at 24 °C. For morphological studies, MEA slide cultures were prepared and mounted in aniline blue.
Study Areas and Sampling Strategy
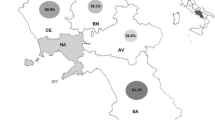
Locations chosen for this study include three mangrove areas where mortalities of mangrove land crabs putatively caused by LCD have been reported in the state of Bahia, viz. Acupi, Saubara, and two areas in Canasvieiras indicated as −1 and −2 (Fig. 1; Table 1). In each location, fragments of plant material and decaying wood showing macroscopic presence of black biofilms, as well as soil samples, were obtained randomly for a total of approximately 1 kg of samples per site. Approximately 20 g from each sample was processed for fungal isolation, with 50 replicates/sample.

Collection sites along the coast of Bahia State. Mangroves where samples were taken are indicated with closed circles
Fungal Isolation
Each sample was incubated at room temperature for 30 min in 100 mL of a sterilized saline solution containing 200 U of penicillin, 200 μg/L of streptomycin, 200 μg/L of chloramphenicol, and 500 μg/L of cycloheximide. After initial incubation, 20 mL of sterilized mineral oil was added to the solution, and the container was vigorously shaken for 5 min and left to settle for 20 min. The oil–water interphase was then collected and inoculated on Mycosel agar (Difco) for 4 weeks at 36 and 30 °C [8, 16, 17]. Dark colonies were selected then isolated and stored on Mycosel agar.
Identification
Identification was carried out using macro- and microscopic features of the colonies after culture on Sabouraud Dextrose Agar (SDA, Difco) at room temperature. The isolates were analyzed by molecular methods using ITS markers [17]. Cardinal growth temperatures were determined on 2 % malt extract agar (MEA, Difco). Plates were incubated in the dark for 3 weeks at temperatures of 21–36 °C at intervals of 3 °C; growth was also recorded at 37 and at 40 °C.
DNA Extraction
Approximately 1 cm2 of 14–21-day-old cultures was transferred to a 2 ml Eppendorf tube containing 400 μL TEx buffer (pH 9.0) and glass beads (Sigma G9143). The fungal material was homogenized with MoBio vortex for 1 min. Subsequently, 120 μL SDS 10 % and 10 μL proteinase K were added and incubated for 30 min at 55 °C, and the mixture was vortexed for 3 min. After addition of 120 μL of 5 M NaCl and 1/10 vol CTAB 10 % (cetyltrimethylammonium bromide) buffer, the material was incubated for 60 min at 55 °C. Then, the mixture was vortexed for 3 min. Subsequently, 700 μL 24:1 chloroform/isoamylalcohol was added, mixed carefully by hand and centrifuged for 5 min at 4 °C at 20,400g force value. The supernatant was transferred to a new Eppendorf tube with 225 μL 5 M NH4-acetate, mixed carefully by inverting, incubated for 30 min on ice water, and centrifuged again for 5 min at 4 °C at 20,400g force value. The supernatant was then transferred to another Eppendorf tube with 0.55 vol. isopropanol and centrifuged for 5 min at 20,400 g force value. Finally, the pellet was washed with 1000 μL ice cold 70 % ethanol. After drying at room temperature, it was re-suspended in 100 μL TE buffer (Tris 0.12 % w/v, Na-EDTA 0.04 % w/v).
PCR Diagnosis of Fungi Associated to LCD
Species-specific PCR-primers for E. cancerae [18] and F. brasiliensis [5], designed on the basis of their ITS rDNA sequence, were applied to the DNA extracts to determine whether the isolate from the environment represented either species. PCR products were electrophoresed on 1.5 % agarose gels, stained with ethidium bromide and were photographed under UV light. The samples were determined to be positive for the yeast strain if they had an amplicon of *450 or *396 bp, which corresponds to the expected product size of E. cancerae (strain CBS 120420) and F. brasiliensis (strain CBS 119710).
DNA Amplification and Sequencing
Sequencing was done for strains that were positive in F. brasiliensis or E. cancerae specific PCRs. ITS amplicons were generated with primers V9G and LS266 [19] and were sequenced with primers ITS1 and ITS4. PCR was performed in a 25 μL volume of a reaction mixture containing 7 μL Go Taq master mix (Promega, Breda, The Netherlands) containing dNTPs, MgCl2, reaction buffer, 1 μL of each primer (10 pmol), and 1 μL rDNA. Amplification was performed in an ABI PRISM 2720 (Applied Biosystems, Foster City, USA) thermocycler as follows: 94 °C for 2 min, followed by 30 cycles consisting of 94 °C for 30 s, 94 °C for 30 s and 65 °C for 1 min, and a delay at 72 °C for 5 min. Amplicons were cleaned with GFX PCR DNA and Gel Band Purification kit (GE Healthcare, Buckinghamshire, UK). Concentrations of amplicons were estimated on gel, photographed and analyzed by the Gel Doc XR system (Biorad, Veenendaal, The Netherlands), with SmartLadder (Eurogentec, Seraing, Belgium) as size and concentration marker. Amplicons were subjected to direct sequencing, with PCR was as follows: 96 °C for 1 min, followed by 35 cycles consisting of 96 °C for 10 s, 50 °C for 5 s and 60 °C for 4 min. Reactions were purified with Sephadex G-50 fine (GE Healthcare Bio-Sciences AB, Uppsala, Sweden), and sequencing was done using ABI prism BigDye™ terminator cycle sequencing kit (Applied Biosystems) and analyzed on an ABI Prism 3730XL Sequencer. Sequences were edited using SeqMan in the Lasergene software (DNASTAR, Wisconsin, USA). Iterative alignment was performed by hand with BioNumerics version 4.61 (Applied Maths, St.-Martens-Latem, Belgium).
Results
A total of 64 environmental samples were processed, 36 of which were positive for black fungi (56.3 %). Among these, samples from branches of mangrove trees presented higher frequencies of isolation (87.5 %; Tables 2, 3). Fifty-two isolates, identified by macro- and micro-morphology as Fonsecaea- or Exophiala-like, were submitted to specific diagnostic markers for the fungi associated to LCD (Fig. 2). Despite the large numbers of black fungi isolated, no positive reaction was obtained for any of the environmental samples when analyzed with specific markers for the main causative agent of the LCD, E. cancerae (Fig. 2A). However, the same isolates submitted to molecular markers developed for F. brasiliensis, considered previously as a secondary agent of the disease [5], resulted in 10 positive bands (Fig. 2B) for the isolates from mangrove tree leaves (Canasvieiras1: CBS 127897; Acupi: R45A3I), mangrove branch (Canasvieiras 2: CBS 127891, CBS 127892, CBS 127893, CBS 127896; Acupi: CBS 127904, CBS 127905, R42C1), and aerial root (Canasvieiras 2: CBS 127898). Samples of areas with or without previous record of LCD mortality outbreaks presented isolates positively identified as F. brasiliensis. Among 10 isolates positive to molecular markers developed for F. brasiliensis, only four were confirmed by ITS sequence (Table 1) suggesting cross-reaction of the marker.

Specific PCR marker for the LCD etiological agents. a DNA extracts with amplification of black fungi using E. cancerae (main LCD agent) specific fragment. L. Molecular size marker (100 bp ladder); +positive control (E. cancerae CBS 120420); 1–41. DNA extracts of black fungi isolated from coast of Bahia state; −negative control. b 1–10. DNA extracts with amplification of black fungi using F. brasiliensis (secondary LCD agent) specific fragment. L. Molecular size marker (100 bp ladder); +positive control (F. brasiliensis CBS 119710); −negative control
Discussion
Many reports of epizootics from the older literature, for example, [20–22], reviewed by de Hoog et al. [4], clearly demonstrate that black yeast infection is a relatively common phenomenon in cold-blood vertebrates. Particularly cold-blooded waterborne vertebrates are susceptible to a diversity of Exophiala species [25]. Recent studies have shown that different pathogenic species are involved [23–25], which are morphologically very similar. Some of these are host-specific to, for example, particular fish taxa [24]. Chaetothyrialean black yeasts and relatives are interesting microorganisms from ecological as well as clinical points of view. Most pathogenic species belong to Exophiala, Cladophialophora, or Fonsecaea (Table 1), while also severe infections are observed in Veronaea [4, 5, 26]. Among the most serious pathogenic Exophiala species, eventually leading to disseminated, fatal infections in healthy hosts, are the neurotrope Exophiala dermatitidis [12], the osteotrope E. spinifera [27], and a species tending to cause disseminated infection, E. asiatica [29]. As virulence factors the ability to assimilate alkylbenzenes, present in sweat and nervous tissues of mammals and in the toxic skin of amphibians, has been suggested [4, 9]. The recurrent and consistent infections caused by many representatives of the order indicates a possible adaptation of the fungi to the vertebrate host. In the environment, they occupy specific microhabitats, probably due to their low competitive ability toward co-occurring microorganisms [16]. Their oligotrophism [10] enables them to thrive and maintain at low density in adverse substrates where common saprobes are absent [25, 28].
The mangrove crab epizootic [1, 2] underlines the ability of black yeasts to infect a larger spectrum of animals, also involving invertebrates. Recently [5] the etiology of the disseminated LCD in mangrove land crabs was clarified, describing the epidemiology of the disease and establishing a relative virulence in the etiological agents. As in most black fungi, an environmental source of infection is hypothesized, similar to, for example, agents of chromoblastomycosis, a disease found in humans and frogs [4]. Molecular siblings of common pathogenic species may be found in association with living plants, growing on debris of tannin-rich cactus spines; therefore, plants were supposed to be the vehicle of introduction into the human body [30].
In our study, F. brasiliensis, considered to be a secondary agent of lethargic crab disease, was frequently isolated from the crab habitat, whereas E. cancerae could never be isolated or detected in the mangrove environment. De Hoog et al. [4] established that the optimum growth temperature of E. cancerae is 24–27 °C, with a maximum at 33 °C, while the maximum of the F. brasiliensis lies at 37 °C, suggesting that temperature tolerance could be an important factor determining host preference.
According to de Hoog et al. [4], the infection pattern of members of Chaetothyriales suggests the existence of intrinsic factors enhancing vertebrate invasion, but infection is probably not a prime factor in the natural habitat of these fungi. In waterborne species, a slight correlation was observed between the maximum growth temperature of the fungus and the natural habitat of the host. Fungi with maximum growth temperatures below 33 °C tend to cause diseases in animals in cold waters. This is a remarkable feature, since elsewhere in the fungal kingdom, species lacking thermotolerance generally are saprobes without infectious abilities. Some species with maximum growth temperatures around 33 °C exceptionally up to 36 °C may cause zoonotics in cold-blooded animals living in shallow tidal zones in the subtropics. A striking example is the emerging Lethargic Crab Disease in crabs inhabiting mangrove areas along the east coast of Brazil [1, 2].
The absence of E. cancerae in the environmental samples evaluated supports the alternative hypothesis of this study, namely that the species would be dependent on the crab species, with direct transmission, thus likely representing a genuine pathogen [1, 4, 25]. In contrast, F. brasiliensis was isolated at least four times from mangrove trees with historical records of LCD outbreaks, providing strong support for the null hypothesis of this study, that is, LCD with an environmental origin. F. brasiliensis is likely an opportunistic species and is not dependent of the host animal, U. cordatus. Epidemiological records of LCD outbreaks indeed indicate that while E. cancerae is systematically detected in sick, moribund, and dead crabs, whereas F. brasiliensis has only been detected in few mortality events. These results support the earlier conclusion [3] that ascribed responsibility for the LCD to E. cancerae alone.
However, two points do not match with this conclusion. First, since E. cancerae has not been detected in the mangrove compartments evaluated in this study, it is not clear how the fungus dispersed from a central point of initial outbreak to mangroves south and north along the Brazilian coast. Our study focused on solid substrates because waterborne black yeasts quickly tend to form biofilms [31], but we have not sampled marine water, and sampling was done during a period without LCD outbreaks. It seems reasonable to postulate that the dispersion of propagules of E. cancerae might occur using the coastal water and currents and probably is more extensive during disease outbreaks. Second, the conclusion that E. cancerae would be an obligate pathogen is negated by the fact that de Hoog et al. [4] listed strains from different environments including water, fruit drinks, and human skin. Strain Cunningham 179/99 mentioned in the same study was involved in a systemic zoonosis in toads in Israel (A. Cunningham, pers. comm.).
During disease outbreaks, the organs and tissues of moribund and dead crabs are overwhelmed by fungal growth [2]. Fungal cells are likely to be released upon decomposition of crab carcasses and carried away into open and coastal water by tide currents, which are responsible for dispersion into neighboring mangrove areas. This hypothesis was suggested previously by Smith [32] based on the distribution of dead animals in the same mangrove area as sampled in the present study, Canasvieiras. Isolation of black yeasts from seawater during and between periods of active LCD outbreaks may provide the evidence for testing this putative mode of dispersion.
References
Boeger WA, Pie MR, Ostrensky A, Patella L. Lethargic crab disease: multidisciplinary evidence supports a mycotic etiology. Mem Inst Oswaldo Cruz. 2005;100:161–7.
Boeger WA, Pie MR, Vicente VA, Ostrensky A, Hungria D, Castilho GG. Histopathology of the mangrove land crab Ucides cordatus (Ocypodidae) affected by lethargic crab disease. Dis Aquat Organ. 2007;78:73–81.
Orélis-Ribeiro R, Boeger WA, Vicente VA, Chammas M, Ostrensky A. Fulfilling Koch’s postulates confirms the mycotic origin of Lethargic Crab Disease. Antonie Van Leeuwenhoek. 2011;99:601–8.
de Hoog GS, Vicente VA, Najafzadeh MJ, Harrak JM, Badali H, Seyedmousavi S. Waterborne Exophiala species causing disease in cold-blooded animals. Persoonia. 2011;27:46–72.
Vicente VA, Orélis-Ribeiro R, Najafzadeh MJ, Sun J, Guerra RS, Miesch S, Ostrenky A, Meis JF, Klaassen CH, de Hoog GS, Boeger WA. Black yeast-like fungi associated with Lethargic Crab Disease (LCD) in the mangrove-land crab, Ucides cordatus (Ocypodidae). Vet Microbiol. 2012;158:109–22.
Ferreira CP, Pie MR, Esteva L, Mancera PFA, Boeger WA, Ostrensky A. Modelling the lethargic crab disease. J Biol Dyn. 2009;3:620–34.
de Hoog GS, Göttlich E, Platas G, Genilloud O, Leotta G, van Brummelen J. Evolution taxonomy and ecology of the genus Thelebolus in Antarctica. Stud Mycol. 2005;51:33–76.
Dixon DM, Shadomy HJ, Shadomy S. Dematiaceous fungal pathogens isolated from nature. Mycopathologia. 1980;70:153–61.
Prenafeta-Boldú FX, Summerbell RC, de Hoog GS. Fungi growing on aromatic hydrocarbons: biotechnology’s unexpected encounter with biohazard. FEMS Microbiol Rev. 2006;30:109–30.
Satow MM, Attili-Angelis D, de Hoog GS, Angelis DF, Vicente VA. Selective factors involved in oil flotation isolation of black yeasts from the environment. Stud Mycol. 2008;61:157–63.
Zhao J, Zeng J, de Hoog GS, Attili-Angelis D, Prenafeta-Boldú FX. Isolation of black yeasts by enrichment on atmospheres of monoaromatic hydrocarbons. Microb Ecol. 2010;60:149–56.
Sudhadham M, Dorrestein GM, Prakitsin S, Sivichai S, Chaiyarat R, Prakitsin S, Menken SBJ, de Hoog GS. The neurotropic black yeast Exophiala dermatitidis has a possible origin in the tropical rain forest. Stud Mycol. 2008;61:145–55.
Sterflinger K, de Hoog GS, Haase G. Phylogeny and ecology of meristematic ascomycetes. Stud Mycol. 1999;43:5–22.
Wang CJK, Zabel RA. Identification manual for fungi from utility poles in the Eastern United States. Lawrence: Allen Press; 1990.
Lutzoni F, Pagel M, Reeb V. Major fungal lineages are derived from lichen symbiotic ancestors. Nature. 2001;411:937–40.
Iwatsu T, Miyaji M, Okamoto S. Isolation of Phialophora verrucosa and Fonsecaea pedrosoi from nature in Japan. Mycopathologia. 1981;75:149–58.
Vicente VA, Attili-Angelis D, Pie MR, Queiros-Telles F, Cruz LM, Najafzadeh MJ, de Hoog GS, Pizzirani-Kleiner A. Environmental isolation of black yeast-like fungi involved in human infection. Stud Mycol. 2008;61:137–44.
Pie MR, Boeger W, Patella L, Vicente VA, Orélis-Ribeiro R, Ostrensky A. Specific primers for the detection of the black-yeast fungus associated with lethargic crab disease (LCD). Dis Aquat Organ. 2011;94:73–5.
De Hoog GS, Gerrits van den Ende AHG. Molecular diagnostics of clinical strains of filamentous basidiomycetes. Mycoses. 1998;41:183–9.
Fijan N. Systemic mycosis in channel catfish. Wildl Dis. 1969;5:109–10.
Richards RH, Holliman A, Helgason S. Exophiala salmonis infection in Atlantic salmon Salmo salar L. J Fish Dis. 1978;1:357–68.
Reuter RE, Hutchinson W, Ham J, Davis S. Exophiala sp. infection in captured King George whiting (Sillaginodes punctata). Bull Eur Assoc Fish Pathol. 2003;23:128–34.
Manharth A, Lemberger K, Mylniczenko N, Pinkerton M, Pessier AP, Kammeyer P, de Hoog GS. Disseminated phaeohyphomycosis due to Exophiala species in a Galapagos tortoise, Geochelone nigra. J Herpetol Med Surg. 2005;15:20–6.
Nyaoke A, Weber ES, Innis C, Stremme D, Dowd C, Hinckley L, Gorton T, Wickes B, Sutton D, de Hoog S, Frasca S Jr. Disseminated phaeohyphomycosis in weedy, Phyllopteryx taeniolatus, and leafy, Phycodurus eques, seadragons caused by species of Exophiala, including a novel species. J Vet Diagn Invest. 2009;21:69–79.
De Hoog GS. Evolution of black yeasts: possible adaptation to the human host. Antonie Van Leeuwenhoek. 1993;63:105–9.
Bonifaz A, Badali H, de Hoog GS, Padilla-Desgarennes C, Vázquez-González D, Navarrete G, Meis JF. Severe disseminated phaeohyphomycosis in an immunocompetent patient caused by Veronaea botryosa. Mycopathologia. 2013 (in press).
Li DM, Li RY, de Hoog GS, Wang YX, Wang DL. Exophiala asiatica, a new species from a fatal case in China. Med Mycol. 2009;47:101–9.
De Hoog GS. Significance of fungal evolution for the understanding of their pathogenicity illustrated with agents of phaeohyphomycosis. Mycoses. 1997;40 Suppl 2:5–8. doi:10.1111/j.1439-0507.1997.tb00555.x.
Li DM, de Hoog GS, Lindhardt Saunte DM, Gerrits van den Ende AHG, Chen XR. Coniosporium epidermidis sp. nov., a new species from human skin. Stud Mycol. 2008;61:131–6.
de Hoog GS, Nishikaku AS, Fernández Zeppenfeldt G, Padín-González C, Burger E, Badali H, Gerrits van den Ende AHG. Molecular analysis and pathogenicity of the Cladophialophora carrionii complex, with the description of a novel species. Stud Mycol. 2007;58:219–34.
Heinrichs G, Hübner I, Schmidt GK, de Hoog GS, Haase G. Compositional analysis of dark fungal biofilms from water taps using Tag Encoded FLX Amplicon Pyrosequencing. Mycopathologia. 2013 (in press).
Schmidt AJ. Estudo da dinâmica populacional do caranguejo-uçá, Ucides cordatus (Linnaeus, 1763) (Crustacea-Decapoda-Brachyura), e dos efeitos de uma mortalidade em massa desta espécie em manguezais do Sul da Bahia. Dissertation (in Portuguese). Universidade de São Paulo, Brazil. 2006.
Acknowledgments
Bert Gerrits van de Ende are thanked for technical assistance. The work of Vania A. Vicente was supported by Brazilian Government fellowship from Coordenação de Aperfeiçoamento de Pessoal de Nivel Superior (CAPES) and the work of Mohammad Javad Najafzadeh by Faculty of Medicine, Mashhad University of Medical Sciences, Mashhad, Iran. The authors also wish to thank the Brazilian agencies CAPES/Ministério da Educação—Brazil and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for financial support.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Guerra, R.S., do Nascimento, M.M.F., Miesch, S. et al. Black Yeast Biota in the Mangrove, in Search of the Origin of the Lethargic Crab Disease (LCD). Mycopathologia 175, 421–430 (2013). https://doi.org/10.1007/s11046-013-9636-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11046-013-9636-1