
Abstract
Candida parapsilosis is yeast capable of forming biofilms on medical devices. Novel approaches for the prevention and eradication of the biofilms are desired. This study investigated the anticandidal activity of sixteen essential oils on planktonic and biofilm cultures of C. parapsilosis complex. We used molecular tools, enumeration of colony-forming units, the colourimetric MTT assay, scanning electron microscopy (SEM) and a chequerboard assay coupled with software analyses to evaluate the growth kinetics, architecture, inhibition and reduction in biofilms formed from environmental isolates of the Candida parapsilosis complex; further, we also evaluated whether essential oils would interact synergistically with amphotericin B to increase their anticandidal activities. Of the environmental C. parapsilosis isolates examined, C. parapsilosis and C. orthopsilosis were identified. Biofilm growth on polystyrene substrates peaked within 48 h, after which growth remained relatively stable up to 72 h, when it began to decline. Details of the architectural analysis assessed by SEM showed that C. parapsilosis complex formed less complex biofilms compared with C. albicans biofilms. The most active essential oil was cinnamon oil (CO), which showed anticandidal activity against C. orthopsilosis and C. parapsilosis in both suspension (minimum inhibitory concentration—MIC—250 and 500 μg/ml) and biofilm (minimum biofilm reduction concentration—MBRC—1,000 and 2,000 μg/ml) cultures. CO also inhibited biofilm formation (MBIC) at concentrations above 250 μg/ml for both species tested. However, synergism with amphotericin B was not observed. Thus, CO is a natural anticandidal agent that can be effectively utilised for the control of the yeasts tested.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fungi are increasingly important causes of acute and chronic deep-seated human infections, especially recurrent mucosal, cutaneous or nail infections that may be severe in debilitated or immunocompromised individuals. Candida spp. are commensal fungi that inhabit various niches of the human body; they are also considered the most common fungal pathogens isolated from clinical sites [1].
During the last decade, Candida parapsilosis was the second most common species of Candida isolated from blood cultures [1–3]. In South American hospitals, the incidence of C. parapsilosis is greater than C. albicans [3, 4]. C. parapsilosis has been renamed as following three distinct species: C. parapsilosis, C. orthopsilosis and C. metapsilosis [5].
Little information is available regarding the ability of the reclassified species of the C. parapsilosis to form biofilms. It is well established that microorganisms contained within biofilms exhibit increased resistance to anticandidal treatments compared to individual cells grown in suspension [6–9]; probably, this phenomenon is most likely caused due to a combination of poor anticandidal agent penetration, nutrient limitation, adaptive stress responses, induction of phenotypic variability and the formation of persistent cells [7, 9].
The small number of drugs available for fungi treatment, most of which are fungistatic, and the emerging resistance to antifungal agents encourage the search for alternatives [10, 11]. Secondary plant metabolites such as essential oils, which are mixtures of multiple heterogeneous compounds, can exhibit anticandidal activity because many essential oils are thought to disrupt cell membranes with their lipophilic components [12, 13].
The potential use of essential oils for the prevention and treatment of Candida biofilms has been researched in several studies [11, 14–21]. Therefore, we chose the following sixteen species of medicinal plants that produce essential oils to determine their anticandidal activity on C. parapsilosis complex biofilms: Boswellia thurifera, Cinnamomum zeylanicum, Citrus bergamia, Citrus aurantium amara, C. bigaradia, Citrus limonum, Citrus maxima, Commiphora myrrha, Copaífera officinalis, Eucalyptus globulus, Juniperus virginiana, Pogostemon patchouli, Salvia officinalis L., Santalum album, Thymus vulgaris and Zingiber officinale. We sought a possible source of new antibiotics for the treatment of biofilms from pathogenic fungi.
The purpose of this study was threefold. First, we aimed to compare the activities of the different essential oils against planktonic cells of Candida parapsilosis complex. Second, we tested the oils that demonstrated anticandidal activity against planktonic cells for their ability to prevent and treat Candida parapsilosis complex biofilms. Lastly, we determined the anticandidal activity of amphotericin B in combination with the oils actives against C. parapsilosis complex biofilms.
Materials and Methods
Yeast Strains
Seventeen environmental isolates of C. parapsilosis that were isolated from a hydraulic circuit of Hemodialysis Center in São Paulo State, Brazil, were tested in this study. They were phenotypically identified as C. parapsilosis by the use of API 20 C AUX (Biomerieux, Paris, France). Type strains ATCC 90018 (C. parapsilosis), ATCC 96141 (C. orthopsilosis), ATCC 96143 (C. metapsilosis), ATCC 6258 (C. krusei), ATCC 22019 (C. parapsilosis) and SC 5314 (C. albicans biofilm reference strain) were used as controls.
Essential Oils
The essential oils from the following plants were tested: Boswellia thurifera, Cinnamomum zeylanicum, Citrus Bergamia, Citrus aurantium amara, C. bigaradia, Citrus limonum, Citrus maxima, Commiphora myrrha, Copaífera officinalis, Eucalyptus globulus, Juniperus virginiana Pogostemon patchouli, Salvia officinalis L., Santalum album, Thymus vulgaris and Zingiber officinale. Plant essential oils were diluted in 5% (vol/vol) in dimethyl sulfoxide (DMSO—Sigma Chemical Co., St. Louis, MO) that had been filter sterilised through a 0.22-μm-pore-size filter (Millipore, Billerica, A, USA). All plant essential oils were obtained from a local Body and Mind Beautiful Com. de Cosméticos Ltda., Franca, SP, Brazil. The oils that exhibited anticandidal activity were related by the manufacturer as the major components: α-pinene and myrcene in juniper and cinnamaldehyde and caryophyllene in cinnamon oil.
Molecular Identification of C. parapsilosis Complex
Yeast genomic DNA was extracted according to the protocol described by Moller et al. [22]. The two-step DNA-based identification test described by Tavanti et al. [5] was used to screen C. parapsilosis isolates. Briefly, a 716-bp fragment of the secondary alcohol dehydrogenase (SADH) gene was amplified and digested with BanI (New England Biolabs, Hitchin, UK). The PCR amplification conditions were as follows: 2 min at 94°C; 35 cycles of 94°C for 30 s, 50°C for 30 s and 72°C for 30 s; and a final step of 10 min at 72°C. The PCR products were then digested with the BanI enzyme in a 40 μl volume containing 20 μl of the PCR product and 40 U of BanI. The digestion products were separated on 2% agarose gels. The isolates were discriminated as C. parapsilosis, C. orthopsilosis or C. metapsilosis according to their SADH gene restriction profiles.
The Effect of Essential Oils on C. orthopsilosis and C. parapsilosis Planktonic Growth
Antifungal testing to determine the MIC for planktonic cells was performed according to the Clinical and Laboratory Standard Institute (formerly the National committee for Clinical Laboratory Standards) M-27A2 broth dilution assay [23]. RPMI 1640 medium containing l-glutamine without sodium bicarbonate (pH 7.0) supplemented with 0.165 M morpholinepropanesulfonic acid (both from Sigma) was utilised. The spectrophotometric method was used for the preparation of inoculums with a concentration of 0.5 × 103 to 2.5 × 103 cells per ml for each of the isolates prepared in the test medium. Stock solutions of essential oils were prepared in 5% DMSO, and twofold serial dilutions were prepared in RPMI in 96-well microtitre plates (Corning Incorporated, Corning, NY, USA); final concentrations ranged from 15.6 to 8,000 μg/ml. Yeast inocula (100 μl) were added to each well in the microtitre plates. Medium alone added to the inoculum was used as a growth control; the blank control contained only the medium. The microtitre plates were then incubated at 35°C; the endpoints were read visually at 48 h. Amphotericin B (final concentration of 0.0156 to 16 μg/ml) and strain ATCC 90018 (C. parapsilosis) were used as quality controls.
The minimum fungicidal concentration (MFC) was determined by plating 10 μl from the wells showing no visible growth on SDA. The plates were incubated at 30°C for 48 h. The minimum concentration of essential oil that showed ≥99.9% reduction in the original inoculums was recorded as the MFC [24].
Preparation of Standard Yeast Cell Suspensions for Biofilm Assays
For biofilm growth, the protocol of Ramage et al. was followed [25]. Briefly, Candida were grown in SDA plates at 37°C for 18 h and then inoculated into yeast nitrogen base (YNB) medium (Difco) supplemented with 50 mM glucose. After overnight culture in a rotary shaker at 75 rpm, the yeast was harvested in the late exponential growth phase and washed twice with 20 ml of 0.1 M phosphate-buffered saline (PBS) pH 7.2. The suspensions were then diluted in RPMI as required to generate a final concentration of 0.5 × 106–2.5 × 106 cells/ml (counting with a haematocytometer).
Kinetics of Biofilm Formation on Microtitre Plates
Biofilms were formed in microtitre plates (Corning) over a series of time intervals (4, 6, 8, 12, 18, 24, 36, 48, 72, 84 or 96 h) at 37°C. At each time interval, biofilm production was quantified using an MTT reduction assay. For each time interval, four biofilm replicates were evaluated.
At overall time points, viable sessile cell counts were assessed. First, wells were extensively washed, and serial tenfold dilutions in sterile PBS were performed. The inoculum size for each cell suspension was confirmed by plating aliquots of 1:10, 1:100 and 1:1,000 dilutions on SDA (Difco) plates; CFU was obtained after a 48-h incubation period at 37°C. Results are expressed as log10 CFU/ml [17].
MTT analysis of replicate biofilms was performed in parallel with the total viable counts assay to demonstrate a correlation between these two techniques.
MTT Reduction Assay
Fungal viability was analysed using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT—Sigma) as described previously [16, 26]. Briefly, 100 μl of prewarmed MTT solution (0.5 mg/ml) in PBS containing 0.1% glucose was added to each well. The plates were incubated at 37°C for 6 h, and the MTT solution was then removed. Biofilms were washed once with PBS, and the formazan product was resuspended in acid isopropanol [5% (vol/vol) 1 M HCl in isopropanol]. Finally, the absorbance was measured (A 540nm). The minimum biofilm reduction concentration (MBRC) was defined as the concentration of essential oil showing A 540nm values equal to or lower than those of the control well that contained a biofilm treated with saline; the minimum biofilm fungicidal concentration (MBFC) was defined as the concentration with A 540nm values below or equal to the background level (acid isopropanol) [12, 27].
Scanning Electron Microscopy
For SEM, biofilms were formed on sterile PVC discs within 24-well microtitre plates (Corning) by dispensing 300 μl of standardised cell suspensions containing 2.5 × 106 cells/ml in RPMI 1640 onto the appropriate discs at 37°C [25]. Following biofilm formation, different CO concentrations (serial twofold dilutions ranging from 500 to 4,000 μg/ml) were added to selected wells, and the plates were incubated for 24 h at 37°C. For SEM observations, the biofilms were assayed as previously described by Priester et al. [28]. Briefly, the discs were removed, washed with PBS and placed in a fixative [4% formaldehyde (vol/vol), 1% glutaraldehyde (vol/vol) in PBS] solution overnight. The samples were rinsed 2 times for 3 min each in 0.1 M phosphate buffer, postfixed with 1% (w/v) OsO4 for 1 h, dehydrated using an ethanol gradient, critical point dried in CO2 (Critical Point Dryer, model CPD-030, Balzers, Oberkochen, Germany) and gold coated by sputtering (Denton Vacum, model Desk II, Freehold, NJ, USA). After processing, samples were observed with a scanning electron microscope (JEOL, model JSM 5410, Japan) in high vacuum mode at 15 kV. The images were processed for display using Photoshop software (Adobe Systems Inc., Mountain View, CA.).
Minimum Biofilm Inhibitory Concentrations
The effect of essential oils on C. parapsilosis complex biofilm formation was examined by the microbroth dilution method, similar to the MIC assay described above for planktonic cells (M27-A2). Essential oils (100 μl) diluted in RPMI to concentrations that ranged from 15.6 to 8,000 μg/ml were added to 100 μl of the inoculums suspension in 96-well polystyrene microtitre plates (Corning) and incubated for 48 h at 35°C [13, 29]. Controls containing inocula in RPMI and broth alone were included. The reference biofilm strain C. albicans SC 5314 was used as a control [25]. Following incubation, the culture supernatants from each well were aspirated, and the non-adherent cells were removed by washing the wells with sterile PBS. A semi-quantitative measure of biofilm formation was calculated using the MTT reduction assay.
Minimum Biofilm Reduction Concentrations and Minimum Biofilm Fungicidal Concentrations
Biofilms were grown as described above but without the presence of essential oils. After 24 h incubation at 37°C, the biofilms were washed three times with sterile PBS and exposed to 100 μl of essential oils, with oil concentrations ranging from 8,000 to 15.6 μg/ml in RPMI. The plates were then incubated for 48 h at 37°C, and the wells were washed once with sterile PBS and the fungal viability was determined using the MTT reduction assay [17, 29]. This assay was performed in duplicate microtitre plates. The reference strain C. albicans SC 5314 was used as a biofilm control [25].
Chequerboard Assays to Assess the Anticandidal Activity of Cinnamomum zeylanicum Essential Oil in Combination with Amphotericin B Against C. parapsilosis Complex Biofilms
The anticandidal activity of CO in combination with AMB was assessed by the chequerboard method [27, 30]. Microtitre plates containing C. parapsilosis complex biofilms were washed once with sterile PBS to remove any unbound cells. Serial double dilutions of the anticandidal compounds were prepared (16,000 μg/ml to 62.5 μg/ml for CO and 32 to 0.25 μg/ml for AMB) in RPMI. Next, 50 μl of the essential oil solutions was added to the rows of a 96-well microtitre plates in diminishing concentrations, and 50 μl of AMB was added to the columns in diminishing concentrations. Columns 10 and 11 contained biofilms and anticandidal compounds alone at various concentrations. Column 12 contained the biofilm and saline, and the saline alone controls. The microtitre plates were incubated at 37°C for 48 h. Fractional inhibitory concentrations (FICs) were calculated as the MIC of the combination of amphotericin B and CO divided by the MIC of CO or AMB alone. The FIC index (FICI) was calculated by adding both FICs and was interpreted in the following manner. A synergistic effect was reported when the FIC value was ≤0.5, no interaction was reported when the FIC value fell between 0.5 and 4.0 ranges, and an antagonistic effect was reported when the FIC value was >4.0 [27, 29]. Assays were performed in duplicate microtitre plates.
Data Analysis
Data analysis was performed using GraphPad Prism version 4.00 for Windows (GraphPad Software, San Diego, CA, USA). Descriptive statistics, such as the mean, standard deviation, median, upper and lower quartile, and highest and lowest value and the Pearson correlation coefficient, were used for statistical analysis. Comparisons of two groups were done using Mann–Whitney test, and a P value <0.05 was considered significant.
Results
C. parapsilosis Complex Identification
To identify the C. parapsilosis complex isolates, a fragment of the SADH gene (716 bp) was amplified by PCR (Fig. 1a) from genomic DNA samples. BanI restriction analysis of the SADH amplicons was used to identify C. orthopsilosis, C. metapsilosis and C. parapsilosis; C. orthopsilosis SADH amplicons do not contain a BanI restriction site, while C. metapsilosis amplicons contain three BanI restriction sites (at positions 96, 469 and 529), and C. parapsilosis amplicons contain only one BanI restriction site (at position 196). Using this technique, 53% of isolates were assigned to the most commonly encountered subtype, C. parapsilosis, and 47% of isolates were identified as C. orthopsilosis (Fig. 1-b). No C. metapsilosis isolates were identified in this study.

Discrimination of C. parapsilosis complex according to their SADH gene restriction profiles. (a) Amplification of a 716-bp SADH gene fragment from the genomic DNA from C. parapsilosis complex isolates to provide genetic confirmation. (b) Representative SADH gene restriction profile for C. orthopsilosis (lane 1) and C. parapsilosis (lanes 2–10) isolates. Lanes MM, 100-bp ladder
Quantification and Visualisation of Biofilms
The ability of Candida species to form biofilms was quantified by determining biofilm metabolic activity using the MTT assay and total viable cell counts. The kinetics of biofilm formation for both C. parapsilosis species and the C. albicans SC 5314 reference strain were similar; the MTT readings peaked at 48 h and declined thereafter (Fig. 2). When the Pearson correlation test was used to compare the spectrometric profile with the CFU counts, a significant correlation was found for C. parapsilosis, C. orthopsilosis and C. albicans (r = 0.940, r = 0.951, r = 0.958, respectively). Maximum cell density was observed at 48 h; decreased OD readings at 72 h suggest that the biofilms entered a declining growth phase at this time point (Fig. 2). SEM microscopy was used in conjunction with these semi-quantitative methods to monitor 24-h biofilm growth. Examination of single species biofilms showed the presence of different cellular morphologies in the biofilm structure. C. albicans SC 5314 produced a more profuse biofilm compared to the two C. parapsilosis species. C. orthopsilosis biofilms presented short and filamentous forms, while C. parapsilosis biofilms exhibited a blastospore aggregate layer with irregular clusters. In contrast, C. albicans developed a multilayer biofilm structure with a basal yeast layer that was covered by a relatively open network of hyphae penetrating the matrix (Fig. 3). SEM imaging of CO-treated Candida biofilms found that not only sessile Candida cells effectively are damaged by CO (Fig. 4) but that CO also detached biofilms (Fig. 4). These results indicate that CO has a dual mode of action against C. parapsilosis complex biofilms; it is able to detach adhering yeast from a substratum surface while also killing sessile cells.

Biofilm formation by C. parapsilosis (a), C. orthopsilosis (b) and C. albicans SC 5314 (c) growing on microtitre plates. Assays consisted of measurements of MTT formazan formation (dashed with dotted line) and enumeration of viable cells (solid line). Data represent the means ± standard errors of three independent experiments performed in duplicate. Error bars are omitted because the standard errors of the mean were significantly lower

Scanning electron micrographs (SEM) showing 24-h biofilms formed on PVC discs by different Candida species: (a) C. orthopsilosis, (b) C. parapsilosis and (c) C. albicans SC 5314. Samples were fixed prior to processing for SEM

Representative micrographs showing biofilms (24 h) formed by Candida orthopsilosis (a, d, e), C. parapsilosis (b, e, h) and reference strain C. albicans SC 5314 (c, f, i) after 48-h Cinnamomum zeylanicum oil treatment in different magnifications. Sessile Candida cells exhibited notable alterations in the membrane and the cell wall, besides a disordered profile of germination (a–f). Visualisation of the ultra structure in general revealed a marked reduction in biofilm constituents in all Candida species (g–i). Bars, a–e 5 μm; f 10 μm; g–i 50 μm
Effects of Essential Oils on C. parapsilosis Complex Planktonic Growth
The MIC and MFC of sixteen essential oils were evaluated in vitro against nine C. parapsilosis strains and eight C. orthopsilosis strains; all values are listed in Table 1. The MIC and MFC tests showed similar trends for the yeast isolates tested, where the MFC values were similar to or more than twofold greater than the MIC values. The C. parapsilosis complex was more susceptible to Cinnamomum zeylanicum oil, with MIC values between 250 and 500 μg/ml and MFC values between 500 and 1,000 μg/ml for C. orthopsilosis and C. parapsilosis, respectively.
Biofilm Susceptibility Assays
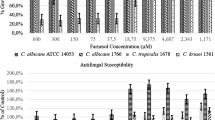
Essential oils from two plants, Juniperus virginiana and Cinnamomum zeylanicum, were selected for the evaluation of their antifungal activity on C. parapsilosis complex biofilm growth. The inhibitory effect on the biofilm formation by CO is depicted in Fig. 5. CO concentrations above 250 μg/ml effectively abolished C. parapsilosis complex biofilm formation, and in the reference strain, C. albicans SC 5314 was abolished at a CO concentration of 125 μg/ml. For the J. virginiana essential oil, higher concentrations (4,000 μg/ml) were required to inhibit biofilm formation (data not shown).

Effect of cinnamon oil on Candida biofilm formation (MBICs). Different cinnamon oil concentrations (7.81–8,000 μg ml) were added to Candida cells and incubated under biofilm growing conditions. The lines depict the per cent survival of Candida cells exposed to cinnamon oil. No statistically significant differences were found among the C. parapsilosis complex (Mann–Whitney test, P = 0.938). Assays consisted of measurements of MTT formazan formation; the metabolic activity was normalised to the control without oil, which was taken as 100%. All values are means and standard deviations of three independent experiments done in quadruplicate
The MBRC and MBFC values after treatment with CO were similar between C. orthopsilosis, C. parapsilosis and the C. albicans SC 5314 reference strain (1,000, 2,000 and 2,000 μg/ml, respectively) (Table 2). For C. orthopsilosis, the concentration necessary for the reduction of preformed biofilms (1,000 μg/ml, Table 2) was fourfold greater than the concentration required to inhibit biofilm formation (250 μg/ml, Fig. 5). Whereas for C. parapsilosis, the concentration necessary for the reduction of preformed biofilms (2000 μg/ml, Table 2) was eightfold greater than the concentration required to inhibit biofilm formation (250 μg/ml, Fig. 5). J. virginiana essential oil reduced C. parapsilosis complex or C. albicans SC 5314 biofilm growth at high concentrations (Table 2). Five per cent (v/v) DMSO, which was used as a cosolvent in the oil suspensions, did not show anticandidal activity against C. parapsilosis complex when grown in suspension or as a biofilm.
Chequerboard Assays to Assess the Anticandidal Activity of Cinnamon Oil in Combination with AMB Against C. parapsilosis Complex Biofilm Growth
The combination of CO with AMB demonstrated indifferent activity against C. parapsilosis and C. orthopsilosis, with an FICI value of 1.5 for both (Table 3). Indifference was also demonstrated with the combination against C. albicans SC 5314, with an FICI value of 1.5.
Discussion
The increasing incidence of hospital-acquired infections caused by drug-resistant fungal pathogens, host toxicity, the poor efficacy of drugs and high treatment costs has drawn attention to the potential of natural products as antifungals. Several studies have investigated the anticandidal effects of essential oils on Candida spp. [11, 31–35]. However, there is very limited information regarding the effect of essential oils on C. parapsilosis, either in planktonic or biofilm cultures.
The yeast species Candida parapsilosis, a nosocomial pathogen, exhibits both genomic and genetic heterogeneity. Recently, based on restriction polymorphism analysis of the SADH gene, which encodes a secondary alcohol dehydrogenase that is common to all species, C. parapsilosis complex isolates were distinguished into three species [5]. In this study, molecular analysis found that 53% of the isolates were C. parapsilosis, while 47% corresponded to C. orthopsilosis. Although substantial regional variation has been noted, most studies [36–39] found that C. orthopsilosis occurs in less than 10% of C. parapsilosis complex infections. However, a previous study [40] reported a C. parapsilosis complex isolate with 23.8% of C. orthopsilosis. This study reinforces a hypothesis that C. parapsilosis may be related to its ubiquitous nature because it is commonly isolated from different environmental sources.
When grown on a surface, C. parapsilosis adapts to a biofilm lifestyle [16]. Sessile, biofilm-associated cells can be distinguished from their planktonic, free-floating counterparts by their generation of an extracellular polymeric matrix, reduced growth rates and the up- and down-regulation of specific genes. In this study, the ability of C. parapsilosis complex to adhere to and form biofilms on polystyrene and PVC surfaces under static conditions was demonstrated. Biofilm growth was monitored by a colourimetric assay and total viable cell counts. Our quantitative results are in accordance with a previous study, which reported that C. parapsilosis complex exhibited an increased ability to form biofilms [41, 42].
Here, microscopy findings confirmed that C. albicans produces larger biofilms compared to C. parapsilosis species. Studies of non-albicans Candida species such as C. parapsilosis, which do not naturally form true hypha, suggest that hyphal morphology is not a prerequisite for biofilm formation [43–45]. Although the current data on C. parapsilosis complex biofilm corroborate these observations, a previous study [17] suggested that Candida hyphae contribute to biofilm architecture by forming a scaffold that provides robust support for exopolymeric substances and the blastospores to develop a three-dimensional, spatially organised community with several layers.
Selected natural products of plant origin can influence microbial biofilm formation through different mechanisms. Many plant essential oils, which are mixtures of numerous organic chemicals, contain compounds that inhibit microbial growth. In our study, the responses of C. parapsilosis complex biofilms to juniper oil and CO were measured by the MBRC and MBIC assays. The J. virginiana essential oil weakly inhibited the formation of biofilms and reduced preformed yeast biofilms. Alpha-pinene and myrcene are the predominate components (61.56%) of J. virginiana essential oil. It has been reported that, in yeast cells and isolated mitochondria, α-pinene and β-pinene destroy cellular integrity, inhibit respiration and ion transport and increase membrane permeability [46].
In assessing the anticandidal activity of essential oils, the difficulties in comparing results obtained experimentally with the existing data should be taken into account. The chemical composition of essential oils can vary within species for a variety of reasons, including the presence of chemotypes, the time of harvest and the use of different extraction methods [47]. Other factors that may explain our results include the absence of standardised anticandidal assays. Differences regarding experimental conditions, such as broth dilution or agar diffusion, culture medium composition, inoculum concentration, incubation time and temperature, prohibit strict comparisons [34].
In the present study, it was shown that CO exhibited anticandidal activity against both planktonic and biofilm cultures of environmental C. orthopsilosis and C. parapsilosis isolates; these results are consistent with other accounts where CO has shown effectiveness against other important pathogens [12, 34, 48]. Cinnamomum zeylanicum oil, which was predominately composed of cinnamaldehyde (54%), inhibited the growth of C. parapsilosis complex biofilms with low MBICs (250 μg/ml). A possible explanation for CO fungicidal activity is that the hydrophobicity of their volatile components allows for their insertion into the lipid bilayer of the cell membrane, disturbing it and increasing its permeability to protons. Subsequently, extensive leakage or the removal of critical molecules and ions eventually leads to microbial cell death [49]. This hypothesis is supported by the SEM results after CO treatment. Obtained images showed a marked decrease in cell aggregates and the complete absence of an extracellular matrix in Candida biofilms. Additionally, disruption resulting in the subsidence of the external portion of the cells was observed, indicating sessile cell damage. However, the susceptibility of C. parapsilosis complex to an essential oil derived from Cinnamomum zeylanicum has not been published for either planktonic organisms or for sessile cells.
Additionally, the MIC of CO for planktonic C. orthopsilosis and C. parapsilosis was similar to their MBIC values (250 and 500 μg/ml, 250 and 250 μg/ml, respectively). This suggests that CO has similar anticandidal activity against both planktonic and biofilm growth, which could be due to its action on cell membranes. CO and one of its main components, cinnamaldehyde, inhibit the proton motive force, respiratory chain, electron transfer and substrate oxidation in cells, resulting in the uncoupling of oxidative phosphorylation, inhibition of active transport, loss of pool metabolites and the disruption of DNA, RNA, protein, lipid and polysaccharide synthesis [12, 31, 50–52]. The main limitation of this study was the lack of evaluation in vivo anticandidal activities imposed by the complexity of the assays. Although CO and AMB are associated with activity in the plasma membrane and cell wall, suggesting the action potential, synergism was surprisingly not observed when CO and AMB were combined. It is possible that the synergistic interaction does not involve a single target or a single component [12]. It has been suggested that the diffusion of cationic compounds in biofilms is hindered by the negative charge of the extracellular matrix, which changes the physicochemical and tertiary structure of the biofilm. Because essential oils are composed of heterogeneous compounds and AMB is an amphoteric drug, it is possible that AMB treatment alters both the ionic interactions in the biofilm extracellular matrix and its mechanism of action against the extracellular matrix. However, further studies are needed to establish the mode of action of amphotericin B and CO in combination.
In conclusion, this study demonstrated that CO has excellent antifungal activity against biofilm or planktonic cultures of environmental C. orthopsilosis and C. parapsilosis isolates. CO was able to inhibit biofilm formation and kill yeast cells in C. parapsilosis complex biofilms. Importantly, biofilms were equally as sensitive to CO as their planktonic counterparts for C. orthopsilosis, mostly likely due to the similar activity of CO on yeast cell membranes. Further studies are required to elucidate the complex mechanism of action of CO and its components against biofilms formed by C. parapsilosis complex.
References
Pfaller MA, Messer SA, Boyken L, Tendolkar S, Hollis RJ, Diekema DJ. Geographic variation in the susceptibilities of invasive isolates of Candida glabrata to seven systemically active antifungal agents: a global assessment from the ARTEMIS Antifungal surveillance program conducted in 2001 and 2002. J Clin Microbiol. 2004;42:3142–6.
Mujica MT, Finquelievich JL, Jewtuchowicz V, Iovannitti CA. Prevalence of Candida albicans and Candida non-albicans in clinical samples during 1999–2001. Rev Argent Microbiol. 2004;36:107–12.
Nucci M, Queiroz-Telles F, Tobón AM, Restrepo A, Colombo AL. Epidemiology of opportunistic fungal infections in Latin America. Clin Infect Dis. 2010;51:561–70.
Medrano DJ, Brilhante RS, Cordeiro A, Rocha MF, Rabenhorst SH, Sidrim JJ. Candidemia in a Brazilian hospital: the importance of Candida parapsilosis. Rev Inst Med Trop Sao Paulo. 2006;48:17–20.
Tavanti A, Davidson AD, Gow NAR, Maiden MCJ, Odds FC. Candida orthopsilosis and Candida metapsilosis spp nov. To replace Candida parapsilosis groups II and III. J Clin Microbiol. 2005;43:284–92.
Baillie GS, Douglas JL. Effect of growth rate on resistance of Candida albicans biofilms to antifungal agents. Antimicrob Agents Chemother. 1998;42:1900–5.
Mah TC, O’Toole GA. Mechanism of biofilm resistance to anticandidal agents. Trends Microbiol. 2001;9:34–9.
Kuhn DM, George T, Chandra J, Mukherjee PK, Ghannoum MA. Antifungal susceptibility of Candida biofilms: unique efficacy of amphotericin B lipid formulations and echinocandins. Antimicrob Agents Chemother. 2002;46:1773–80.
Seneviratne CJ, Samaranayake LP. Biofilm lifestyle of Candida: a mini review. Oral Dis. 2008;14:582–90.
Cowan MM. Plant products as anticandidal agents. Clin Microbiol Rev. 1999;12:564–82.
Oliveira DR, Leitão GG, Santos SS, Bizzo DHR, Lopes D, Alviano CS, Alviano DS, Leitão SG. Ethnopharmacological study of two Lippia species from Oriximina, Brazil. J Ethnopharmacol. 2006;108:103–8.
Mesa-Arango AC, Montiel-Ramos J, Zapata B, Duran C, Betancur-Galvis L, Stashenko E. Citral and carvone chemotypes from the essential oils of Colombian Lippia alba (Mill.) N.E. Brown: composition, cytotoxicity and antifungal activity. Mem Inst Oswaldo Cruz. 2009;104:878–84.
Giordani R, Regli P, Kaloustian J, Mikaïl C, Abou L, Portugal H. Antifungal effect of various essential oils against Candida albicans. Potentiation of antifungal action of amphotericin B by essential oil from Thymus vulgaris. Phytother Res. 2004;18:990–5.
Shelef LA. Anticandidal effects of spices. J Food Saf. 1983;6:29–44.
Tavares AC, Gonçalves MJ, Cavaleiro C, Cruz MT, Lopes MC, Canhoto J, Salgueiro LR. Essential oil of Daucus carota subsp. halophilus: composition, antifungal activity and cytotoxicity. J Ethnopharmacol. 2008;119:129–34.
Nuryastuti T, van der Mei HC, Busscher HJ, Iravati S, Aman AT, Krom BP. Effect of cinnamon oil on icaA expression and biofilm formation by Staphylococcus epidermidis. Appl Environ Microbiol. 2009;75:6850–5.
Ali I, Khan FG, Suri KA, Gupta BD, Satti NK, Dutt P, Afrin F, Qazi GN, Khan IA. In vitro antifungal activity of hydroxychavicol isolated from Piper betle L. Ann Clin Microbiol Antimicrob. 2010;3:7–9.
Knobloch K, Pauli A, Iberl B, Weigand H, Weis N. Antibacterial and antifungal properties of essential oils components. J Ess Oil Res. 1989;1:119–28.
Alviano WS, Mendonça-Filho RR, Alviano DS, Bizzo HR, Souto-Padrón T, Rodrigues ML, Bolognese AM, Alviano CS, Souza MM. Anticandidal activity of Croton cajucara Benth linalool-rich essential oil on artificial biofilms and planktonic microorganisms. Oral Microbiol Immunol. 2005;20:101–5.
Dalleau S, Cateau E, Bergès T, Berjeaud JM, Imbert C. In vitro activity of terpenes against Candida biofilms. Int J Antimicrob Agents. 2008;31:572–6.
Thaweboon S, Thaweboon B. In vitro anticandidal activity of Ocimum americanum L. essential oil against oral microorganisms. Southeast Asian J Trop Med Public Health. 2009;40:1025–33.
Möller EM, Bahnweg G, Sandermann H, Geiger HH. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992;25:6115–6.
National Committee for Clinical Laboratory Standards. Reference method for broth dilution antifungal susceptibility testing of yeasts: approved standard, 2nd ed. NCCLS document M27–A2. Wayne, PA: National Committee for Clinical Laboratory Standards; 2002.
Canton E, Peman J, Viudes A, Quindos G, Gobernado M, Espinel-Ingroff A. Minimum fungicidal concentrations of amphotericin B for bloodstream Candida species. Diagn Microbiol Infect Dis. 2003;45:203–6.
Ramage G, Saville SP, Wickes BL, Lopez-Ribot JL. Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl Environ Microbiol. 2004;68:5459–63.
Krom BP, Cohen JB, Feser GEM, Cihlar RL. Optimized candidal biofilm microtiter assay. J Microbiol Methods. 2007;68:421–3.
Karpanen TJ, Worthington T, Hendry ER, Conway BR, Lambert PA. Anticandidal efficacy of chlorhexidine digluconate alone and in combination with eucalyptus oil, tea tree oil and thymol against planktonic and biofilm cultures of Staphylococcus epidermidis. J Antimicrob Chemother. 2008;62:1031–6.
Priester JH, Horst AM, Van de Werfhorst LC, Saleta JL, Mertes LA, Holden PA. Enhanced visualization of microbial biofilms by staining and environmental scanning electron microscopy. J Microbiol Methods. 2007;68:577–87.
Hendry ER, Worthington T, Conway BR, Lambert PA. Anticandidal efficacy of eucalyptus oil and 1,8-cineole alone and in combination with chlorhexidine digluconate against microorganisms grow in planktonic and biofilms cultures. J Antimicrob Chemother. 2009;64:1219–25.
Shin S, Lim S. Antifungal effects of herbal essential oils alone and in combination with ketoconazole against Trichophyton spp. J Appl Microbiol. 2004;97:1289–96.
Mastura M, Nor Azah MA, Khozirah S, Mawardi R, Manaf AA. Anticandidal and antidermatophic activity of Cinnamomum species oils. Cytobios. 1999;98:17–23.
Mondello F, De Bernardis F, Girolamo A, Salvatore G, Cassone A. In vitro and in vivo activity of tea tree oil against azole-susceptible and -resistant human pathogenic yeasts. J Antimicrob Chemother. 2003;51:1223–9.
Romano L, Battaglia F, Masucci L, Sanguinetti M, Posteraro B, Plotti G, Zanetti S, Fadda G. In vitro activity of bergamot natural essence and furocoumarin-free and distilled extracts, and their associations with boric acid, against clinical yeast isolates. J Antimicrob Chemother. 2005;55:110–4.
Pozzatti P, Scheid LA, Spader TB, Atayde ML, Santurio JM, Alves SH. In vitro activity of essential oils extracted from plants used as spices against fluconazole-resistant and fluconazole-susceptible Candida spp. Can J Microbiol. 2008;54:950–6.
Silva CB, Guterres SS, Weisheimer V, Schapoval EE. Antifungal activity of the lemongrass oil and citral against Candida spp. Braz J Infect Dis. 2008;12:63–6.
Lockhart SR, Messer SA, Tendolkar S, Diekema DJ. Geographic distribution and antifungal susceptibility of newly described species Candida orthopsilosis, in comparison to the closely-related species Candida parapsilosis. J Clin Microbiol. 2008;46:2659–64.
Gomez-Lopez A, Alastruey-Izquierdo A, Rodriguez D, Almilrante B, Pahissa A, Rodriguez-Tudela JL, Cuenca-Estrella M. Barcelona Candidemia Project Study Group. Prevalence and susceptibility profile of Candida metapsilosis and Candida orthopsilosis: results from population-based surveillance of candidemia in Spain. Antimicrob Agents Chemother. 2008;52:1506–9.
Silva AP, Miranda IM, Lisboa C, Pina-Vaz C, Rodrigues AG. Prevalence, distribution, and antifungal susceptibility profiles of Candida parapsilosis, C. orthopsilosis, and C. metapsilosis in a tertiary care hospital. J Clin Microbiol. 2009;47:2392–7.
Gonçalves SS, Amorim CS, Nucci M, Padovan AC, Briones MR, Melo AS, Colombo AL. Prevalence rates and antifungal susceptibility profiles of the Candida parapsilosis species complex: results from a nationwide surveillance of candidaemia in Brazil. Clin Microbiol Infect. 2010;16:885–7.
Tay ST, Na SL, Chong J. Molecular differentiation and antifungal susceptibilities of Candida parapsilosis isolated from patients with bloodstream infections. J Med Microbiol. 2009;58:185–91.
Lattif AA, Mukherjee PK, Chandra J, Swindell K, Lockhart SR, Diekema DJ, Pfaller MA, Ghannoum MA. Characterization of biofilms formed by Candida parapsilosis, C. metapsilosis, and C. orthopsilosis. Int J Med Microbiol. 2010;300:265–70.
Melo AS, Bizerra FC, Freymüller E, Arthington-Skaggs BA, Colombo AL. Biofilm production and evaluation of antifungal susceptibility amongst clinical Candida spp. isolates, including strains of the Candida parapsilosis complex. Med Mycol. 2011;49:253–62.
Nikawa H, Nishimura H, Hamada T, Makihira S, Samaranayake LP. Relationship between thigmotropism and Candida biofilm formation in vitro. Mycopathologia. 1998;144:125–9.
Seidler M, Salvenmoser S, Muller FM. In vitro effects of micafungin against Candida biofilms on polystyrene and central venous catheter sections. Int J Antimicrob Agents. 2006;28:568–73.
Thein ZM, Samaranayake YH, Samaranayake LP. In vitro biofilm formation of Candida albicans and non-albicans Candida species under dynamic and anaerobic conditions. Arch Oral Biol. 2007;52:761–7.
Andrews RE, Parks LW, Spence KD. Some effects of Douglas fir terpenes on certain microorganisms. Appl Environ Microbiol. 1980;40:301–4.
Mercier B, Prost J, Prost M. The essential oil of turpentine and its major volatile fraction α- and β-pinenes: a review. Int J Occup Med Environ Health. 2009;22:331–42.
Shan B, Cai YZ, Brooks JD, Corke H. Antibacterial properties and major bioactive components of cinnamon stick (Cinnamon burmannii): activity against foodborne pathogenic bacteria. J Agric Food Chem. 2007;55:5484–90.
Prabuseenivasan S, Jayakumar M, Ignacimuthu S. In vitro antibacterial activity of some plant essential oils. BMC Complement Altern Med. 2006;6:39.
Donlan RM, Costerton JW. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev. 2002;15:167–93.
Farag RS, Daw ZY, Hewedi FM, Elbatory GSA. Anticandidal activity of some Egyptian spice essential oils. J Food Prot. 1989;52:665–7.
Nychas GJE. Natural anticandidals from plants. In: Gould GW, editor. New methods of food preservations. London, UK: Blackie Academic; 1995. p. 58–89.
Acknowledgments
This work was supported by grants from Funded by Scientific Development Support Program of the School of Pharmaceutical Sciences at Unesp (PADC/FCFAr-UNESP proc. 2009/54) and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). We thank Prof. Arnaldo Lopes Colombo for C. albicans SC5314, C. parapsilosis ATCC 90018, C. orthopsilosis ATCC 96141 and C. metapsilosis ATCC 96143 strains.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pires, R.H., Montanari, L.B., Martins, C.H.G. et al. Anticandidal Efficacy of Cinnamon Oil Against Planktonic and Biofilm Cultures of Candida parapsilosis and Candida orthopsilosis . Mycopathologia 172, 453–464 (2011). https://doi.org/10.1007/s11046-011-9448-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11046-011-9448-0