
Abstract
On the family Brassicaceae, the causal agent responsible for downy mildew disease was originally regarded as a single species, Peronospora parasitica (now under Hyaloperonospora), but it was recently reconsidered to consist of many distinct species. In this study, 11 specimens of Peronospora drabae and P. norvegica parasitic on the genus Draba were investigated morphologically and molecularly. Pronounced differences in conidial sizes (P. drabae: 14–20 × 12.5–15.5 μm; P. norvegica: 20–29 × 15.5–22 μm) and 7.8% sequence distance between their ITS1-5.8S-ITS2 rDNA sequences confirmed their status as distinct species. Based on ITS phylogeny and morphology (monopodially branching conidiophores, flexuous to sigmoid ultimate branchlets, hyaline conidia and lobate haustoria), the two species unequivocally belong to the genus Hyaloperonospora and not to Peronospora to which they were previously assigned. Therefore, two new combinations, Hyaloperonospora drabae and H. norvegica, are proposed. The two taxa are illustrated and compared using the type specimen for H. norvegica and authentic specimens for H. drabae, which is lectotypified.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Downy mildew (Peronosporales, Oomycetes) is an economically destructive disease of brassicaceous crops in different geographical areas worldwide. After the first record of the disease on the family by Persoon [1] as Botrytis parasitica, Gäumann [2, 3] split it into as many as 52 species of Peronospora based on host specificity and simple morphological characteristics, which was widely accepted by Oomycete taxonomists. Subsequently, Gustavsson [4] mainly agreed with the narrow species concept of Gäumann, while Yerkes and Shaw [5] lumped all Peronospora species parasitic on the Brassicaceae under a single species, P. parasitica, which was widely followed by plant pathologists. Acknowledging the substantial morphological and molecular differences to Peronospora, Constantinescu and Fatehi [6] recently established the two genera Hyaloperonospora and Perofascia to accommodate the Peronospora species on Brassicaceae and allied families. They accepted six Hyaloperonospora species but still classified most of Hyaloperonospora specimens from numerous hosts as H. parasitica. Although they partially recognized that there are indeed several downy mildew pathogens on Brassicaceae, it was impossible to compare and segregate the numerous species of Gäumann [2, 3] based on morphological characteristics. The integrity of H. parasitica sensu Constantinescu and Fatehi [6] has been challenged by the advent of molecular phylogenetic analyses [7–12], which showed that there is a high degree of phylogenetic diversity, correlating with high host specificity, and that a narrow species concept is more suitable for taxonomy of Hyaloperonospora species.
Twenty-seven Peronospora species from the Brassicaceae have recently been transferred to Hyaloperonospora [6, 10–12], but Peronospora species from Draba remained unresolved in spite of their phylogenetic positions within Hyaloperonospora, possibly because of their unclear host ranges. No morphological examination was performed to distinguish these strains up to now. During a monographic work of the downy mildews of Korea, Shin and Choi [13, 14] collected many Draba nemorosa specimens infected with downy mildew and identified the causal pathogen as H. parasitica according to the concept of Constantinescu and Fatehi [6]. In the light of recent progress in the taxonomy of Hyaloperonospora, the Draba specimens previously assigned to H. parasitica were re-examined. As a result, two morphologically distinct Hyaloperonospora species were found to cause downy mildew on Draba nemorosa, each of which were similar to the descriptions given for P. drabae and P. norvegica. Subsequently, a comprehensive study using original and type specimens was undertaken to clarify the circumscription of the two species. As a result, both species are formally transferred to Hyaloperonospora, and H. drabae is lectotypified .
Materials and Methods
Oomycete Specimens
Eleven specimens of downy mildews on Draba (ten on Draba nemorosa and one on D. glabella) were phylogenetically and morphologically analysed in this study. For comparison, 26 sequences for ITS rDNA of Hyaloperonospora spp. were obtained from GenBank. To clarify the circumscription of P. norvegica and P. drabae, original and type specimens were obtained and examined. Information about the specimens is shown in Table 1.
Morphological Analysis
Herbarium specimens were moistened with 70% alcohol, and oomycetes were transferred to 60% lactic acid on a slide. The microscope preparations were warmed up, covered with coverslips and examined using an Olympus BX40 (Olympus, Tokyo, Japan) or a Zeiss AxioImager.A1 (Zeiss, Jena, Germany) microscope. Measurements were performed at 1,000× for conidia and ultimate branchlets and at 100–200× for other organs; they are reported as follows: (minimum)—standard deviation towards the minimum—standard deviation towards the maximum—(maximum).
DNA Extraction, PCR, Sequencing and Phylogenetic Analysis
Genomic DNA was extracted from conidiophores and conidia obtained from the lower surface of the infected leaves of herbarium specimens. DNA extraction was performed by the methodology described in Lee and Taylor [16]. For a 130-year-old herbarium specimen, the DNA extract was subsequently purified using the NucleoSpin Extract II kit (Macherey–Nagel, Düren, Germany). The rDNA region containing the partial 18S gene, both internal transcribed spacers (ITS1 and ITS2) and the 5.8S gene was amplified using the primers DC6 [17] and LR-0 [18]. For the extract from the 130-year-old herbarium specimen, primer pairs ITS5-P2—5.8S-Pr [19] and 5.8S-Pf (5′ GCAACTTTCAGCAGTGGATG 3′)—ITS4 [20] were used for amplification of two overlapping fragments containing the ITS1 and ITS2 regions, respectively. The PCR products were purified using the QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany) or an enzymatic PCR cleanup according to the protocol of Voglmayr and Jaklitsch [21]. Sequencing was performed on an ABI Prism TM 377 DNA Sequencer or an AB 3130xl Genetic Analyzer, using the BigDye™ (Applied Biosystems, Foster City, CA, USA) Cycle Sequencing Kit, version 3.1, with the primers ITS1, ITS2 and ITS3 [20] or 5.8S-Pf and 5.8S-Pr. Sequences were edited with the DNASTAR computer package (DNAStar, Inc., Madison, Wis.), version 5.05. Alignment of the sequences was first performed using MegAlign (DNAStar) and CLUSTAL X [22] and then visually checked and refined with Se-Al version 2.0 (A. Rambaut, University of Oxford, UK). Phylogenetic trees were obtained using maximum likelihood (ML), maximum parsimony (MP) and Bayesian methods (MCMC). For ML inference, RAxML version 7.0.3 [23] was used with all parameters set to default values, using the GTRCAT variant, and MP analysis was done using MEGA [24], version 4, with the default settings of the program, for which 1,000 bootstrapping (BS) replicates were performed [25]. Bayesian inference was performed with MRBAYES version 3.0b4 [26], which was used to obtain estimates for the posterior probabilities (PP) of groups. Perofascia lepidii (AY211011 & AY531467) was selected as outgroup taxon according to the results of recent multi-gene phylogenetic analyses [27].
Results
Morphological Analysis
Based on hyaline conidia, lobed haustoria and flexuous to sigmoid ultimate branchlets (Fig. 1), Peronospora drabae and P. norvegica unequivocally belong to the genus Hyaloperonospora, in contrast to Peronospora which is characterized by coloured conidia, hyphal haustoria and straight to curved ultimate branchlets. The conidiophores of the two species showed the more monopodial branching pattern and stout trunk commonly observed in Hyaloperonospora, whereas conidiophores in Peronospora are branching mostly dichotomously and have slender trunks. Peronospora drabae was most easily distinguished from P. norvegica by smaller conidia (15.2–18.1 × 12.6–14.2 μm vs. 22.1–26.4 × 17.3–20.2 μm). The oospore diameter (28.6–35.3 μm) of the former was also larger than that (25.5–31.5 μm) of the latter. In addition, the resting organs of P. drabae also differed from those of P. norvegica by somewhat brighter colour and thicker oospore walls.

Hyaloperonospora drabae (a–i) and H. norvegica (j–s). a, b, j, k: Conidiophores (bar 50 μm); c, d, m, n: Ultimate branchlets (bar 10 μm); l: Trunk; e–g and o–q: Conidia (bar 10 μm); h, i, r, s: Oospores (bar 20 μm). Sources: lectotype (BPI790248) for a–g, KUS-F24022 for h and i, holotype (O) for j–q, KUS-F21727 for r and s
Phylogenetic Analysis of ITS rDNA
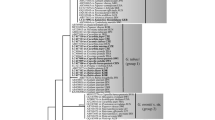
PCR products, including the partial 18S and the complete ITS1-5.8S-ITS2 regions, were amplified from each isolate. The 5′ end of the ITS1 and the 3′ end of the ITS2 sequences, which are significantly conserved, were determined and adjusted by comparison to annotated sequences of other Peronosporaceae. About twenty indels were found between the sequences of Peronospora drabae and P. norvegica, resulting in different total lengths of the complete ITS1-5.8S-ITS2 region of 791–792 bp in the former species and 797–798 bp in the latter. The phylogenetic relationships between downy mildew species on Brassicaceae were inferred from ML, MP and MCMC analyses of the aligned sequences of the ITS rDNA. The results of the phylogenetic reconstructions by ML inference are shown in Fig. 2. In the ITS rDNA alignment, 357 of the 850 characters were parsimony-informative, and the parsimony analysis produced 14 most parsimonious trees of 699 steps, with a consistency index (CI) and retention index (RI) of 0.6665 and 0.7243, respectively. Since no differences were found between the tree topologies of the ML, MP and MCMC analyses, only the ML tree is shown in Fig. 2, with the addition of the support values of the MP and MCMC analyses.

Phylogenetic tree inferred from maximum likelihood analysis of the complete ITS region (ITS1, 5.8S rDNA and ITS2). Support values (maximum likelihood bootstrap/maximum parsimony bootstrap/Bayesian posterior probabilities) above 50% are given above the branches. The number of nucleotide changes between taxa is represented by branch length and the scale bar equals the number of nucleotide substitutions per site. Hyaloperonospora specimens from Draba are shown in bold
In the ITS tree, the specimens from Draba spp. were clearly divided into two independent clades within Hyaloperonospora, with maximum support of 100/100/1.00 (ML BS/MP BS/MCMC PP) for both clades. One clade included seven specimens from D. nemorosa, whereas the other contained two from D. nemorosa and one from D. glabella, the former clade corresponding to Peronospora drabae and the latter to P. norvegica. Sequence distance between them was considerable, as about 7.8% (65 of 850 nucleotide characters) were different. P. drabae further formed a highly supported group with H. erophilae in the three analyses, with a sequence divergence of about 4% between the two species, whereas P. norvegica had no closer relative and was more distantly related to the other Hyaloperonospora species included in the phylogenetic analysis, showing sequence distances to other species of more than 6.5%.
Taxonomy
Due to the findings outlined above, it was concluded that the species described as Peronospora drabae and P. norvegica belong to the genus Hyaloperonospora. Therefore, two new combinations in Hyaloperonospora are proposed here.
Hyaloperonospora drabae (Gäum.) Y.J. Choi, H.D. Shin & Voglmayr, comb. nov. Fig. 1a–i
MycoBank # MB 518038
Basionym: Peronospora drabae Gäum., Beih. bot. Zbl., Abt. 1 35(1): 524 (1918)
Lesions commonly present on leaves, stems, peduncles and fruits, causing discolouration of the tissues and deformation of the attacked organs. Down whitish, consisting of scattered conidiophores to dense, felt-like. Haustoria lobate, up to 22 μm long, up to 10 μm wide, filling the host cell partly to almost completely. Conidiophores colourless, stout, straight, (105–)150–310(–420) μm long (n = 60); trunk mostly straight, (70–)120–205(–350) μm long (n = 65), variable in width, 8–14 μm wide below the first branch, 9–15 μm wide above the base, mostly with somewhat swollen base up to 18 μm wide; callose plugs mostly absent but very rarely present near the base; branching upper part monopodially or subdichotomously branched 5–7 times. Ultimatebranchlets in pairs or rarely single, flexuous to sigmoid, (9–)13–21(–25) μm long, 1.5–2.5 μm wide at the base (n = 56), apex obtuse. Conidia hyaline, subglobose to broadly ellipsoidal, (13.8–)15.2–18.1(–19.8) μm long, (12.5–)12.6–14.2(–15.5) μm wide, l/w ratio (1.10–)1.15–1.29(–1.35) (n = 88), greatest width median, base and tip round; pedicel absent in most conidia but a scar visible at the point of attachment; producing germ tubes. Restingorgans oogonia globose to irregular, golden or pale yellowish (43.1–)48.8–61.3(–64.5) μm diam. (n = 50), wall mostly smooth, 2.5–7 μm thick; oospores plerotic to aplerotic, globose, (24–)28.6–35.3(–37) μm diam. (n = 50), wall smooth, 2.5–4 μm thick.
Typus: Finland: Karelia, Soulajaroi, on leaves of Draba nemorosa (as “var. leiocarpa”) affected by downy mildew, June 1898, J. J. Lindroth (BPI790248; Sydow Phycomyceten et Protomyceten 215—lectotypus hic designatus).
Habitat: On living leaves of Draba nemorosa (Brassicaceae)
Other specimens examined: see Table 1. Additionally, four specimens were morphologically examined: Korea, Namyangju, Deokso, 15 Apr. 2009, Y. J. Choi & H. D. Shin (KUS-F24022); Korea, Yangpyeong, Experimental Forest of Korea University, 20 May 2009, Y. J. Choi & H. D. Shin (KUS-F24030); Korea, Hongcheon, Ecological Park, 29 May 2009, Y. J. Choi & H.D. Shin (KUS-F24031); USA, Colorado, Leadville, 8 Jun 1886,—(BPI790247; Mo. Bot. Gard. Herb. 67476).
Notes: In the original publication, Gäumann [2] listed three collections on which he based his description of Peronospora drabae, one from Draba nemorosa collected in Finland and two from Draba reptans (as D. caroliniana) collected in the USA. As he did not select a type, a lectotype has to be chosen from the authentic collections to ensure nomenclatural stability. We chose a specimen from D. nemorosa due to several reasons: The collection has been distributed among several herbaria as part of an exsiccatum, DNA data are available for accessions from D. nemorosa, whereas they are lacking for D. reptans, there are many more records of H. drabae from D. nemorosa, and Gäumann [2] included D. nemorosa but not D. reptans in his infection studies. From the several duplicates of Sydow Phycomyceten et Protomyceten 215 investigated during the present study from BPI, K, W and WU, the collection BPI790248 was selected as lectotype as the specimen agrees well with the description, contains sufficient material and is in good state of preservation. Whether the parasite from D. reptans is conspecific with H. drabae needs to be investigated by DNA data.
Hyaloperonospora norvegica (Gäum.) Y.J. Choi, H.D. Shin & Voglmayr, comb. nov. Fig. 1j–s
MycoBank # MB 518039
Basionym: Peronospora norvegica (as “norwegica”) Gäum., Beiträge zur Kryptogamenflora der Schweiz 5(4): 252 (1923)
Lesions commonly present on leaves, causing discolouration of the tissues and deformation of the attacked organs. Down whitish, usually sparse, consisting of scattered conidiophores, rarely dense and felt-like. Haustoria lobate, 14–22 μm long, 7.5–14 μm wide, filling the host cell partly to almost completely. Conidiophores colourless, stout, straight to slightly sinuous, (200–)230–355(–425) μm long (n = 30); trunk straight, (115–)130–225(–280) μm long (n = 30), variable in width, 10–15 μm wide below the first branch, 8–14 μm wide above the base, with mostly somewhat swollen base up to 18 μm wide; callose plugs absent; upper part monopodially or subdichotomously branched 5–7 times. Ultimatebranchlets in pairs or rarely single, flexuous to sigmoid, (8–)12–22(–32) μm long, 1.5–2.5 μm wide at the base (n = 55), apex obtuse. Conidia hyaline, broadly ellipsoidal to ellipsoidal, (20.3–)22.1–26.4(–28.9) μm long, (15.5–)17.3–20.2(–21.7) μm wide, l/w ratio (1.15–)1.22–1.37(–1.5) (n = 50), greatest width median, base and tip round; pedicel absent in most conidia but a scar visible at the point of attachment; producing germ tubes. Restingorgans oogonia globose to irregular, yellowish to brown, (37.5–)43.5–51.0(–62.5) μm diam. (n = 50), wall mostly smooth, 2.5–6 μm thick; oospores plerotic to aplerotic, globose, (22–)25.5–31.5(–32.5) μm diam. (n = 50), wall 1.5–2 μm thick, smooth, yellowish.
Typus: Norway: Sør-Trøndelag, Oppdal, Dovre, Kongsvold, on leaves of Draba glabella (as D. hirta) affected by downy mildew, 8 August 1880, A. Blytt (O—holotypus).
Habitat: On living leaves of Draba glabella and D. nemorosa (Brassicaceae)
Other specimens examined: see Table 1.
Discussion
The most useful morphological characteristics for distinguishing between Hyaloperonospora and Peronospora are the branching pattern, shape of haustoria, colour of conidia and the shape of ultimate branchlets [6, 28]. In the present study, H. drabae and H. norvegica clearly had a monopodial branching pattern, lobed haustoria, flexuous to sigmoid ultimate branchlets and hyaline conidia, showing that they unequivocally belong to Hyaloperonospora but not to Peronospora. In the ITS-based phylogenetic analyses, the two species were also clearly placed within Hyaloperonospora (Fig. 2).
An easy way to distinguish Hyaloperonospora drabae and H. norvegica is by their different conidial sizes, which never overlapped. The suitability of conidial size for species discrimination is not confined to Hyaloperonospora but has also been shown for related genera. Hamilton and Cunnington [29] showed that precise statistical estimates of spore dimensions give higher resolution for Gäumann’s Peronospora species, and Choi et al. [30–33] demonstrated that the conidial size and shape are useful characteristics to distinguish among Peronospora species. Similarly, Gustavsson [4] recognized that H. drabae and H. norvegica show significant differences in conidial size. Phylogenetic analyses of ITS sequence data revealed species-specific indels and high genetic distances between the two species. Therefore, the results support their status as two distinct and distantly related species within Hyaloperonospora.
After Gäumann first described Hyaloperonospora drabae from D. reptans and D. nemorosa [2] and H. norvegica from D. glabella [3], the former species was recorded in several monographic studies [34–36], in which the morphological characteristics perfectly fit the present descriptions. However, the spore sizes given for Peronospora drabae in Novotelnova and Pystina [37] (11–24 × 9–21) indicate that their description could be based on a mixture of both H. drabae and H. norvegica. In contrast to the widely distributed H. drabae, to our knowledge, H. norvegica has not been recorded for about 90 years. Before the species was described, only Rouppert [38] recorded similar measurements of conidia (ca. 22.9 × 18.3 μm) and oospores (23–31 μm) for P. parasitica on D. nemorosa. Compared with H. drabae, the few records of H. norvegica seem to be linked to its inconspicuous disease symptoms. Although the two species were collected from the same host plant, the symptoms were substantially different. In H. drabae, it was easily observed as a whitish felt-like down over leaves, stems, peduncles and fruits, whereas in H. norvegica, it was hardly visible in the field, as conidiophores were scattered and sparse and could be observed only when viewed under a dissecting microscope.
Interestingly, H. drabae and H. norvegica were found to co-exist on D. nemorosa within a limited geographical area in South Korea. This is remarkable, as in the highly host-specific downy mildews, usually only a single species is known from a specific host. Voglmayr et al. [39] record a similar case of sympatric co-occurrence of two distinct species on the same host species and even on the same host individual, for the distantly related Plasmopara geranii-sylvatici and P. praetermissa. Likewise, for the distantly related Albuginaceae, Thines et al. [40] recorded two distinct species of Albugo from Arabidopsis thaliana under the same environmental conditions. This indicates that sympatric co-occurrence of more than one species on the same host may be more common than previously expected but has remained unnoticed due to the lack of thorough investigations. These species pairs should be investigated in detail, as they may provide important insights into the evolutionary diversification and niche adaptation processes of obligate plant parasites.
References
Persoon CH. Observationes mycologicae. Vol. 1. Leipzig: P.P. Wolf; 1796.
Gäumann E. Über die Formen der Peronospora parasitica (Pers.) Fries. Ein Beitrag zur Speziesfrage bei den parasitischen Pilzen. Beih Bot Centralb. 1918;34(1):395–533.
Gäumann E. Beiträge zu einer Monographie der Gattung Peronospora Corda. Beitr Kryptogamenfl Schweiz. 1923;5(4):1–360.
Gustavsson A. Studies on nordic peronosporas. I. Taxonomic revision. Opera Bot. 1959;3(1):1–271.
Yerkes WD, Shaw CG. Taxonomy of the Peronospora species on Cruciferae and Chenopodiaceae. Phytopathology. 1959;49:499–507.
Constantinescu O, Fatehi J. Peronospora-like fungi (Chromista, Peronosporales) parasitic on Brassicaceae and related hosts. Nova Hedwigia. 2002;74:291–338.
Riethmüller A, Voglmayr H, Göker M, Weiß M, Oberwinkler F. Phylogenetic relationships of the downy mildews (Peronosporales) and related groups based on nuclear large subunit ribosomal DNA sequences. Mycologia. 2002;94:834–49.
Choi YJ, Hong SB, Shin HD. Diversity of Hyaloperonospora parasitica complex from core brassicaceous hosts based on ITS rDNA sequence. Mycol Res. 2003;107:1313–21.
Voglmayr H. Phylogenetic relationships of Peronospora and related genera based on nuclear ribosomal ITS sequences. Mycol Res. 2003;107:1132–42.
Göker M, Voglmayr H, Riethmüller A, Weiß M, Oberwinkler F. Taxonomic aspects of Peronosporaceae inferred from Bayesian molecular phylogenetics. Can J Bot. 2003;81:672–83.
Göker M, Riethmüller A, Voglmayr H, Weiss M, Oberwinkler F. Phylogeny of Hyaloperonospora based on nuclear ribosomal internal transcribed spacer sequences. Mycol Prog. 2004;3:83–94.
Göker M, Voglmayr H, García–Blázquez G, Oberwinkler F. Species delimitation in downy mildews: the case of Hyaloperonospora in the light of nuclear ribosomal ITS and LSU sequences. Mycol Res. 2009;113:308–25.
Shin HD, Choi YJ. A first check-list of Peronosporaceae from Korea. Mycotaxon. 2003;86:249–67.
Shin HD, Choi YJ. Peronosporaceae of Korea. Suwon, Korea: National Institute of Agricultural Science and Technology; 2006.
Holmgren PK, Holmgren NH. 1998. Index Herbariorum. New York Botanical Garden. http://sciweb.nybg.org/science2/indexHerbariorum.asp.
Lee SB, Taylor JW. Isolation of DNA from fungal mycelia and single spores. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editors. PCR protocols: a guide to methods and applications. San Diego: Academic Press; 1990. p. 282–7.
Cooke DEL, Drenth A, Duncan JM, Wagels G, Brasier M. A molecular phylogeny of Phytophthora and related Oomycetes. Fungal Genet Biol. 2000;30:17–32.
Moncalvo JM, Wang HH, Hseu RS. Phylogenetic relationships in Ganoderma inferred from the internal transcribed spacer and 25S ribosomal DNA sequences. Mycologia. 1995;87:223–38.
Voglmayr H, Constantinescu O. Revision and reclassification of three Plasmopara species based on morphological and molecular phylogenetic data. Mycol Res. 2008;112:487–501.
White TJ, Bruns T, Lee S, Taylor JW. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editors. PCR protocols: a guide to methods and applications. San Diego: Academic Press; 1990. p. 315–22.
Voglmayr H, Jaklitsch W. Prosthecium species with Stegonsporium anamorphs on Acer. Mycol Res. 2008;112:885–905.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997;24:4876–82.
Stamatakis E. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006;22:2688–90.
Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol. 2007;24:1596–9.
Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39:783–91.
Ronquist F, Huelsenbeck JP. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–4.
Göker M, Voglmayr H, Riethmüller A, Oberwinkler F. How do obligate parasites evolve? A multi-gene phylogenetic analysis of downy mildews. Fungal Genet Biol. 2007;44:105–22.
Thines M. Evaluation of characters available from herbarium vouchers for the phylogeny of the downy mildew genera (Chromista, Peronosporales), with focus on scanning electron microscopy. Mycotaxon. 2006;97:195–218.
Hamilton AJ, Cunnington JH. Calculating minimum sample sizes for taxonomic measurements: examples using Gäumann’s Peronospora spore data. Mycotaxon. 2006;95:189–94.
Choi YJ, Constantinescu O, Shin HD. A new downy-mildew of the Rosaceae: Peronospora oblatispora sp. nov. (Chromista, Peronosporales). Nova Hedwigia. 2007;85:93–101.
Choi YJ, Hong SB, Shin HD. Re-consideration of Peronospora farinosa infecting Spinacia oleracea as distinct species, Peronospora effusa. Mycol Res. 2007;111:381–91.
Choi YJ, Denchev CM, Shin HD. Morphological and molecular analyses support the existence of host-specific Peronospora species infecting Chenopodium. Mycopathologia. 2008;168:155–64.
Choi YJ, Shin HD, Thines M. Two novel Peronospora species are associated with recent reports of downy mildew on sages. Mycol Res. 2009;113:1340–50.
Kochman J, Majewski T. Grzyby (Mycota). Tom IV. Glonowce (Phycomycetes), Wroslikowe (Peronosporales). Państwowe Wydawnictwo Naukowe: Warzawa; 1970.
Ul’yanishchev VI, Osipian LL, Kanchaveli LA, Akhundov TM. Peronosporovye griby (Peronosporaceous Fungi). In: Ul’yanishchev VI, editor. Opredelitel’ gribov zakavkaz’ya: Erevan University, Erevan; 1985.
Staneviciené S. Peronosporales. In: Mazelaitis J, Staneviciené S, editors. Lietavos Grybai. I. Gleivunai (Myxomycota), Peronosporieciai (Peronosporales) 1. Vilnius: Mokslo ir enciklopediju leidykla; 1995. p. 85–291.
Novotelnova NS, Pystina KA. Ordo Peronosporales (Fam. Pythiaceae, Phytophthoraceae, Peronosporaceae, Cystopaceae). In: Gorlenko MV, editor. Cryptogamic flora of URSS, vol, 11, Fungi (3). Leningrad: Nauka; 1985.
Rouppert K. Przyczynek do znajomości grzybów Galicyi i Bukowiny. Kosmos, Lwów. 1911;36:936–44.
Voglmayr H, Fatehi J, Constantinescu O. Revision of Plasmopara (Chromista, Peronosporales) parasitic on Geraniaceae. Mycol Res. 2006;110:633–45.
Thines M, Choi YJ, Kemen E, Ploch S, Holub EB, Shin HD, Jones JDG. A new species of Albugo parasitic to Arabidopsis thaliana reveals new evolutionary patterns in white blister rusts (Albuginaceae). Persoonia. 2009;22:123–8.
Acknowledgments
The authors are grateful to the herbarium curators of the U.S. National Fungus Collections (BPI), the Royal Botanic Gardens Kew (K), the Botanical Museum Oslo (O), the Naturhistorisches Museum Wien (W) and the Universität Wien (WU) for providing the Peronospora specimens. Financial support via a research grant from the Korea University to H. D. S. in 2007 and a EU SYNTHESYS grant (GB-TAF-4625) to H. V. for a research stay at the Royal Botanic Gardens Kew are gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Choi, YJ., Shin, HD. & Voglmayr, H. Reclassification of Two Peronospora Species Parasitic on Draba in Hyaloperonospora Based on Morphological and Molecular Phylogenetic Data. Mycopathologia 171, 151–159 (2011). https://doi.org/10.1007/s11046-010-9340-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11046-010-9340-3