
Abstract
Sheep are valuable resources for the wool industry. Wool growth of Aohan fine wool sheep has cycled during different seasons in 1 year. Therefore, identifying genes that control wool growth cycling might lead to ways for improving the quality and yield of fine wool. In this study, we employed Agilent sheep gene expression microarray and proteomic technology to compare the gene expression patterns of the body side skins at August and December time points in Aohan fine wool sheep (a Chinese indigenous breed). Microarray study revealed that 2,223 transcripts were differentially expressed, including 1,162 up-regulated and 1,061 down-regulated transcripts, comparing body side skin at the August time point to the December one (A/D) in Aohan fine wool sheep. Then seven differentially expressed genes were selected to validated the reliability of the gene chip data. The majority of the genes possibly related to follicle development and wool growth could be assigned into the categories including regulation of receptor binding, extracellular region, protein binding and extracellular space. Proteomic study revealed that 84 protein spots showed significant differences in expression levels. Of the 84, 63 protein spots were upregulated and 21 were downregulated in A/D. Finally, 55 protein points were determined through MALDI-TOF/MS analyses. Furthermore, the regulation mechanism of hair follicle might resemble that of fetation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
DNA microarray analysis has been proved to be an effective method in investigating the characteristics of hair follicle stem cells in mice [1–4]. A serial of skin-expressed microRNAs with possible roles in goat and sheep wool growth has also been described [5]. Yu et al. [6, 7] testified the effectiveness of cDNA chip for expression profiling of wool growth cycling in whole skin. Transcriptomic studies on sheep and goats have been reported, related to different traits, such as wool follicle development [8], pigmentation traits of skin and wool [9], natural fleece rot resistance [10], mammary development and milk quality [11–13], and resistance to parasites [14, 15]. Comparative transcriptomic analysis of the primary and secondary follicles of a cashmere goat reveal 51 genes that are differentially expressed between the two types of hair follicles [16].
Fine wool sheep provides high quality wool for the industry. Identifying genes that regulate wool growth cycling helps to find a way to improve the quality and yield of wool products in breeding programs. Aohan fine wool sheep, bred in Inner Mongolia, is an excellent breed, supplying both fine wool and meat. The advantages of this breed is high-quality wool, high resistance to disease and strong ability to adapt. Ultimately, Aohan fine wool sheep is a preferable breed for fine wool production in China. Wang et al. [17] demonstrated that climate factors displayed appreciable impact on wool growth of Aohan fine wool sheep. The peak of growth rate is in summer and the low in winter [17]. That is to say, the wool growth has cycled during different seasons in a year. The expression profiling of immune genes and type I inner root sheath (IRS) keratin genes in the whole skin of Aohan fine wool sheep has been described previously by our lab [18, 19].
Hair follicle is a regenerating system. The follicle demonstrates the unusual ability to completely regenerate itself. The basis for this regeneration rests in the unique follicular epithelial and mesenchymal components and their interactions [20]. Yu et al. studied the wool follicle cycling through adjusting the artificial illumination time [6, 7]. That is only part of the explanation, however. After all, natural lighting could provide a preferable answer.
As far as we know, no microarray or proteomic study at a genome-wide level has been conducted on genes that were possibly responsible for controlling wool growth cycling of adult sheep at natural conditions to date. The purpose of this study was to investigate and compare the gene expression level of body side skin at different time points through gene chip and proteomic technology, and to search and select the candidate genes and proteins responsible for wool growth cycling control of Aohan fine wool sheep.
Materials and methods
Animals and sample preparation
All animals were handled according to the animal processing protocols approved by national and local animal welfare agencies, and all animal experiments was authorized by the Shandong Province Biological Studies Animal Care and Use Committee.
Sampling methods were described previously [18, 19, a paper submitted to BMC Genetics]. One ram and two ewes were used in the cDNA microarray study. These animals were half sibs (sharing the same father). In August and December time points of 2010, whole skin from body side area (wool bearing) were picked up from the three animals under local anaesthesia for cDNA microarray and proteomic experiments. All samples were put into liquid nitrogen immediately after collection. The area of each skin sample was approximately one square centimetre. There were 15, 208 probes in Agilent Sheep Gene Expression Microarray (Santa Clara, CA, USA).
RNA extraction and microarray hybridization
Total RNA was extracted from the skin samples using TRIzol reagent (Invitrogen) following the manufacturer’s protocols. RNA concentration was assessed with a NanoDrop spectrophotometer (NanoDrop Technologies) and RNA integrity was verified using an Agilent 2100 bioanalyzer (Agilent). The RNA samples were then incubated with the Agilent Sheep Gene Expression Microarray (Santa Clara, CA, USA). Each RNA sample was hybridized to one microarray slide (one-to-one). 1 μg of total RNA from each sample was doubled and transcribed into fluorescent cRNA using the manufacturer’s Agilent’s Quick Amp Labeling protocol (version 5.7, Agilent Technologies). The marked cRNAs were added onto the Whole Genome Oligo Array (4×44K, Agilent Technologies). All the microarray experiments were performed by Kangchen Biotechnology Limited Company (Shanghai, China).
Microarrays data analysis
After hybridization and washing, the microarray slides were scanned with the Agilent Scanner G2505B. The resulting text files derived from Agilent Feature Extraction Software (version 10.5.1.1) were introduced into the Agilent GeneSpring GX software (version 11.0) for subsequent calculation. Gene expression levels were standardized relative to the level of GAPDH. Differentially expressed genes were found out through fold-change screening. The fold-change of 2.0 were set up as the threshold. The statistical analysis was performed with Student’s t test and then the P-values were corrected for multiple testing. The false discovery rate was set as approximately 20 %, in order to get more differentially expressed genes for further analysis. The functional annotation of differentially expressed genes was performed by the Database for Annotation, Visualization and Integrated Discovery (DAVID) gene annotation tool (http://david.abcc.ncifcrf.gov/) [21].
qPCR
The total RNA samples prepared for microarray detection were also used for qPCR experiments. Reverse transcriptions were performed using M-MLV Reverse Transcriptase (Promega) according to the manufacturer’s protocols. The primers were designed using the Primer Premier 5 software. The primer sequences, melting temperatures and product sizes are shown in Table 1. GAPDH was used as the house keeping gene.
Tissue protein extraction
Lysis buffer was prepared using ddH2O as dissolvent as follows: 42 % urea, 15.2 % thiourea, 4 % CHAPS, 1 % DTT. The buffer was usually separated into 1 ml volume and stored at −20 °C. The skin samples were cut into small pieces by scissors, and left at 4 °C for 1 h, vortexed it every 15 min. Sampled tissues were homogenized in lysis buffer (containing 1 % cocktail and 2 % IPG-buffer, added right before use) at the ratio of 1:7 (w/v). Then, the tissue homogenate was centrifuge at 40,000 g for 30 min. Supernatants were saved and stored at −70 °C. The protein concentration in each sample was determined by the bicinchoninic acid (BCA) method using PBS as the standard.
Two-dimensional electrophoresis (2-DE)
The electrophoresis programme is as follows:
-
Transfer: voltage 300 V, current 50 mA, time 1 h.
-
Separation: voltage 300 V, current 200–250 mA, time 4–5 h.
-
Fixative preparation: 40 % ethanol and 10 % acetic acid.
-
Fixation: take out the rubber strip and put it into fixative for 1 h.
Image analysis
Gels were silver stained, scanned and analyzed using ImageMaster TM 2D platinum software (Version 5.0, GE Healthcare, San Francisco, CA, USA). The expression level was determined by the relative volume of each spot in the gel and expressed as %Vol (%Vol = [spot volume/Σvolumes of all spots resolved in the gel]). The means and standard deviations of both sample groups were calculated. Statistical significance with Student’s t tests using ImageMaster TM 2D platinum software. The P values obtained with the t test were corrected for multiple testing. The false discovery rate was set as approximately 20 %, in order to get more differentially expressed proteins for further analysis.
Protein identification
Protein spots were excised, dehydrated in acetonitrile, and dried at room temperature. Proteins were reduced and alkylated. Gel pieces were denatured, alkylated, trypsin digested and analyzed by the Ultraflex II MALDI-TOF-TOF mass spectrometer (Bruker Daltonics GmbH, Bremen, Germany) under the control of FlexControl TM 2.4 software (Bruker Daltonics GmbH). Acquired peptide mass fingerprint (PMF) were analysed with the program FlexAnalysis™ 3.0 (Bruker Daltonics, Bremen, Germany). The peak detection algorithm was: SNAP (Sort Neaten Assign and Place); S/N threshold: 1.5; Quality Factor Threshold: 50. The tryptic auto-digestion ion picks (trypsin [108–115] 842.51 Da, trypsin [58–77] 2211.10 Da) were used as internal standards to validate the external calibration procedure. Matrix and/or auto-proteolytic trypsin fragments and known contaminant ions, were excluded. The derived peptide mass lists were used to aligning the Matrixscience database (http://www.matrixscience.com). The following search parameter criteria were used: mass tolerance 100 ppm, miss cleavage ≤1, modification comprises carbamidomethyl and methionine oxidation. Matched peptides number between experimental PMF and theoretical PMF ≥5 [22].
Results
Summary of differentially expressed genes in microarray analysis
Two thousand two hundred and twenty-three annotated transcripts were differentially expressed between the two time points, including 1,162 up-regulated and 1,061 down-regulated transcripts, as shown in Table S1 (Supplementary material). 24 genes were up-regulated more than 10 folds, while 16 genes downregulated more than 10 folds, in August time point versus December time point (A/D).
Furthermore, as shown in Table S2 (Supplementary material), many gene families that regulate different aspects of hair follicle growth and cycling show differential expression in A/D, such as growth factors, immune genes, enzymes,pregnancy-associated glycoprotein, and so on.
Biological process gene ontology (GO) analysis
Based on the DAVID, scores of genes classified into 9 categories (Table S3 in Supplementary material). The majority of the genes possibly related to follicle development and wool growth could be assigned into the categories including regulation of receptor binding, extracellular region, protein binding and extracellular space.
Selective verification for microarray data by qPCR
To confirm the data on the transcriptional level, we picked up 7 DE genes, including FGFR2, IGFBP2, Connexin 43, ZO1, HSP70.1, IL8 and YY1 to comparatively detect their expression patterns by real time PCR. As illustrated in Fig. 1, the qPCR results for the selected 6 genes were consistent with the microarray data except IGFBP2, which verified the credibility of the microarray data.

Q-PCR validation of the microarray data. P values (T-test) of the Q-PCR data are 0.021 (FGFR2), 0.157 (IGFBP2), 0.010 (Connexin 43), 0.023 (ZO1), 0.035 (HSP70.1), 0.042 (IL8) and 0.026 (YY1), respectively. S1/S2 represent body side skin at the August and December time points, respectively
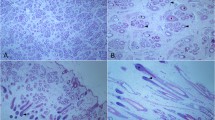
Identification of DE protein spots on 2-DE gels
To detect differential protein expression between the two groups, triplicate 2D maps of protein samples for each group were created. Figure 2 shows two representative 2-DE gel images of the protein expression patterns of the two groups. 84 protein spots showed significant differences in expression levels between the two groups. Of the 84, 63 protein spots were upregulated and 21 were downregulated in A/D. Some of them could not be identified causing by incomplete polypeptide fragments or low abundance (beyond the detection limit). Finally, 55 protein points were determined through MALDI-TOF/MS analyses. These DE proteins is displayed in Table S4 (Supplementary material).

Representative image of 2-DE silver stained polyacrylamide gel
Discussion
Significance of differentially expressed genes at the transcriptional level
In this study, we investigated the differentially expressed genes and proteins possibly regulating wool growth and cycling using cDNA microarray and proteomic technologies. Gene chip analysis identified hundreds of differentially expressed genes showing more than twofold difference between the two time points: August and December. While upregulated transcripts (1,162) in A/D was a little more than that of the downregulated ones (1,061). The qPCR experiments verified the reliability of our microarray results.
IL-1A and IL-1B inhibit hair growth in vitro as published previously [20]. However, IL-1B has shown upregulation at the August time point in our study. Wool was supposed to grow faster during August than in December.
A log linear linkage exists between the relative level of ITGB1 on the cell surface and proliferative capacity in keratinocytes [23]. ITGB1 signalling is also essential for human hair growth [24]. Skin and hair follicle integrity is crucially dependent on ITGB1 expression in keratinocytes [25]. In our study, however, ITGB1 expression in A/D was down-regulated. And ITGB1 level was also lower in wool bearing area and in wool-less area in another study (data not shown). The reasons how those happen deserve further investigation.
During development process, GluD1 was distributed to the hippocampus, cochlear and vestibular hair cells, and spiral ganglion cells [26–28]. Hair follicles are also derived from ectoderm. So why GLUD1 showed down-regulated at the transcription level in A/D remains to be further investigated.
Connexin 43 is a gap junction protein distributed in the follicular dermal papilla [28]. A recent study by mouse model suggested Connexin 43 plays vital roles in hair regeneration, growth, and cuticle formation [29]. However, Connexin 43 displayed downregulation in A/D in the transcriptional level of our study, and it was subsequently validated by real time qPCR.
We found that a high percentage of DE genes play important roles during embryo development. As displayed in Table S2 (Supplementary material), 3 pregnancy-associated glycoproteins (PAG 6, PAG 9 and PAG 11) were up-regulated in A/D. Furthermore, we found dozens of immune-associated genes from the DE gene list, such as heat shock proteins, interleukins, matrix metallopeptidases and clusters of differentiation. Similar phenomena were found in another study (data submitted to BMC Genetics). The immune privilege mechanism exists both in hair follicle cycling and embryo development. Therefore, in a sense, hair follicles revealed similar features with the embryo, and that the regulation mechanism of hair follicle might resemble that of fetation.
Significance of differentially expressed proteins in the proteomic study
Eighty-four protein spots were differentially expressed. Of the 84, 63 protein spots were upregulated and 21 were downregulated in A/D. 55 protein points were determined through MALDI-TOF/MS analyses. Most of the DE proteins were not discussed above.
Cryab protein was detected in 2DE in August time point but not in December time point. However, the Cryab transcripts show no differential expression in the microarray level. This gene, downregulated in the transcriptional level during the depilation-induced hair cycle [30], was considered to play a role in regulation of apoptosis. So our results support this inference.
KRT5, KRT14, KRT2.11 (KRT86) and KRT19 proteins were all upregulated in A/D. These proteins were reported to play important roles in wool follicles [31–35]. KRT14 and KRT19 are type I keratin, while KRT2.11 (KRT86) and KRT5 are type II keratin [16, 32]. In secondary versus primary follicles of Yunnan black goat, the two differentially expressed keratin genes (keratin 40 and 72) were type I and type II, respectively [16]. So similar mechanism or related biological process might exist between wool cycling and cashmere development.
FGFR3 RNA distributed in precuticle cells in the periphery of the hair bulb [36, 37]. Consistently, FGFR3 protein was upregulated in August time point compared to the control. We could deduced that FGFR3 may play positive roles in wool growth.
Concordance between the results of transcriptomic and proteomic levels
In the results of transcriptomic level (microarray), the number of up-regulated genes was higher than that of down-regulated ones (1,162 vs. 1,061). Consistently, up-regulated protein spots were more than down-regulated ones (63 vs. 21).
The concordance between these two results from microarray and proteomic experiments was shown in Table S4 (Supplementary material). Of the 55 identified proteins, 5 corresponding transcripts also shown differential expression. Among these 5 transcripts (CNN1, HSP70.1, OVGP1, TYR and Flt-1), the regulation trends of 4 were consistent with their protein counterparts except HSP70.1. The inconsistency of the two levels of HSP70.1 expression might be produced by post transcriptional regulation.
Conclusion
In summary, the results in this research suggested that the body side skin at August time point displayed a differentially expressed pattern in comparison with that at December time point. Most of these DE genes were supposed to be associated with the wool growth cycling regulation. The majority of the genes possibly related to follicle development and wool growth could be assigned into the categories including regulation of receptor binding, extracellular region, protein binding and extracellular space. Several gene families of growth factors might take part in hair growth regulation, including fibroblast growth factors, transforming growth factor-β, insulin-like growth factor, and so on. Proteomic analysis also identified dozens of differentially expressed proteins. This whole genomic study may shed light on mechanism of the wool growth cycling control in Aohan fine wool sheep.
References
Janich P, Pascual G, Merlos-Suarez A, Batlle E, Ripperger J, Albrecht U, Obrietan K, Di Croce L, Benitah SA (2011) The circadian molecular clock creates epidermal stem cell heterogeneity. Nature 480:209–214
Rhee H, Polak L, Fuchs E (2006) Lhx2 maintains stem cell character in hair follicles. Science 312:1946–1949
Cotsarelis G (2006) Gene expression profiling gets to the root of human hair follicle stem cells. J Clin Invest 116:19–22
Ohyama M, Terunuma A, Tock CL, Radonovich MF, Pise-Masison CA, Hopping SB, Brady JN, Udey MC, Vogel JC (2006) Characterization and isolation of stem cell-enriched human hair follicle bulge cells. J Clin Invest 116:249–260
Wenguang Z, Jianghong W, Jinquan L, Yashizawa M (2007) A subset of skin-expressed microRNAs with possible roles in goat and sheep hair growth based on expression profiling of mammalian microRNAs. OMICS 11:385–396
Yu Z-D, Bawden CS, Henderson HV, Nixon AJ, Gordon SW, Pearson AJ (2006) Micro-arrays as a discovery tool for wool genomics. Proc NZ Soc Anim Prod 66:129–133
Yu Z-D, Gordon SW, Pearson AJ, Henderson HV, Craven AJ, Nixon AJ (2008) Gene expression profiling of wool follicle growth cycles by cDNA microarray. Proc NZ Soc Anim Prod 68:39–42
Norris BJ, Bower NI, Smith WJ, Cam GR, Reverter A (2005) Gene expression profiling of ovine skin and wool follicle development using a combined ovine–bovine skin cDNA microarray. Aust J Exp Agric 45:867–877
Penagaricano F, Zorrilla P, Naya H, Robello C, Urioste JI (2012) Gene expression analysis identifies new candidate genes associated with the development of black skin spots in Corriedale sheep. J Appl Genet 53:99–106
Smith WJ, Li Y, Ingham A, Collis E, McWilliam SM, Dixon TJ, Norris BJ, Mortimer SI, Moore RJ, Reverter A (2010) A genomics-informed, SNP association study reveals FBLN1 and FABP4 as contributing to resistance to fleece rot in Australian Merino sheep. BMC Vet Res 6:27
Bongiorni S, Chillemi G, Prosperini G, Bueno S, Valentini A, Pariset L (2009) A tool for sheep product quality: custom microarrays from public databases. Nutrients 1:235–250
Faucon F, Rebours E, Bevilacqua C, Helbling JC, Aubert J, Makhzami S, Dhorne-Pollet S, Robin S, Martin P (2009) Terminal differentiation of goat mammary tissue during pregnancy requires the expression of genes involved in immune functions. Physiol Genom 40:61–82
Ollier S, Robert-Granie C, Bernard L, Chilliard Y, Leroux C (2007) Mammary transcriptome analysis of food-deprived lactating goats highlights genes involved in milk secretion and programmed cell death. J Nutr 137:560–567
Keane OM, Zadissa A, Wilson T, Hyndman DL, Greer GJ, Baird DB, McCulloch AF, Crawford AM, McEwan JC (2006) Gene expression profiling of naive sheep genetically resistant and susceptible to gastrointestinal nematodes. BMC Genom 7:42
MacKinnon KM, Burton JL, Zajac AM, Notter DR (2009) Microarray analysis reveals difference in gene expression profiles of hair and wool sheep infected with Haemonchus contortus. Vet Immunol Immunopathol 130:210–220
Dong Y, Xie M, Jiang Y, Xiao NQ, Du XY, Zhang WG, Tosser-Klopp G, Wang JH, Yang S, Liang J, Chen WB et al (2013) Sequencing and automated whole-genome optical mapping of the genome of a domestic goat (Capra hircus). Nat Biotechnol 32(2):135–141
Wang HR, Feng ZC, Du M, Ren JK, Li HR (1994) Initial research for seasonal variation of wool growth of Aohan fine wool sheep. Inner Mong Anim Sci 3:1–3 (in Chinese)
Yu JJ, Liu JF, Zhao JS, Cheng M, Liu KD, Liu N (2012) Gene chip analysis of expression pattern of type I inner root sheath (IRS) keratin in Aohan wool sheep. Agric Sci Technol 13:1171–1174
Zhao JS, Li HG, Liu KD, Liu N, Li JQ (2012) Differential expression of immune genes between body side skin and groin skin of Aohan fine wool sheep. Agric Sci Technol 13:2475–2479
Stenn KS, Paus R (2001) Controls of hair follicle cycling. Physiol Rev 81:449–494
Dennis G Jr, Sherman BT, Hosack DA, Yang J, Gao W, Lane HC, Lempicki RA (2003) DAVID: database for annotation, visualization, and integrated discovery. Genome Biol 4:P3
Berndt P, Hobohm U, Langen H (1999) Reliable automatic protein identification from matrix-assisted laser desorption/ionization mass spectrometric peptide fingerprints. Electrophoresis 20:3521–3526
Jones PH, Watt FM (1993) Separation of human epidermal stem cells from transit amplifying cells on the basis of differences in integrin function and expression. Cell 73:713–724
Kloepper JE, Hendrix S, Bodo E, Tiede S, Humphries MJ, Philpott MP, Fassler R, Paus R (2008) Functional role of beta 1 integrin-mediated signalling in the human hair follicle. Exp Cell Res 314:498–508
Brakebusch C, Grose R, Quondamatteo F, Ramirez A, Jorcano JL, Pirro A, Svensson M, Herken R, Sasaki T, Timpl R et al (2000) Skin and hair follicle integrity is crucially dependent on beta 1 integrin expression on keratinocytes. EMBO J 19:3990–4003
Ryu K, Yokoyama M, Yamashita M, Hirano T (2012) Induction of excitatory and inhibitory presynaptic differentiation by GluD1. Biochem Biophys Res Commun 417:157–161
Lein ES, Hawrylycz MJ, Ao N, Ayres M, Bensinger A, Bernard A, Boe AF, Boguski MS, Brockway KS, Byrnes EJ et al (2007) Genome-wide atlas of gene expression in the adult mouse brain. Nature 445:168–176
Magdaleno S, Jensen P, Brumwell CL, Seal A, Lehman K, Asbury A, Cheung T, Cornelius T, Batten DM, Eden C et al (2006) BGEM: an in situ hybridization database of gene expression in the embryonic and adult mouse nervous system. PLoS Biol 4:e86
Churko JM, Chan J, Shao Q, Laird DW (2011) The G60S connexin43 mutant regulates hair growth and hair fiber morphology in a mouse model of human oculodentodigital dysplasia. J Invest Dermatol 131:2197–2204
Umeda-Ikawa A (2009) Time-course expression profiles of hair cycle-associated genes in male mini rats after depilation of telogen-phase hairs. Int J Mol Sci 10:1967–1977
Powell BC, Beltrame JS (1994) Characterization of a hair (wool) keratin intermediate filament gene domain. J Invest Dermatol 102:171–177
Yu Z, Gordon SW, Nixon AJ, Bawden CS, Rogers MA, Wildermoth JE, Maqbool NJ, Pearson AJ (2009) Expression patterns of keratin intermediate filament and keratin associated protein genes in wool follicles. Differentiation 77:307–316
Goh BK, Common JE, Gan WH, Kumarasinghe P (2009) A case of dermatopathia pigmentosa reticularis with wiry scalp hair and digital fibromatosis resulting from a recurrent KRT14 mutation. Clin Exp Dermatol 34:340–343
Lugassy J, Itin P, Ishida-Yamamoto A, Holland K, Huson S, Geiger D, Hennies HC, Indelman M, Bercovich D, Uitto J et al (2006) Naegeli–Franceschetti–Jadassohn syndrome and dermatopathia pigmentosa reticularis: two allelic ectodermal dysplasias caused by dominant mutations in KRT14. Am J Hum Genet 79:724–730
Horner ME, Parkinson KE, Kaye V, Lynch PJ (2011) Dowling-Degos disease involving the vulva and back: case report and review of the literature. Dermatol Online J 17:1
Lueking A, Huber O, Wirths C, Schulte K, Stieler KM, Blume-Peytavi U, Kowald A, Hensel-Wiegel K, Tauber R, Lehrach H et al (2005) Profiling of alopecia areata autoantigens based on protein microarray technology. Mol Cell Proteomics 4:1382–1390
Rosenquist TA, Martin GR (1996) Fibroblast growth factor signalling in the hair growth cycle: expression of the fibroblast growth factor receptor and ligand genes in the murine hair follicle. Dev Dyn 205:379–386
Acknowledgments
We are very grateful to Prof. Yinlin Ge for technical assistance. This research was supported by the Project of National Hair Sheep Industry Technology System (CARS-40) and the Project of Qingdao People’s Livelihood Science and Technology (13-1-3-88-nsh).
Author information
Authors and Affiliations
Corresponding author
Additional information
Nan Liu and Hegang Li contributed equally to this paper.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Liu, N., Li, H., Liu, K. et al. Differential expression of genes and proteins associated with wool follicle cycling. Mol Biol Rep 41, 5343–5349 (2014). https://doi.org/10.1007/s11033-014-3405-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-014-3405-1