
Abstract
The presenting study was investigated the associations between individual susceptibility and cigarette smoke on maternal chromosomal damage and neonatal birth growth in smoking mothers since little known about genetic susceptibility to cigarette smoke in relation to adverse pregnancy outcome such as birth growth. Sixty-one pregnant women who completed a questionnaire at Ankara Education and Research Hospital, Department of Obstetrics and Gynecology have enrolled in this study. GSTM1 and OGG1 ser326Cys gene polymorphisms were analysed by RFLP-PCR (Restriction Fragment Length Polymorphism-Polymerase Chain Reaction) as possible genetic factors affecting susceptibility to such health effects of smoking and chromosomal damage was performed by chromosomal aberration assay (CAA) in maternal blood lymphocytes. Maternal self-reported history of pregnancy smoking was informed by questionnaire declaration. Our results showed that maternal smoking had significant effect on chromosomal damage, birth weight, and length. The frequencies of CA in smokers was significantly higher than that of the nonsmokers (3.46 ± 2.06 and 2.00 ± 1.3, P = 0.001). Birth weight and length in smokers were significantly higher that of nonsmokers (3,355 g and 49.57 cm, P = 0.001; 3,639 g and 50.79 cm, P = 0.002). On the other hand, there was a slightly increased in the frequencies of CA and reduction birth weight and length in GSTM1 null and length in OGG1 variant genotypes, those differences were not statistically significant (P > 0.05); likely due to small sample size. Larger sample size needs to reach significance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Numerous studies have shown that maternal cigarette smoking during pregnancy is associated with reduced birth weight or increased risk of low birth weight [1–3]. However, not all women who smoke cigarettes during pregnancy have low birth weighted neonates. In a similar manner, studies covering large populations for CAs were assessed have given conflicting results, with one study finding that CA frequencies were not increased by smoking and another finding shows that smoking caused a 10–20% increase [4]. Gene–environmental interactions might be the reason for these variability, which could partly explain why some pregnant women who are smoking do not experience low birth weight or some people either smoke or expose to the genotoxic agents do not develop adverse health outcomes or cancer. Thus, while previously the researchers were focused on solely on genotoxicity studies, today they switched on the evaluation of gene–environmental interactions in combination with genotoxicity tests. In accordance with the Pub Med database, there is only one study about maternal smoking and focused on this combination. The metabolic gene polymorphisms (CYP1A1, GSTT1, GSTM1, and NAT2), as well as DNA damage that was assessed by comet assay, of mothers who exposed to environmental tobacco smoke were investigated [2].
Tobacco smoke contains approximately 4,000 compounds; the most important carcinogens are polycyclic aromatic hydrocarbons (PAHs), aryl amines, and N-nitrosamines. Cigarette smoking induces oxidative stress which is attributed to the action of reactive oxygen species (ROS) on different cellular components [5, 6]. Fetal toxicity may be mediated, at least in part, by the embryonic bioactivation of xenobiotics to free radical intermediates that can lead to oxidative stress and ultimately teratogenesis and in some cases carcinogenesis [7]. Placenta is one of the main sources for ROS [8]. The first step in xenobiotic-initiated toxicity is often the bioactivation of the xenobiotic to a more toxic form, which if possible, is detoxified by the body’s defense mechanisms, such as detoxifying enzymes; GSTs, CYP1A, NAT2 etc. The role of genetic polymorphisms is important in bioactivating and detoxifying enzymes and cancer risk [9, 10]. One of the detoxifying enzymes is glutathione-s- transferases (GSTs) which are involved in the detoxification phase and detoxify smoke-derived toxic chemicals. They allow conjugation between electrophilic metabolites and glutathione molecules [11, 12]. Both GST genes have either ‘null’ alleles or can be polymorphic [13].
Mammalian DNA is exposed to continuous attack by ROS that can cause significant oxidative damage. Smoking is known to be linked to increased oxidative DNA damage [14]. 8-Hydroxyguanosine (8-OHG) is an abundant DNA base lesion formed during oxidative stress which, if not repaired, can give rise to G: C → T:A transversions in DNA [15]. Protective systems maintain the vital integrity of DNA. Among these protective systems, base excision repair (BER) is specialized to remove the oxidatively damaged bases [16, 17]. 8-Oxoguanine DNA glycosylase-1 (OGG1) has significant importance in DNA integrity and OGG1-initiated BER pathway operates to remove 8-OHG lesions. OGG1 deletion and polymorphism may result in a hypermutator phenotype and susceptibility to oxidative pathologies including cancer. Limited and conflicting evidence exists regarding the repair capacity of a prevalent the OGG1 Cys326 variant [15]. Both GSTM1 and OGG1 genes are highly polymorphic in the population and have important roles in metabolizing and repairing of the compounds in cigarette smoke and repairing of DNA damage induced by cigarette smoke. Compounds in cigarette smoke are genotoxic and therefore, chromosome damage appears to be an excellent biomarker for determining the effect of exposure to smoking [18]. CAA is important for monitoring populations exposed to genotoxic agents because it allows the evaluation of the entire genome to identify mutagenic and carcinogenic chemicals [18, 19].
In general, it is an important topic to investigate effects of smoking and possible modifiers of susceptibility among mothers themselves as well as among neonates born to smoking mothers. Thus, the aim of our study was contribute to the controversial and very limited data on associations between smoking and gene polymorphisms on chromosomal damage, neonatal birth weight and length in smoking mothers. It showed, like numerous previous studies have done, a significant influence of maternal smoking during the pregnancy on mean birth weight/length of the neonates. In addition, our study observed an increased frequency of CAs in mothers who were smokers as compared to nonsmokers. Investigation of the role of polymorphisms in gene encoding enzymes of xenobiotic metabolism (GSTM1 in our study) and DNA repair (OGG1 in the present study) as potential to reveal important gene–environment interactions, especially in the context of tobacco smoke exposure. To date, there has been a few biological and epidemiological evidence of an association between “the SNP polymorphisms and smoking” or “DNA damage and smoking”, no study exists all parameters together like our study. To the best of our knowledge, our study constitutes the first study which investigates the interaction with maternal metabolic gene (GSTM1)—repair gene (OGG1) and smoking and their effect on birth weight and length and possible genotoxic effects of maternal smoking.
Materials and methods
Study subjects and sample collection
Healthy, pregnant women (n = 61) who participated in this study were from the largest Ankara Hospital, Gynecology and Obstetrics Clinics in Ankara, Turkey. They were divided into two groups (28 smokers and 33 nonsmokers) as non-smoking and smoking according to questionnaire declaration. Maternal peripheral blood samples were collected at the end of the delivery from all subjects February to September 2005 period. Five milliliters of peripheral blood was collected in a sterile EDTA container via venipuncture from each subject to determine gene polymorphisms by PCR–RFLP. After birth, 2 ml of peripheral blood samples were collected in heparinized vacutainer glass tube processed on the same day for genotoxicity analysis (CAA).
The groups (smokers and nonsmokers) were well matched for age (28.46 ± 4.58 years for smokers and 28.06 ± 5.18 years for nonsmokers) and weight (152.36 lb [69.11 ± 9.25 kg] for smokers and 181.2 lb [73.45 ± 12.32 kg] for nonsmokers) (P > 0.05, Table 1). Age, weight and Body Mass Index (BMI; kg/m2) (before and after birth), birth weight, neonate age (weeks), medical history and reproductive history were obtained from questionnaire declaration. The study was approved by Ankara Education and Research Hospital in Ankara, Turkey. Details are given in Table 1.
Lymphocyte cultures and chromosomal aberration scoring
Peripheral blood lymphocytes were taken from mothers at the end of delivery and processed immediately. The detailed method for analyzing of the CA can be found elsewhere [19]. A total of 100 well-spread metaphases were analyzed from each subject using ×100 (magnification) oil objective on a Zeiss Axioskop microscope. Only complete metaphase cells were selected for analysis. The following types of aberrations were examined during analysis: [1] chromosome break; [2] chromosome gap; [3] chromatid break; [4] chromatid gap; [5] acentric fragment (a pair of chromatids without a centromere); [6] minute, [7] dicentric and [8] ring.
DNA isolation and genotypic analysis
DNA was extracted from whole blood using a sodium perchlorate/chloroform extraction method.
GSTM1 and OGG1 Ser326Cys gene polymorphisms were analyzed by PCR–RFLP assay as described previously [13].
GSTM 1 genotyping
The PCR primers used were: Primer 1 (AA1): 5′-CGCCATCTTGTGCTA CATTGCCCG-3′; Primer 2 (AA2): 5′-ATCTTCTCCTCTTCTGTCTC-3′ and Primer 3 (AA3): 5′-TTCTGGATTGTAGCAGATCA-3′. The primers were obtained from Operon Technologies, Inc., Alameda, CA, USA.
A 550 μl PCR reaction was performed using 1 μl genomic DNA, 25 μM each primer (5.5 μl AA1, AA2, AA3), 16.5 μl 10 mM dNTPmix (dTTP, dATP, dGTP, dCTP), 2.75 Unit Taq DNA polymerase (Bioline) using 16.5 μl 50 mM MgCl2 and 55 μl 10× NH4 buffer supplied by the manufacturer and 444 μl 5% sterile water with DMSO. The reaction consisted of 35 cycles of 1 min at 93°C, 1.5 min at 52°C, 2 min at 70°C and 1 cycle of final extension for 10 min at 70°C. The specific size of GSTM1 gene PCR product (230 bp) was initially assessed at 100 V by electrophoresis with 1% Tris-Borate-EDTA electrophoresis buffer (TBE buffer) in 2% agarose gel.
OGG1 genotyping
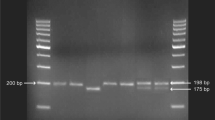
We used a simple PCR–RFLP method to identify the Ser326Cys variant, because the C to G transversion creates a Sat I restriction site. Briefly, the 207 bp fragment was amplified by PCR in a 30 μl reaction volume that contained 100 ng genomic DNA, 0.2 mM of dNTP (Fermentas Life Sciences, Lithuania), 1.5 mM of MgCl2 (Fermentas Life Sciences, Lithuania), 0.3 pmol of each primer and 1 unit of Taq DNA polymerase (Fermentas Life Sciences, Lithuania). The primers used for amplification of OGG1 gene exon 7, containing Ser326Cys, were 5′-ACT GTC ACT AGT CTC ACC AG-3′ forward (Iontek) and 5′-TGA ATT CGG AAG GTG CTT GGG GAA T-3′ reverse (Iontek). Cycling conditions were as follows: initial denaturation at 94°C for 2 min, then amplification by 33 cycles of denaturation at 94°C for 15 s, annealing at 60°C for 30 s, and elongation at 72°C for 35 s, followed by extension at 72°C for 10 min. The PCR product was digested with 3 units of Sat I (Fermentas, Lithuania) at 37°C for 16 h, and then electrophoresed on a 3% agarose gel. The Cys/Cys homozygote (G/G) is cleaved by Sat I, and yields 2 bands (100 and 107 bp bands). The Ser/Ser homozygote (C/C) is not cleaved by Sat I, and the single 207 bp band remains. The Ser/Cys heterozygote (C/G) contains all 3 bands (100, 107, and 207 bp bands) following restriction digestion. Negative controls (no template) and positive controls were included in all sets.
Statistical analysis
Statistical analysis was done at SPSS for Windows Release 11.5. Descriptive statistics were given as mean ± SD for continuous data, frequencies and percentages for categorical data. Chi-square test was used to analysis categorical data. For continuous data like age, weight, length, Student’s t test was used to compare two groups. To analyze difference between groups with more than two categories, one way ANOVA was used. When the difference between groups was significant at the result of ANOVA, Bonferroni test was used as post-hoc test. Backward elimination linear regression analysis with dummy variables was performed to find out which factors have effects on birth weight and length. A P value < 0.05 was considered statistically significant.
Results
Our results showed that smoking during pregnancy had significant effect on birth weight and length. Neonates of smoking mothers were lighter than those of non-smokers. The mean birth weight of smokers was 3,355 g and that of the nonsmokers 3,639 g, resulting in a significantly lower mean birth weight of 284 g for neonates of smoking mothers (P = 0.001, Table 1). The mean birth length of smokers was 49.57 cm and that of the nonsmokers 50.79, resulting in a significantly lower mean birth length of 1.22 cm for neonates of smoking mothers (P = 0.002, Table 1). Our results suggested that maternal smoking had a significant effect on birth weight and length without taking the impact of the genetic polymorphisms into account. We compared smokers to nonsmokers in terms of birth weight (≥3,000 and <3,000 g or ≥50 and <50 cm) and length to demonstrate the effect of smoking on gestational outcome. We found that the risk of low birth weight and birth length in smokers was statistically significant increased than nonsmokers (OR = 10.667, 95% CI = 1.222–93.084 and OR = 4.500 95% CI = 1.419–14.266; respectively) (Table 2). On other hand, there was a significant effect of neonate birth weight on mother’s weight (before and after delivery) and BMI (before and after delivery) (P < 0.05) however, there were no any effect of mother age, mothers’ weight gained during pregnancy, genotypes used and frequency of chromosomal aberration on birth weight and length according to logistic regression analysis (Table 3).
Smokers with the GSTM1 null and OGG1 C/G + G/G, variant genotypes had neonates with lower mean birth weight than non-smokers with the same genotype (3,394 ± 417.2 and 3,309 ± 408.5 for smokers, 3,603 ± 176.7 and 3,715 ± 294.8 for nonsmokers, P < 0.05, Table 4). Similarly, smokers with the GSTM1 null and OGG1 C/G + G/G genotypes had neonates with shorter mean birth length than non-smokers with the same genotype (49.89 ± 1.6 and 49.27 ± 1.8 for smoker, 50.47 ± 1.3 and 51.06 ± 1.4 for nonsmokers, P < 0.05, Table 4). When considering a combination of genotypes, no effect was found on both birth weight and length, likely due to small sample size (P > 0.05, Table 4). Sample size was reduced since only joint genotypes were evaluated.
The frequencies of CA in smoking mothers were statistically significant higher than those of nonsmokers (3.46 ± 2.06 and 2.00 ± 1.3, P = 0.001, Table 1).When the frequencies of CA in smoking mothers were compared to nonsmoking mothers for each variant (GSTM1 null and wild type; OGG1 C/C and C/G + GG), a significant increase was found in smokers (C/C; 2.06 ± 1.14 and C/G + G/G; 1.94 ± 1.50 for nonsmokers; C/C; 3.12 ± 1.76 and C/G + G/G; 4.00 ± 2.45 for smokers P < 0.05, Table 4). The frequency of CAs in smokers (3.70 ± 2.06) with the GSTM1 wild type was found higher than nonsmokers (1.61 ± 1.20) with the same genotype (P = 0.002 for smokers, Table 4). However, when the frequency of CAs in all mothers with the GSTM1 wild type was compared to those with the GSTM1 null, the significance was disappeared. Similar comparisons were made between wild and variant genotypes and a slightly higher frequency of OGG1 variant genotype was observed, but not significant (Table 4).
Our results showed that birth weight and length were not influenced by genetic polymorphisms without taking the impact of the maternal smoking into account as an environmental factor. When birth weight and length in GSTM1 wild type were compared to GSTM1 null genotypes and in OGG1 C/C and variant genotypes, there was no difference found. Similar comparisons were performed on birth weight and length regarding gene–gene interactions taking the joint genotypes into account, although the sample size was reduced, certainly no difference was observed.
We also did not find any difference in gestational age between smokers and nonsmokers (38.71 ± 1.09 and 38.53 ± 0.96, P = 0.5, Table 1). It might be considered maternal smoking had no impact on gestational age.
Discussion
Numerous studies have shown that maternal cigarette smoking during pregnancy is associated with increased risk of low birth growth [1, 20]. Furthermore, cigarette smoking during pregnancy leads to many complications and a neonate has twice the risk of low birth weight when compared to neonates who mothers do not smoke [21, 22]. However, not all women who smoke cigarettes during pregnancy have low birth weight neonates and some adverse pregnancy outcomes. Variations among women might be explained by genetic susceptibility. Recently, a gene–environmental interaction model has been suggested, which could partly explain why some pregnant women with smoking exposure do not have adverse effects [3]. Gene–environmental interactions relate to variations in the mother’s ability to metabolize smoking-related adducts to less harmful products and to repair the smoking induced oxidative base damage. Thus, we hypothesized that maternal smoking induced low birth weight/length and chromosomal damage might be modulated by genetic polymorphisms in metabolism and repair, GSTM1 and OGG1, respectively.
Compounds present in cigarette smoke undergo bioactivation and/or detoxication. Phase I of this process results in the formation of reactive epoxides, which can form DNA adducts initiating and promoting mutagenesis, carcinogenesis, or teratogenesis [23]. Polymorphic gene studies of cigarette smoke carcinogens have found variability between individuals [1, 12]. Multiple tobacco carcinogens require metabolic activation to interact with DNA and exert their genotoxic potential. GSTs are a supergene family of enzymes which is associated with the metabolism of some compounds in cigarette and involved in the detoxification of these compounds [24]. GSTM1 detoxifies specific biologically active metabolites of PAHs, and carriers of the GSTM1 null genotype have a reduced ability to detoxify them [25]. Smoking are known to be linked to increased oxidative DNA damage [14]. The formation of 8-OHG is induced by ROS from cigarette smoke, environmental stressors etc. and 8-OHdG, one of the major products of oxidative DNA modifications, was measured as a marker of oxidative stress [15] OGG1 protein has a critical role in repairing various types of DNA base modifications, especially major lesion 8-OHdG. It is well known that smoking induces 8-OHdG and that this lesion is repaired less in smokers with susceptible genotype [26]. Both GSTM1 and OGG1 genes are highly polymorphic in the population [13, 27]. We investigated the associations between individual susceptibility and cigarette smoke on maternal chromosomal damage and neonatal birth growth in smoking mothers since little known about genetic susceptibility to cigarette smoke in relation to adverse pregnancy outcome such as birth growth. Our present findings showed that neonatal birth weight and length are significantly lower for mothers who were continuous smokers during pregnancy compared with nonsmokers. Our results are consistent with previous studies [22, 28–30]. These findings are worthy since low birth weight might be associated with the increase in the incidence of a number of disease in later life, including coronary heart disease, stroke, hypertension, type 2 diabetes mellitus, insulin resistance, serum lipids, and premature pubarche [31].
To date, studies have focused on metabolic gene polymorphisms in smoking mothers due to metabolizing of compounds in cigarette smoke. We first time investigated the relative role of metabolic gene (GSTM1) versus other gene which has repair function (OGG1) in determining genetic susceptibility to chromosomal damage and birth weight/length. Wang et al. [1, 32] found that maternal cigarette smoking was associated with reduced birth weight and the adverse effects of maternal smoking on neonatal birth weight and gestational age were modified by maternal CYP1A1 and GSTT1 genotypes. Delpisheh et al. determined the role of maternal metabolic gene polymorphisms (CYP1A1, GSTT1 and GSTM1) in modulating the association between pregnancy smoking exposure and fetal growth restriction. They found that variant and null genotypes were more frequent in intrauterine growth restriction (IUGR) cases than non-IUGR controls and the mean birth weight of neonates of smokers (2,816 g) was 365.5 g lower than that of nonsmokers (3,181.5 g) [3]. Our results showed that there was a slight reduction on birth weight and length, but not statistically significant association was found between neonatal birth weight/length and maternal smoking as linked to the GSTM1 and OGG1 Ser326Cys genotype. Gloria-Bottini et al. investigated the possible influence of adenylate kinase genetic variability on the effect of maternal smoking on intrauterine selection and development. They observed that zygotes carrying AK1*2 allele were relatively protected from the damaging effects of smoking [33]. Sasaki et al. [25] conducted a prospective cohort study to estimate the effects of maternal smoking and genetic polymorphisms on infant birth weight and length and found that birth weight and length were significantly lower among infants born to continuously smoking women having the aryl hydrocarbon receptor (AhR) wild type genotype, the CYP1A1 variant genotype or the GSTM1 null genotype. All these studies demonstrate associations between genetic polymorphisms and birth weight/length or adverse pregnancy outcomes in maternal smoking. As we known that the smoking habit is considered to be an important means of exposure to genotoxins for humans [4]. Several lines of evidence suggest fetal susceptibility to carcinogens and that in utero exposure to cigarette smoke and environmental pollutants can result in carcinogenic DNA adducts, DNA damage, CAs and increased risk of childhood cancer [34–36]. There is one research article carried by Wu et al. They determined the effects of environmental tobacco smoke (ETS) on fetal growth and maternal DNA damage, and investigated the relationships among metabolic gene polymorphisms, genotoxicity, and pregnancy outcome in smoker mothers. Wu Our hypothesis is similar to Wu et al.’s. However, they only analyzed the metabolic gene polymorphisms and investigated the extent of DNA damage by comet assay. In our study, both metabolic and repair gene polymorphisms were examined. They did not find significant associations between metabolic genotypes and DNA damage [2]. Our results are consistent with theirs’. We also did not find significant association. The frequencies of CA in variant genotypes was slightly higher than those in wild type genotypes, however this increase was not significant. We considered that sample size was inadequate for genotypic analysis. Another research was carried out by Wu et al. on associations of passive smoking and genetic susceptibility with birth weight. They analysed only two metabolic gene polymorphisms, namely CYP1A1 MspI and EPHX1 Tyr13 His and found that there was a remarkable decrease in birth weight in the passive group. They concluded that their data was suggestive of possible gene–environmental interaction [2].
A number of methodological limitations should be considered when interpreting our results. First, cigarette smoke is a complex mixture of chemicals and other metabolic genes may be involved, also similarly other DNA repair genes. We only examined one metabolic and one repair genes, might not enough to show the role of genotypes on birth weight and length. Second, possible uncontrolled or inadequately controlled risk factors may cause confounding effects. We tried to standardize most of factors via questionnaire but still there might be some deficiencies are left. Third, we considered that sample size was sufficient in comparisons of groups for cytogenetic analysis, not adequate for genotyping analysis since sample size was extremely reduced in comparisons of gene–gene interaction.
It is uneasy to increase sample size, but multiple metabolic and repair genes (more than one associated with each other) in determining precise genetic susceptibility and genotoxicity tests may be analyzed and well-controlled the considerable factors belonging to neonates and their mothers.
Conclusion
In conclusion, to date, there has been very limited study, only one study as mentioned above on the associations between individual susceptibility and cigarette smoke on maternal chromosomal damage, neonate birth weight and length. Our data showed that smoking had significant effect on chromosomal damage, neonatal birth weight and length and they might be considered to be modulated by gene polymorphisms, if sample size was increase. It seems that still there is a need to extend studies due to its importance to gene–environmental interaction model and further studies are also needed to confirm and compare/discuss with previous study.
References
Wang X, Zuckerman B, Pearson C, Kaufman G, Chen C, Wang G, Niu T, Wise PH, Bauchner H, Xu X (2002) Maternal cigarette smoking, metabolic gene polymorphism, and infant birth weight. JAMA 287:195–202
Wu FY, Wu HD, Yang HL, Kuo HW, Ying JC, Lin CJ, Yang CC, Lin LY, Chiu TH, Lai JS (2007) Associations among genetic susceptibility, DNA damage, and pregnancy outcomes of expectant mothers exposed to environmental tobacco smoke. Sci Total Environ 386:124–133
Delpisheh A, Brabin L, Topping J, Reyad M, Tang AW, Brabin BJ (2009) A case-control study of CYP1A1, GSTT1 and GSTM1 gene polymorphisms, pregnancy smoking and fetal growth restriction. Eur J Obstet Gynecol Reprod Biol 143:38–42
DeMarini DM (2004) Genotoxicity of tobacco smoke and tobacco smoke condensate: a review. Mutat Res 567:447–474
Tabacova S, Baird DD, Balabaeva L (1998) Exposure to oxidized nitrogen: lipid peroxidation and neonatal health risk. Arch Environ Health 53:214–221
Dizdaroglu M, Jaruga P, Birincioglu M, Rodriguez H (2002) Free radical-induced damage to DNA: mechanisms and measurement. Free Radic Biol Med 32:1102–1115
Wan J, Winn LM (2006) In utero-initiated cancer: the role of reactive oxygen species. Birth Defects Res (Part C) 78:326–332
Qanungo S, Murherjea M (2000) Ontogenic profile of some antioxidants and lipid peroxidation in human placental and fetal tissues. Mol Cell Biochem 215:11–19
Anderson LM (2006) Environmental genotoxicants/carcinogens and childhood cancer: bridgeable gaps in scientific knowledge. Mutat Res 608:136–156
Chang JS (2009) Parental smoking and childhood leukemia. Methods Mol Biol 472:103–137
Stucker I, de Waziers I, Cenee S, Bignon J, Depierre A, Milleron B, Beaune P, Hemon D (1999) GSTM1, smoking and lung cancer: a case-control study. Int J Epidemiol 28:829–835
Hong YC, Lee KH, Son BK, Ha EH, Moon HS, Ha M (2003) Effects of the GSTM1 and GSTT1 polymorphisms on the relationship between maternal exposure to environmental tobacco smoke and neonatal birth weight. J Occup Environ Med 45:492–498
Kocabas NA, Karahalil B, Karakaya AE, Sardas S (2000) Influence of GSTM1 genotype on comet assay and chromosome aberrations after induction by bleomycin in cultured human lymphocytes. Mutat Res 469:199–205
Karahalil B, Emerce E, Koçer B, Han S, Alkiş N, Karakaya AE (2008) The association of OGG1 Ser326Cys polymorphism and urinary 8-OHdG levels with lung cancer susceptibility: a hospital-based case-control study in Turkey. Arh Hig Rada Toksikol 59:241–250
Smart DJ, Chipman JK, Hodges NJ (2006) Activity of OGG1 variants in the repair of pro-oxidant-induced 8-oxo-2′-deoxyguanosine. DNA Repair (Amst) 5:1337–1345
Stuart JA, Karahalil B, Hogue BA, Souza-Pinto NC, Bohr VA (2004) Mitochondrial and nuclear DNA base excision repair are affected differently by caloric restriction. FASEB J 18:595–597
Radaka Z, Gotob S, Nakamotob H, Ududc K, Papaic Z, Horvathc I (2005) Lung cancer in smoking patients inversely alters the activity of hOGG1 and hNTH1. Cancer Lett 219:191–195
Au WW, Cajas-Salazar N, Salama S (1998) Factors contributing to discrepancies in population monitoring studies. Mutat Res 400:467–478
Karahalil B, Yağar S, Bahadir G, Durak P, Sardaş S (2005) Diazepam and propofol used as anesthetics during open-heart surgery do not cause chromosomal aberrations in peripheral blood lymphocyte. Mutat Res 581:181–186
Windham GC, Hopkins B, Fenster L, Swan SH (2000) Prenatal active or passive tobacco smoke exposure and the risk of preterm delivery or low birth weight. Epidemiology 11:427–433
Ellard GA, Johnstone FD, Prescott RJ, Ji-Xian PW, Jian-Hua M (1996) Smoking during pregnancy: the dose dependence of birthweight deficits. Int J Obstet Gynecol 103:806–813
Ohmi H, Hirooka K, Mochizuki Y (2002) Fetal growth and the timing of exposure to maternal smoking. Pediatr Int 44:55–59
Hartsfield JK Jr, Hickman TA, Everett ET, Shaw GM, Lammer EJ, Finnell RA (2001) Analysis of the EPHX1 113 polymorphism and GSTM1 homozygous null polymorphism and oral clefting associated with maternal smoking. An J Med Genet 102:21–24
Masetti S, Botto N, Manfredi S, Colombo MG, Rizza A, Vassalle C, Clerico A, Biagini A, Andreassi MG (2003) Interactive effect of the glutathione S-transferase genes and cigarette smoking on occurrence and severity of coronary artery risk. J Mol Med 81:488–494
Sasaki S, Kondo T, Sata F, Saijo Y, Katoh S, Nakajima S, Ishizuka M, Fujita S, Kishi R (2006) Maternal smoking during pregnancy and genetic polymorphisms in the Ah receptor, CYP1A1 and GSTM1 affect infant birth size in Japanese subjects. Mol Hum Reprod 12:77–83
Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, Yamaguchi R, Kasai R, Itoh H (1997) Cigarette smoking induces an increase in oxidative DNA damage, 8-hydroxyguanosin, in a central site of the human lung. Carcinogenesis 18:1763–1766
Karahalil B, Kocabas NA, Ozcelik T (2006) DNA repair gene polymorphisms and bladder cancer susceptibility in a Turkish population. Anticancer Res 26(6C):4955–4958
Blake KV, Gurrin LC, Evans SF, Beilin LJ, Landau LI, Stanley FJ, Newnham JP (2000) Maternal cigarette smoking during pregnancy, low birth weight and subsequent blood pressure in early childhood. Early Hum Dev 57:137–147
Wills RA, Coory MD (2008) Effect of smoking among indigenous and non-indigenous mothers on preterm birth and full-term low birthweight. Med J Aust 189:490–494
Nijiati K, Satoh K, Otani K, Kimata Y, Ohtaki M (2008) Regression analysis of maternal smoking effect on birth weight. Hiroshima J Med Sci 57:61–67
Barker DJP (1998) In utero programming of chronic disease. Clin Sci 95:115–128
Tsai HJ, Liu X, Mestan K, Yu Y, Zhang S, Fang Y, Pearson C, Ortiz K, Zuckerman B, Bauchner H, Cerda S, StubbleWeld PG, Xu X, Wang X (2008) Maternal cigarette smoking, metabolic gene polymorphisms, and preterm delivery: new insights on G£E interactions and pathogenic pathways. Hum Genet 123:359–369
Gloria-Bottini F, Magrini A, Pietropolli A, Bergamaschi A, Bottini E (2009) Smoking and human reproduction: the effect of adenylate kinase genetic polymorphism. Am J Perinatol 26:117–121
Ramsey MJ, More DH II, Briner JF, Lee DA, Olsen L, Senft JR, Tucker JD (1995) The effects of age and lifestyle factors on the accumulation of cytogenetic damage as measured by chromosome painting. Mutat Res 338:95–106
Pluth JM, Ramsey MJ, Tucker JD (2000) Role of maternal exposures and newborn genotypes on newborn chromosome aberration frequencies. Mutat Res 465:101–111
Perera FP, Tang D, Tu YH, Cruz LA, Borjas M, Bernert T, Whyatt RM (2004) Biomarkers in maternal and newborn blood indicate heightened fetal susceptibility to procarcinogenic DNA damage. Environ Health Perspect 112:1133–1136
Acknowledgments
Experiments carried out in the present study comply with the current laws of the country in which the experiments were performed. The authors would like to give special thanks to all participants. We wish also thank to Assoc. Prof. Kenan Özköse for statistical analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Karahalil, B., Emerce, E., Kocabaş, N.A. et al. Associations between GSTM1 and OGG1 Ser326Cys polymorphisms and smoking on chromosomal damage and birth growth in mothers. Mol Biol Rep 38, 2911–2918 (2011). https://doi.org/10.1007/s11033-010-9953-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-010-9953-0