
Abstract
A novel tyrosine aminotransferase gene (designated as SmTAT) involved in rosmarinic acid biosynthesis pathway is cloned from Salvia miltiorrhiza Bung. The full-length cDNA of SmTAT is 1,603 bp long with an open reading frame (ORF) of 1,233 bp encoding a polypeptide of 411 amino acid residues. The deduced amino acid sequence of the SmTAT gene shared high homology with other known TATs. Analysis of SmTAT genomic DNA reveals that it contains 6 exons, 5 introns. The analysis of SmTAT promoter region and terminator region was also presented. Semi-quantitative RT-PCR analysis reveals that the constitutive expression of SmTAT in stem is much higher than that in root, leaf. Further expression analysis reveals that the signaling components of defense/stress pathways, such as methyl jasmonate (MeJA), abscisic acid (ABA), salicylic acid (SA) and ultraviolet-B radiation (UV-B), up-regulate the SmTAT transcript levels over the control. This study provides useful information for further studying this gene and its function in rosmarinic acid biosynthetic pathway in S. miltiorrhiza, the roots of which so-called ‘‘Danshen’’ possess many pharmaceutical properties for human health.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Salvia miltiorrhiza Bunge (Lamiaceae) is a well-known Chinese herb, and its roots, known as Danshen in Chinese, have been widely used for the treatment of menstrual disorders and cardiovascular diseases, and for the prevention of inflammation. S. miltiorrhiza roots contain two major classes of chemicals, a class of lipid-soluble diterpene quinone pigments, generally known as tanshinones, and a class of water-soluble phenolic acids including salvianolic acids, rosmarinic acid (RA), lithospermic acid, and 3,4-dihydroxyphenyllactic acid (DHPLA) or Danshensu [1]. Although tanshinones were originally thought to be the main active ingredients of Danshen, the phenolic acid constituents have been recently found to have significant bioactivities such as antioxidant and anti-ischemia reperfusion which may contribute to the therapeutic effects of Danshen [2].
Rosmarinic acid is an ester of caffeic acid and 3, 4-dihydroxyphenyllactic acid, which is mainly found in the plant species of Boraginaceae and Lamiaceae. RA has a number of interesting biological activities, such as antiviral, antibacterial, antiinflammatory, antioxidant and so on. In the plant kingdom, rosmarinic acid is supposed to act as a preformed constitutively accumulated defence compound. RA biosynthesis in these plants has been suggested to involve both the phenylpropanoid pathway (for the caffeic acid moiety) and a tyrosine-derived pathway (for the DHPLA moiety) as depicted by Fig. 1 [3].

The metabolic pathway leading to rosmarinic acid and 3, 4-dihydroxyphenyllactic acid (3, 4-DHPLA) biosynthesis found in Coleus blumei plant cell cultures [3]. Abbreviations: PAL, phenylalanine ammonia-lyase; C4H, cinnamic acid 4-hydroxylase; 4CL, hydroxycinnamate: coenzyme A ligase; TAT, tyrosine aminotransferase; 4-HPPA, 4-hydroxyphenylpyruvic acid; HPPR, hydroxyphenylpyruvate reductase; HPLA, 4-hydroxyphenyllactic acid; RAS, rosmarinic acid synthase; 3′-H and 3-H, hydroxycinnamoyl-hydroxyphenyllactate 3- and 3′-hydroxylases
Tyrosine aminotransferase (TAT), which catalyses the transamination from l-tyrosine to 4-hydroxyphenylpyruvate, is the first enzyme in the tyrosine-derived branch of the biosynthetic pathway of RA. Since 4-hydroxyphenylpyruvate is not an intermediate of aromatic amino acid biosynthesis in plants, it has to be formed from tyrosine as described above. In addition to rosmarinic acid biosynthesis it serves as precursor for homogentisic acid, a precursor for the essential metabolites plastoquinones and tocopherols, the latter of which are known to be radical scavengers in plants. The enzyme was completely purified some years ago [4], and cDNA clones were reported for a few plants, such as Coleus blumei (GenBank Accession number: AJ458993), Arabidopsis thaliana [5], Medicago truncatula (GenBank Accession number: DQ006809), and Glycine max (AAY21813). At present, the specific biosynthetic pathways for the production of RA and related phenolic acids in S. miltiorrhiza are still not clear. Furthermore, the lack of detailed knowledge of pathway regulation has led to the speculations that every enzymatic step in the rosmarinic acid biosynthesis pathway has either a “key” or regulatory role. The cloning and characterization of the genes involved in RA biosynthetic pathway would greatly help to reinforce the knowledge in higher plants.
In this study, we hope to use S. miltiorrhiza to extend our knowledge on rosmarinic acid pathway. In our study, we report the cloning and characterization of a novel tyrosine aminotransferase (SmTAT) from the S. miltiorrhiza in rosmarinic acid biosynthesis pathway. Comparison of these reported TAT gene sequences from different species reveals that the TAT genes are structurally conserved, and might possess similar functions. The work also aims to examine the expression profiles of SmTAT in different tissues and under the stress conditions including light or signal molecules. The abscisic acid (ABA), methyl jasmonate (MeJA) and salicylic acid (SA) were used as signal molecules, and ultraviolet-B radiation (UV-B) was used as light. Their effects on the accumulation of SmTAT mRNA were determined.
Materials and methods
Plant materials
Seeds of S. miltiorrhiza were pretreated with 75% alcohol for 1 min, washed 3 times with distilled water, followed by the treatment of 0.1% HgCl2 for 5 min and by four rinses with sterile distilled water. The sterilized seeds were then incubated between several layers of sterilized wet filter paper under 30°C for germination. Germinated seeds were sowed and cultured in small plastic flowerpots naturally. The seedlings were all grown at 25°C under 14 h light/10 h dark photoperiod cycles for 2 months until various treatments, RNA and DNA isolation.
Various treatments
The leaves from 2-month-old S. miltiorrhiza seedlings grown in small plastic flowerpots were sprayed with solution of 50 μM abscisic acid (ABA), 500 μM methyl jasmonate (MeJA) and 500 μM salicylic acid (SA) respectively followed by RNA isolation. Another set of control plants were similarly treated with distilled water. For ultraviolet-B radiation (UV-B) treatment, the seedlings were exposed under 1500 μJ/m2 intensity of illumination for 30 min.
RNA and DNA isolation
Total RNAs of various treatments from 2-month-old S. miltiorrhiza plant were extracted using TRIzol Reagent (GIBCO BRL) according to the manufacturer’s instruction [6]. The genomic DNA of S. miltiorrhiza was isolated using a Cetyl trimethyl ammonium bromide (CTAB)-based method [7]. The quality and concentration of RNA and DNA samples were examined by EB-stained agarose gel electrophoresis and spectrophotometer analysis.
Molecular cloning of the SmTAT full-length cDNA
Molecular cloning of SmTAT from S. miltiorrhiza was carried out by Rapid amplification of cDNA ends (RACE) method using a SMART™ RACE cDNA Amplification Kit (Clontech, USA).
For 3′RACE of SmTAT, about 100 ng of total RNA was reverse transcribed with 3′-CDS primer by BD PowerScript Reverse Transcriptase (Clontech, USA). Universal Primer A Mix (UPM), Nested Universal Primer A (NUP) (Clontech, USA), gene-specific primers SM-TAT-3′-1 [5′-AACCCTGGGAACCC(A/T)TGTGG(A/G)AA(C/T)GTG-3′, as 3′ RACE first amplification primer] and SM-TAT-3′-2 [5′-AGATGGTTGGTTCCTGGCTGGCGCC-3′, as 3′ RACE nest amplification primer] were used. The PCR was conducted in accordance with the protocol provided by the manufacture (Clontech, USA). The nested amplified PCR product was purified and cloned into PMD18-T vector (TaKaRa, Japan) and then sequenced.
For 5′ RACE of SmTAT, about 100 ng of total RNA was reverse transcribed with 5′-CDS primer and SMART™ II A Oligonucleotide (Clontech, USA). Universal Primer A Mix (UPM), Nested Universal Primer A (NUP) (Clontech, USA), gene-specific primers SM-TAT-5′-anti1 (5′-AAGCCATTGCTCCCTTAGGTTTTGTTG-3′, as 5′ RACE first amplification primer) and SM-TAT-5′-anti2 (5′-TGTCACAGTATTTCTTGATGCGTTCC-3′, as 5′ RACE nest amplification primer) were used. The PCR was conducted in accordance with the protocol provided by the manufacture (Clontech, USA). The nested amplified PCR product was purified and cloned into PMD18-T vector (TaKaRa, Japan) and then sequenced. By aligning and assembling the products of 3′ and 5′RACE, the full-length SmTAT from S. miltiorrhiza was deduced and subsequently amplified by proof-reading PCR amplification with primers SM-TAT-C-F (5′-CTCGATTCAGTTGCGGCAATGGAGTTG-3′) and SM-TAT-C-R (5′-CGACTTAGTAGGAGTGCCGTTCACAGA-3′). The PCR procedure was conducted under the following conditions: 5 min at 94°C, 5 cycles (30 s at 94°C, 30 s at 70°C, 2.5 min at 72°C), 30 cycles (30 s at 94°C, 30 s at 65°C, 2.5 min at 72°C) and 7 min at 72°C. The amplified PCR product was purified and cloned into PMD18-T vector (TaKaRa, Japan) and then sequenced.
Isolation of genomic sequence of SmTAT
In order to detect whether there exist introns within the SmTAT, PCR amplification was carried out using the same reaction system as that for the cloning of the full-length cDNA except that the template was substituted by 1.5 μg of total genomic DNA and the extension time at 72°C in the amplification cycles was prolonged up to 3 min.
Genome Walker DNA libraries were constructed using the Universal Genome Walker Kit (Clontech, USA). The genomic DNA was completely digested with different blunt-end restriction enzymes (EcoRV, Pvu II, Stu I and DraI) (Takara, Japan) and DNA fragments were ligated separately to the Genome Walker adaptor using the DNA Blunting Kit (Takara, Japan).
The amplification of upstream sequence of the known sequence consists of two PCR amplifications. The primary PCR uses the outer adaptor primer AP1 (5′-GTAATACGACTCACTATAGGGC-3′) provided in the kit and an outer, gene specific primer 5GSP1 (5′-AGCAGCATTACTAGCGTGGAAGCAC-3′). The amplification was performed in a GeneAmp PCR System 2400 for 7 cycles with 25 s at 94°C, 3 min at 72°C and then for 32 cycles with 25 s at 94°C, 3 min at 67°C. After the final cycle, the amplification was extended for 4 min at 67°C. The primary PCR mixture was then diluted and used as a template for nested PCR with the nested adaptor primer AP2 (5′-ACTATAGGGCACGCGTGGT-3′) provided in the kit and a nested gene-specific primer 5GSP2 (5′-ACTCCGAATCTCAGCTGCTTCAATGAG-3′). The amplification was performed in a GeneAmp PCR System 2400 for 5 cycles with 25 s at 94°C, 3 min at 72°C and then for 20 cycles with 2 s at 94°C, 3 min at 67°C. After the final cycle, the amplification was extended for 4 min at 67°C. The PCR products were analyzed by electrophoresis, purified from agarose gel, and then cloned into pMD18 vector followed by sequencing. Sequencing result showed that a clone contained a fragment of about 1,200 bp. The amplification of downstream sequence of the known sequence consists of two PCR amplifications. The primary PCR uses the outer adaptor primer AP1 and an outer, gene-specific primer 3GSP1 (5′-ATGGTAGCCGTTGAAGGAGTTGGATAG-3′). The nested PCR uses the nested adaptor primer AP2 and a nested gene-specific primer 3GSP2 (5′-ATGTATTGCTCTCCTCGAGTAAATATTG-3′). The conditions of the PCR reaction were the same as mentioned above. The PCR product was cloned into the pMD18-T-vector (TaKaRa) and sequenced with DYEnamic Direct dGTP Sequencing Kit (Amersham) and a 373A DNA sequencer.
Sequence analyses of SmTAT
Sequence alignments, ORF translation and molecular mass calculation of the predicted protein were carried out on Vector NTI Suite 8. Genbank BLASTs were carried out on NCBI (http://www.ncbi.nlm.nih.gov/), while bioinformatic analysis of the predicted SmTAT protein was carried out on website of Expasy Molecular Biology Server (http://www.expasy.org). Phylogeneticanalysis of SmTAT and other known TATs from other plant species retrieved from GenBank (Table 1) were aligned with CLUSTAL W (1.82) and subsequently a phylogenetic tree was constructed by the neighborjoining (NJ) method using the software of MEGA 2.0 [8–10]. The reliability of the tree was measured by bootstrap analysis with 1,000 replicates [11]. Homology-based structural modeling was performed by Swiss-Model [12], and Web Lab View Lite 4.0 was used to display 3-D structures. Promoter motifs were analyzed using the PlantCARE database (a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences, http://www.intra.psb.ugent.be:8080/PlantCARE).
Expression profile of SmTAT in different tissues and under various stresses
Semi-quantitative RT-PCR was used to investigate the expression profiling of SmTAT in different tissues of the plant and under various stresses. Total RNA was extracted from root, stem and leaf tissues of S. miltiorrhiza, and from leaves of the plant subjected to various treatments, followed by incubation with Rnase-free Dnase I at 37°C for 30 min according to the manufacturer’s instruction (Takara, Japan). A mock treatment was carried out with H2O as a control to exclude any other factors induced SmTAT gene expression that might result from the spraying. After establishing agreement between the OD values of the RNAs from different tissues, aliquot of 1 μg total RNA (1 μg/μl) was used as the template in one-step RT-PCR analysis using One-step RT-PCR Kit (TaKaRa, Japan) with SM-TAT-C-F (5′-CTCGATTCAGTTGCGGCAATGGAGTTG-3′) and SM-TAT-C-R (5′-CGACTTAGTAGGAGTGCCGTTCACAGA-3′) as primers. The template was reversely transcribed at 50°C for 30 min and denatured at 94°C for 5 min, followed by 26 cycles of amplification (94°C for 35 s, 50°C for 35 s, 72°C for 2.5 min). The RT-PCR reaction for the house-keeping gene (18 S gene) using specific primers 18SF (5′-ATGATAACTCGACGGATCGC-3′) and 18SR (5′-CTTGGATGTGGTAGCCGTTT-3′) was performed as described above as the control. The densities of the target bands were measured with a WEALTEC Dolphin-DOC ultraviolet analyzer (WEALTEC, USA). Each sample was assayed in triplicate.
Results
Cloning of the full-length cDNA of SmTAT
Using the RACE method and primers mentioned above, cDNA ends of 706 bp and 1003 bp were amplified by 3′ RACE and 5′RACE respectively. 3′ and 5′ ends were assembled with Vector NTI Suite 8.0 and the full-length SmTAT cDNA (Genbank accession no. DQ334606) was subsequently amplified by proof-reading PCR amplification with primers mentioned above. The full-length cDNA of SmTAT was 1,603 bp with 159 bp 5′ and 182 bp 3′ untranslated regions, 29 bp poly A tail and contained a 1,233 bp ORF encoding a 411-amino-acid protein (Fig. 2). BLAST search revealed that the nucleotide sequence of SmTAT had sequence similarities to TAT genes from other plant species.

The full-length cDNA sequence and the deduced amino acid sequence of SmTAT gene. The start codon (ATG) is in italics and the stop codon (TAA) is in bold. The putative poly (A) signal (AATAA) is boxed, and aminotransferases family-I pyridoxal-phosphate attachment site (SLSKrwLVpGWRLG) is shaded
Generation and characterization of the full-length DNA of SmTAT
Gene-specific primers derived from the start and stop codon regions of the SmTAT cDNA were designed and synthesized to isolate the genomic SmTAT gene by PCR. A 2,492 bp PCR product was amplified, which had 100% identity in the coding region with the full-length SmTAT cDNA sequence. It was found that 5 introns were present in the genomic SmTAT sequence by comparing the genomic and cDNA sequences, and exon 1 (255 bp) exon 2 (339 bp) exon 3 (219 bp), exon 4(220 bp), exon 5 (91 bp), exon 6 (109 bp) were separated by the intron 1 (462 bp), intron 2 (92 bp), intron 3 (400 bp), intron 4 (105 bp) and intron 5 (91 bp) respectively (Fig. 4). It was also found that the putative splicing site obeyed the GU/AG rule.
Sequence alignment of the overlapping DNA sequences yielded approximately 4.8 kb of genomic DNA. The complete sequence has been submitted to the NCBI database (GenBank Accession No: EF192320). Analysis of 5′ flanking region of SmTAT gene by comparing the genomic and cDNA sequences identified a transcription start site (A) 119 bp upstream from the start codon ATG. Usually the structure of 5′ flanking region of eukaryotic genes includes four parts: site of transcription start, TATA box, CAAT box and GC box. The consensus sequence for TATA box is [T(CG)TATA(TA)A1–3(CT)A] that has been shown to be important for eukaryotic transcription [13]. Using the PlantCARE database, 54 TATA sequences were found upstream from the start of transcription in SmTAT gene and the most probable TATA box was located at −28 bp of the predicted transcription initiation site.
About 30 CAAT sequences were found upstream from the start of transcription. These sequences may correspond to the CAAT box that is sometimes important for the efficiency of eukaryotic transcription [14].
In total six types of cis-acting elements were predicted with relation to disease resistance in SmTAT promoter: (1) 2 sites of ABRE, cis-acting element involved in the abscisic acid responsiveness, TACGTG or CACGTG; (2) 1 site of TCA-element which is a cis-acting element involved in salicylic acid responsiveness, with conform sequence of AGAAAA or AGAAGA; (3) 1 site of ethylene-responsive element ERE, containing core sequence of ATTTCTAA or ATTTGAAA; (4) 1 site of fungal elicitor responsive element Box-W1, ATTAAT; (5) 2 sites of TC-rich repeats, cis-acting element involved in defense and stress responsiveness, ATTTTCTCCA or ATTTTCTTCA; (6) CGTCA-motif and TGACG-motif, cis-acting regulatory element involved in the MeJA-responsiveness.
Other types of cis-acting elements were also predicted within SmTAT promoter region, including eight types of light responsive elements such as ACE, Box 4, Box I, CATT-motif, G-Box, G-box, MRE and Pc-CMA2c, 1 cis-acting regulatory element involved in circadian control named circadian and 1 Skn−1_ motif, the cis-acting regulatory element required for endosperm expression (Figs. 3 and 4).

Nucleotide sequence of SmTAT gene promoter region, the predicted cis-acting regulatory elements, enhancers and repressors related to basic promoter activities. Different elements are painted with different background. The most probable TATA box is with purple background. The CAAT boxes in the upsteam are boxed. Abscisic acid responsive elements are painted with yellow background, a salicylic acid responsive element is in the blue background, an ethylene-responsive element is in the gray background, a fungal elicitor responsive element is in the green background, two cis-acting elements involved in defense and stress responsiveness are in the brown background, the cis-acting regulatory elements involved in the MeJA-responsiveness are painted with bright green background, three cis-acting regulatory element required for endosperm expression are underlined, while a cis-acting regulatory element involved in circadian control is painted with violaceous background. The transcription initiation site A is shown in the red background

The start codon (ATG) is in bold and the stop codon (TAA) is in bold italics. Exons are sequences in the black background, and shadowed sequences denote introns
The 3′-untranslated region is 1,191 bases long and follows the coding region of SmTAT. It is also rich in A+T and 41 predicted stem structures (over the threshold −4.0 Kkal mol−1) reside in the transcribed mRNA sequence corresponding to this region. The stems are tandem organized and the whole mRNA sequence of this region forms a complicated secondary structure which resembles the clover-shaped tRNA structure with a free energy of −208.3 kkal mol−1. The pairwised stems and the unpairwised loops within this clover-shaped structure constitute many stem-loops or “panhandles”, and many of these stem-loops are followed by poly (A) sequences. Hence it is clear that this terminator region contains the basic structure information for transcriptional termination.
Characterization of the deduced SmTAT protein
By using the software of Computer pI/Mw Tool at http://www.expasy.org, the calculated isoelectric point (pI) and molecular weight of the deduced SmTAT protein were predicted to be 5.8 and 45.1 kDa, respectively.
Protein–protein BLAST showed that on the amino acid level SmTAT protein had high homology to TATs from other plant species. Through Clustal W/X, full-length alignment result showed that SmTAT protein shared 92.9% identity (the highest identity) to SsTAT from Solenostemon scutellarioides. In addition, SmTAT was 66.1, 66.1, and 67.2% identical to AtTAT, GmTAT and MtTAT respectively (Fig. 5).

Sequence alignment of the deduced SmTAT protein with other plant TATs. Motif 1 is the aminotransferases family-I pyridoxal-phosphate attachment site, and motif 2 is highly conserved residue Arg. Amino acid residues identical at a given position are denoted with dark backgrounds. Gaps are inserted in the sequences for better alignment
Many sites essential for TAT activities conserved in different plant species were also found in SmTAT. As shown in Fig. 5, the predicted SmTAT protein included a aminotransferases family-I pyridoxal-phosphate attachment site (SLSKrwLVpGWRLG) and the highly conserved residue in family I for the binding of these enzymes’ substrates, Arg385.
Three-dimensional structure analysis of SmTAT protein
In order to better characterize the SmTAT protein, a comparative modeling of three dimensional structure of SmTAT performed using SWISS-MODEL [15]. The theoretical monomer and homomer structures for SmTAT were portrayed against the template of TAT of Trypanosoma cruzi by Blankenfeldt et al. [16] using crystallography (Fig. 6). Alignments of three dimensional structure and structural analyses were performed using three dimensional molecular Viewer and Weblab viewerlite. As could be seen from (Fig. 6a, b), SmTAT strongly resembled the other TAT proteins such as SsTAT. This result indicated that the SmTAT shared the same biological activity with SsTAT, implying that SmTAT might have similar functions with SsTAT, a protein whose function had been verified previously. The TAT proteins functioned as a homodimer, and the predicted three dimensional structure of SmTAT revealed that two monomers formed a symmetric dimmer (Fig. 6c). It contains the highly conserved residue in family I for the binding of these enzymes’ substrates, Arg385 in S. miltiorrhiza TAT, which fixes the a-carboxylate of the incoming amino acid or a-ketoacid, which was shown in the homodimer.

The three-dimensional models of SsTAT (6b for monomer) and the deduced SmTAT (6a for monomer and 6c for homodimer). The models are predicted with SWISS-MODEL and showed with WebLab viewer. (a) and (b) α-helices were indicated by helices in red and β-sheets were indicated by patches in blue. Turns and loops were indicated by gray lines. (c) Ribbon representation of SmTAT homodimer. Monomer A was yellow-colored and monomer B was blue-colored. Two Arg sites were marked with balls
Molecular evolution analysis
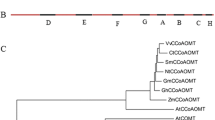
To investigate the evolutionary relationships among SmTAT and other TAT proteins, a phylogenetic tree was constructed based on the deduced amino acid sequences of predicted SmTAT and other TAT proteins from other species (Table 1). The analysis result highlighted three important features in all inferred species. Firstly, the phylogenetic tree analysis of the 13 TAT proteins was consistent with the evolutionary relationship between these species (Fig. 7), from prokaryote to eukaryote, from bacteria, botany and amphibian to mammal, from lower organisms to advanced organisms, suggesting TAT proteins have been conserved during evolution. Secondly, TAT sequence formed several distinct species-specific clusters. For example, all plant species formed a cluster, no matter whether they belonged to the gymnosperm or angiosperm, monocot or dicot plants. It was obvious that all animal species were grouped into a cluster. Meanwhile, TAT from E. coli, the prokaryote species, was separated from other eukaryote species in the tree (Fig. 7). Thirdly, SmTAT had the closest relationship with SsTAT from Solenostemon scutellarioides, lamiaceae species. Our result suggested that SmTAT shared a common evolutionary origin based on their similar roles and conserved structural and sequence characteristics such as amino acid homologies and conserved motifs.

Neighbor-joining phylogenetic tree of the complete sequences of SmTAT and other TAT proteins. Sequences were identified by the names of species. The numbers at each node represented the bootstrap values
Semi-quantitative RT-PCR Analysis of SmTAT
Total RNAs extracted from root, stem and leaf tissues were used to investigate spatial expression profiles of SmTAT by semi-quantitative RT-PCR analysis. The result showed that SmTAT constitutively expressed in all the tested tissues (Fig. 8a), with stronger expression in stems (Fig. 8a).

Expression profiles of SmTAT in different tissues and various stresses. Expression of SmTAT (a) in different tissues; (b) was induced under 50 μM ABA, 500 μM MeJA, 500 μM SA and UV-B (30 min) treatment. The S. miltiorrhiza 18S gene (126 bp) was used in RT-PCR amplification as the control
Because SA, MeJA, ABA and light signal motifs were found in the promoter region of SmTAT gene, in this study, to examine whether SmTAT was induced by these stimuli, S. miltiorrhiza leaves were treated with various inducers and harvested for RNA isolation at different time points. Under UV-B treatment, SmTAT transcript accumulation gradually increased in 24 h. SmTAT expression in S. miltiorrhiza was induced with a maximum at 8 h after SA treatment. Transcript levels of SmTAT increased as well under MeJA treatment, but then remained constant for at least 4–48 h, and then increased thereafter. ABA also up-regulated SmTAT expression, reached the highest expression level after 24 h and then declined (Fig. 8b).
Discussion
In our search for a novel TAT gene in S. miltiorrhiza, SmTAT was isolated with the predicted protein showing high similarity to members of TAT family. Analysis of SmTAT genomic DNA indicates that it contains 6 exons, 5 introns. By contrast, TAT gene revealed in Arabidopsis thaliana has 7 exons and 6 introns. The number of exons and introns in TAT genes appears to vary among species. Sequence analyses, in particular the three dimensional structure, implied that SmTAT might have the same catalytic function as other TAT proteins.
Since the major roles of plant secondary metabolites are to protect plants from attack by insect, herbivores and pathogens, or to survive other biotic and abiotic stresses, some strategies for culture production of the metabolites based on this principle have been developed to improve the yield of such plant secondary metabolites. These include treatment with various elicitors, signal compounds, and abiotic stresses [17–22]. Many such treatments indeed effectively promote the production of a wide range of plant secondary metabolites, both in vivo and in vitro.
TAT catalyses the reaction from tyrosine to p-hydroxyphenylpyruvate. It is the first enzyme in the tyrosine-derived pathway. Meantime, it is also the first enzyme in the biosynthetic pathway leading via homogentisic acid to plastoquinone and tocopherols, which are known to function as radical scavengers in plants and thus protect the plant in a variety of different stress situations. Increases in radicals and reactive oxygen species have been described after wounding of plants or exposure to high light and ultraviolet (UV) light [23–24].
In this study, to examine whether SmTAT was induced by other stimuli, S. miltiorrhiza leaves were treated with various inducers and harvested for RNA isolation at different time points.
Salicylic acid is globally involved in defense response during plant–pathogen interactions [25]. Exogenous application of SA, which is indigenously produced and functions in the development of SAR, results in the induction of resistance against pathogens and the expression of a set of SAR genes [26, 27]. SA induces the expression of SmTAT gene, indicating that SA is involved in the SmTAT -mediated pathway.
Methyl jasmonate and its free-acid jasmonic acid (JA), collectively referred to as jasmonates, are important cellular regulators involved in diverse developmental processes, such as seed germination, root growth, fertility, fruit ripening and senescence [28, 29]. In addition, jasmonates activate the defense mechanisms of plants in response to insect-driven wounding, various pathogens, and environmental stresses, such as drought, low temperature, and salinity. Recently, jasmonates have also been implied in the signaling pathway mediating induced defense responses in pathogen- or insect-attacked plants. Exogenously applied JA and its methyl ester, MeJA, are capable of inducing defense proteins and secondary defense metabolites in a wide range of plant species [29, 30]. Sandorf and Hollander-Czytko reported that JA played an important role in the induction of TAT and the biosynthesis of tocopherols upon environmental stress [31]. As expected, MeJA could also induce SmTAT expression in S. miltiorrhiza in this study.
Abscisic acid is a well-known senescence-triggering plant hormone. ABA plays important roles throughout plant life and development, affecting seed germination, plant growth, senescence and plant stress responses. Especially under some stresses like hyper- or hypo-osmotic stress, salt, cold and drought stress, ABA acts as an important signal molecule to regulate expression of sets of defense genes. ABA is also reported to regulate biosynthesis of secondary metabolites in some plant cell cultures. The results in the present study suggest that SmTAT gene expression was stimulated by ABA, indicating that SmTAT may be related to ABA signal transduction pathway.
The practical results above also are consistent with the previous prediction, because signal elements including SA, MeJA and ABA are found in the promoter region of SmTAT gene (Fig. 3).
UV light, as an abiotic stimulus, can promote secondary metabolites accumulation, and phenylpropanoids are the major plant secondary metabolite class that absorbs UV-B irradiation and provides a means of protection against UV-B damage and subsequent cell death by protecting DNA from dimerization and breakage [32, 33]. As expected, UV-B treatment on S. miltiorrhiza seedlings strongly increased SmTAT expression.
Rosmarinic acid played very important roles as health-protecting natural products in S. miltiorrhiza. TAT is a key enzyme in rosmarinic acid biosynthetic pathway. Although much research about TAT has been focused on the tyrosine, there has been an explosion of interests in rosmarinic acid health-promoting components of the human diet. With the increasing knowledge of RA biosynthetic pathway, it will be possible to have a complete understanding of the whole pathway, not only in the model plants but also in some valuable species, for the pathway in different plant may be different from each other. S. miltiorrhiza is one of the most economic valuable Chinese herbs. For the unique pharmacological properties of S. miltiorrhiza, the knowledge on the secondary metabolic pathway, including the gene cloning and characterization, will be a great motivation for bioengineering the secondary metabolics in S. miltiorrhiza.
References
Tang W, Eisenbrand G (1992) Chinese drugs of plant origin: chemistry, pharmacology, and use in traditional and modern medicine. Springer-Verlag, Berlin, pp 891–902
Li LN (1997) Water soluble active components of Salvia miltiorrhiza and related plants. J Chin Pharm Sci 6:57–64
Petersen M, Hausler E, Karwatzki B, Meinhard J (1993) Proposed biosynthetic-pathway for rosmarinic acid in cell-cultures of Coleus-blumei Benth. Planta 189:10–14
De Eknamkul W, Ellis BE (1987) Tyrosine aminotransferase: the entrypoint enzyme of the tyrosine-derived pathway in rosmarinic acid biosynthesis. Phytochemistry 26:1941–1946
Nakamura Y, Sato S, Kaneko T, Kotani H, Asamizu E, Miyajima N, Tabata S (1997) Structural analysis of Arabidopsis thaliana chromosome 5. III. Sequence features of the regions of 1,191,918 bp covered by seventeen physically assigned P1 clones. DNA Res 4:401–414
Jaakola L, Pirttila AM, Halonen M, Hohtola A (2001) Isolation of high quality RNA from bilberry (Vaccinium myrtillus L.) fruit. Mol Biotechnol 19:201–203
Rechards EJ (1995) Preparation and analysis of DNA. In: Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (eds) Short protocol in molecular biology. John Wiley and Sons, New York, pp 36–38
Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Thompson JD, Higgins DG, Gibson TJ (1994) Clustal-W—improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22:4673–4680
Kumar S, Tamura K, Jakobsen IB, Nei M (2001) MEGA2: molecular evolutionary genetics analysis software. Bioinformatics 17:1244–1245
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783–791
Guex N, Peitsch MC (1997) SWISS-MODEL and the Swiss-PdbViewer: an environment for comparative protein modeling. Electrophoresis 18:2714–2723
Joshi CP (1987) An inspection of the domain between putative TATA box and translation start site in 79 plant genes. Nucl Acids Res 15:6643–6653
Kozak M (1981) An analysis of 5′-noncoding sequence from 699 vertebrate messenger RNAs. Nucl Acids Res 9:5233–5252
Schwede T, Kopp J, Guex N, Peitsch MC (2003) SWISS-MODEL: an automated protein homology-modeling server. Nucleic Acids Res 31:3381–3385
Blankenfeldt W, Nowicki C, Montemartini-Kalisz M, Kalisz HM, Hecht HJ (1999) Crystal structure of Trypanosoma cruzi tyrosine aminotransferase: substrate specificity is influenced by cofactor binding mode. Protein Sci 8:2406–2417
Yukimune Y, Tabata H, Higashi Y, Hara Y (1996) Methyl jasmonate-induced overproduction of paclitaxel and baccatin III in Taxus cell suspension cultures. Nat Biotechnol 14:1129–1132
Zhao J, Fujita K, Yamada J, Sakai K (2001) Improved beta-thujaplicin production in Cupressus lusitanica suspension cultures by fungal elicitor and methyl jasmonate. Appl Microbiol Biotechnol 55:301–305
Zhao J, Hu Q, Guo YQ, Zhu WH (2001) Elicitor-induced indole alkaloid biosynthesis in Catharanthus roseus cell cultures is related to Ca2+ influx and the oxidative burst. Plant Sci 161:423–431
Zhao J, Zhu WH, Hu Q (2001) Enhanced catharanthine production in Catharanthus roseus cell cultures by combined elicitor treatment in shake flasks and bioreactors. Enzyme Microb Technol 28:673–681
Zhao J, Zhu WH, Hu Q, He XW (2000) Improved indole alkaloid production in Catharanthus roseus suspension cell cultures by various chemicals. Biotechnol Lett 22:1221–1226
Zhang CH, Yan Q, Cheuk WK, Wu JY (2004) Enhancement of tanshinone production in Salvia miltiorrhiza hairy root culture by Ag+ elicitation and nutrient feeding. Planta Med 70:147–151
Fryer MJ (1992) The antioxidant effects of thylakoid vitamin-e (alpha-tocopherol). Plant Cell Environ 15:381–392
Munne-Bosch S, Alegre L (2002) The function of tocopherols and tocotrienols in plants. Crit Rev Plant Sci 21:31–57
Ryals JA, Neuenschwander UH, Willits MG, Molina A, Steiner HY, Hunt MD (1996) Systemic acquired resistance. Plant Cell 8:1809–1819
Gaffney T, Friedrich L, Vernooij B, Negrotto D, Nye G, Uknes S, Ward E, Kessmann H, Ryals J (1993) Requirement of salicylic-acid for the induction of systemic acquired-resistance. Science 261:754–756
Delaney TP, Uknes S, Vernooij B, Friedrich L, Weymann K, Negrotto D, Gaffney T, Gutrella M, Kessmann H, Ward E, Ryals J (1994) A central role of salicylic-acid in plant-disease resistance. Science 266:1247–1250
Creelman RA, Mulpuri R (2002) The oxylipin pathway in Arabidopsis. In: Somerville CR, Meyerowitz EM (eds), The Arabidopsis book. American Society of Plant Biologists, Rockville, MD, pp 1–24
Wasternack C, Hause B (2002) Jasmonates and octadecanoids: signals in plant stress responses and development. Prog Nucl Acid Res Mol Biol 72:165–221
Wasternack C, Parthier B (1997) Jasmonate signalled plant gene expression. Trends Plant Sci 2:302–307
Sandorf I, Hollander-Czytko H (2002) Jasmonate is involved in the induction of tyrosine aminotransferase and tocopherol biosynthesis in Arabidopsis thaliana. Planta 216:173–179
Li JY, Oulee TM, Raba R, Amundson RG, Last RL (1993) Arabidopsis flavonoid mutants are hypersensitive to UV-b irradiation. Plant Cell 5:171–179
Lois R (1994) Accumulation of UV-absorbing flavonoids induced by UV-b radiation in Arabidopsis-thaliana l.1. mechanisms of UV-resistance in Arabidopsis. Planta 194:498–503
Acknowledgment
This research was financially supported by the National Natural Science Foundation of China (20572130).
Author information
Authors and Affiliations
Corresponding author
Additional information
Database Accession No: DQ355979
Rights and permissions
About this article
Cite this article
Huang, B., Yi, B., Duan, Y. et al. Characterization and expression profiling of tyrosine aminotransferase gene from Salvia miltiorrhiza (Dan-shen) in rosmarinic acid biosynthesis pathway. Mol Biol Rep 35, 601–612 (2008). https://doi.org/10.1007/s11033-007-9130-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-007-9130-2