
Abstract
Tolerance to waterlogging is an important breeding objective for barley (Hordeum vulgare L.); however, it is a complex quantitative trait. It is difficult to screen large numbers of lines in the field due to environmental variability, and it is also challenging to screen large numbers in controlled conditions if yield data are to be collected. The direct measurement of traits that contribute to waterlogging tolerance, such as aerenchyma development in roots, may offer advantages especially if molecular markers can be developed to screen breeding populations. A doubled haploid population from a cross between Franklin and YuYaoXiangTian Erleng was screened for adventitious root porosity (gas-filled volume per unit root volume) as an indicator of aerenchyma formation. A single QTL for root porosity was identified on chromosome 4H which explained 35.7 and 39.0 % of phenotypic variation in aerated and oxygen-deficient conditions, respectively. The nearest marker was EBmac0701. This QTL is located in the same chromosomal region that contributed to tolerance when the same population was screened in an earlier independent soil waterlogging experiment. Comparative mapping revealed that this QTL is syntenic with the Qaer1.02-3 QTL in maize and the Sub1A-1 gene in rice, which are associated with aerenchyma formation (maize) and submergence tolerance (rice), respectively. This is the first report of a QTL for root porosity in barley which elucidates a major mechanism of waterlogging tolerance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Waterlogging remains a significant constraint to cereal production across the globe in areas with high rainfall and/or poor drainage. It is estimated that constraints to soil drainage adversely affect 10 % of the global land area (FAO 2002 after Setter and Waters 2003) and climate change may also increase the severity and frequency of flooding events (Arnell and Liu 2001). In waterlogged soils, O2 is rapidly depleted, whilst gases that are produced, like CO2 and ethylene, rapidly accumulate. Reduced compounds such as Mn2+, Fe2+, S2− and carboxylic acids can also increase to toxic levels in waterlogged soils (Ponnamperuma 1984). Additionally, secondary metabolites including phenolics, volatiles, ethanol and acetaldehyde can have phytotoxic effects as a result of waterlogging (Shabala 2011). Plants in waterlogged soils face energy, carbohydrate and nutrient deficiencies (Colmer and Voesenek 2009).
Yield losses due to waterlogging can be significant in cereals. In barley (Hordeum vulgare L.), waterlogging is estimated to reduce yields on average by 20–25 %, but losses can exceed up to 50 % depending on the stage of plant development when the waterlogging occurs (Setter et al. 1999). The most economical way to reduce damage from waterlogging is to introduce tolerance into current varieties. However, breeding for waterlogging tolerance is difficult as it is a complex quantitative trait. In a study on the combining ability of waterlogging tolerance in barley, Zhou et al. (2007) found that although high heritability values could be achieved, they were easily reduced by varying experimental conditions or if heritability was determined from individual plant scores (as opposed to population averages). It is difficult to screen large numbers of lines in the field due to environmental variability (spatial and temporal) and also challenging to screen large numbers in controlled conditions, especially if the experiment attempts to grow plants to maturity to obtain yield data.
The direct measurement of traits that contribute to waterlogging tolerance, such as the development of aerenchyma in roots, may offer advantages. Aerenchyma is tissue containing a high proportion of gas-filled spaces which increase porosity (% gas volume per unit tissue volume) and provides a low-resistance pathway for long-distance gas transport (Armstrong 1979). Lysigenous aerenchyma, as occurs in barley, is promoted by the accumulation of ethylene and a biochemical cascade that leads to programmed cell death of cortical cells (Drew et al. 2000). Root porosity values in control conditions tend to be relatively low in non-wetland species such as wheat and barley (3–7 %) but can be induced to levels of up to 16–22 % under O2 deficit (Colmer 2003). The volume of gas-filled intercellular spaces (i.e. porosity) in roots without aerenchyma (e.g. young root tissues when grown in aerated conditions and with sufficient nutrients) is determined by the pattern of cell arrangements and can differ markedly for species (Justin and Armstrong 1987).
Although barley is considered sensitive to waterlogging compared with other cereals, significant variation for waterlogging tolerance amongst genotypes has been observed in barley. Several large-scale screening studies have identified lines with good tolerance (Takeda and Fukuyama 1986; Qiu and Ke 1991), and variation in tolerance levels has also been observed in other studies (Pang et al. 2004; Zhou et al. 2007). Pang et al. (2004) evaluated six barley genotypes from Australia, China and Japan for waterlogging tolerance and measured a variety of parameters including root and shoot biomass, root anatomy, chlorophyll content, stomatal conductance and rates of photosynthesis and transpiration. The Chinese variety, TX9425, displayed good tolerance and was considered a suitable waterlogging-tolerant parent. In contrast the Japanese variety, Nasu Nijo, was considered the most sensitive to waterlogging.
Positive correlations between aerenchyma development and plant growth under waterlogged conditions have been observed in wheat (Huang et al. 1994; Setter et al. 1999). Huang et al. (1994) grew six wheat varieties in aerated nutrient solution culture for 14 days then imposed treatments of aeration (21 kPa O2) or hypoxia (5 kPa O2). After 21 days hypoxia, the percentage of aerenchyma was determined microscopically in the cortical region of adventitious roots. The percentage of aerenchyma positively correlated with shoot growth (r = 0.938, P < 0.01). Setter et al. (1999) observed a positive correlation between the percentage of aerenchyma in adventitious roots and yield in 17 spring wheat varieties grown under intermittent waterlogging in the field near Esperance, Western Australia (r = 0.768, P < 0.001).
For barley, there are contrasting data. Eight barley varieties were included in the study of Setter et al. (1999), but no correlation was found between aerenchyma formation and yield following waterlogging. In other studies, however, some association between aerenchyma and waterlogging tolerance in barley has been observed. Pang et al. (2004) determined the percentage of aerenchyma in root cross-sections using image analysis software in TX9425 and Nasu Nijo, after 3 weeks waterlogging, and observed significantly more aerenchyma in the waterlogging-tolerant variety TX9425 (23.9 %) compared with the more sensitive Nasu Nijo (7.1 %).
Further analysis of root porosity, as an indicator of aerenchyma formation, and the identification of genes or QTL controlling traits associated with waterlogging tolerance in barley could be useful for marker-assisted selection (MAS) in breeding populations. A number of QTL have been identified for waterlogging tolerance in barley using traits such leaf chlorosis, plant biomass reduction and plant survival/plant health following soil waterlogging (Li et al. 2008; Zhou 2011; Zhou et al. 2012b). To date, however, no QTL for aerenchyma formation have been identified in barley and the relationship between root porosity and waterlogging tolerance has not been confirmed. In maize, a number of QTL for aerenchyma formation have been identified following crosses between maize and the wild relative teosinte, which readily forms aerenchyma (Mano et al. 2007, 2008). The aim of this study was to measure adventitious root porosity, as an indicator of aerenchyma formation, and plant growth in the same doubled haploid (DH) population as described by Zhou et al. (2012b) under aerated and O2-deficient (stagnant) conditions in nutrient solution to identify QTL controlling aerenchyma formation in barley.
Materials and methods
Screening barley varieties for variation in waterlogging tolerance and adventitious root porosity
Seven barley (Hordeum vulgare L.) varieties from Australia (Franklin and Yerong), Japan (Nasu Nijo) and China (TX9425, Yan89110, YSM1 and YYXT) were screened for waterlogging tolerance and adventitious root porosity to determine the best parents for this study. Waterlogging tolerance was assessed in Tasmania using the methodology and facilities outlined in Zhou (2011) which involved soil-based screening. Plants were grown in tanks containing soil and subjected to waterlogging at the three-leaf stage. Continuous waterlogging was imposed for 9 weeks, and then, plants were rated from 0 (susceptible) to 10 (tolerant) based on leaf chlorosis and plant survival. Root porosity was assessed in Western Australia using plants grown in a hydroponic system. All plants were grown in aerated nutrient solution for the first 14 days, then transferred to either a deoxygenated stagnant (0.1 % w/v agar) nutrient solution or maintained in an aerated nutrient solution for a further 21 days. Root porosity, an indicator of aerenchyma formation (Armstrong 1979), was measured on adventitious roots after 21 days of or aerated or stagnant treatment. Fresh and dry weights were taken at the beginning and end of the treatment period. The hydroponic set-up, nutrient solutions, root zone treatments and root porosity measurements were as described below.
Plant materials and solution culture
Two barley varieties which varied significantly in waterlogging tolerance and adventitious root porosity under stagnant conditions were selected following the preliminary screening experiments outlined above. A Chinese barley variety, YuYaoXiangTian Erleng (YYXT), was selected as the waterlogging-tolerant parent and an Australian barley variety, Franklin, was selected as the susceptible parent. A doubled haploid (DH) population was developed from Franklin/YYXT with 172 DH lines. A subset of 126 DH lines was randomly selected for this experiment.
Seeds were surface sterilised with 0.04 % (w/v) sodium hypochlorite for 45 s and rinsed thoroughly with deionised water. Seeds were then imbibed in aerated 0.5 mol m−3 CaSO4 for 3 h before being placed on plastic mesh floating on 0.1-strength aerated nutrient solution in a 20/15 °C (12 h day/night) controlled environment room. The solution was covered with aluminium foil to keep seeds in the dark. After 4 days, the solution was replaced with 0.25-strength solution, and the aluminium foil was removed. After 7 days, seedlings were transferred to aerated pots containing full-strength nutrient solution. Four seedlings were placed into each 4.5-L pot. Plants were held in the lids of the pots using polystyrene foam, and the pots and lids were covered in aluminium foil to ensure the roots were in darkness. The composition of the nutrient solution at full strength was (mol m−3): K+ 4.0, Ca2+ 4.0, Mg2+ 0.40, NH4 + 0.625, NO3 − 4.375, SO4 2− 4.4, H2PO4 − 0.20, Na+ 0.20, H4SiO4 − 0.10; and the micronutrients (mmol m−3): Cl 50, B 25, Mn 2, Zn 2, Ni 1, Cu 0.5, Mo 0.5 and Fe–Na EDTA 50. Additional iron was also supplied (5 mmol m−3) on Days 9 and 12 as iron sulphate, so as to avoid any possible mild iron deficiency that could otherwise occur in the seedlings. The solution also contained 2.5 mol m−3 2-[N-morpholino]ethanesulfonic acid (MES), and the pH was adjusted to 6.5 with KOH. All solutions were renewed weekly. On Day 14, an additional 2.5 mol m−3 NH4NO3 was added to all solutions, and this higher level of mineral nitrogen was maintained for the duration of the experiment. All chemicals were analytical grade.
Experimental design and root zone O2 treatments
Due to the large number of lines tested, the 126 DHs were screened in three batches with 42 DH lines screened in each batch. The duration of each batch was 35 days. Within each batch, lines were sown over three consecutive days as the time required to harvest and take measurements precluded sowing all lines on the same day. Additionally, because seed was pre-germinated for the experiment, it was more practical to include all replicates of a DH line on 1 day. Thus, in the first batch, DH lines 1–14 were sown on Day 1, DH lines 15–28 were sown on Day 2 and DH lines 29–42 were sown on Day 3, and so on for the remaining batches (Supplementary Table 1). The two parent lines were also included each day; thus, a total of 16 lines were sown on each day. When seedlings were transferred to pots of full-strength nutrient solution, 48 pots were prepared each day and two lines (two plants per line) were placed in each pot. Pots were assigned one of two treatments; ‘aerated’ or ‘stagnant’ and each line/treatment combination was replicated three times. The design was a completely randomised block design with two oxygenation treatments, three batches (and three germination days within each batch).
Each set of 48 pots fitted onto one bench in a 4 × 12 configuration. The pots sown on Days 1 and 2 fitted into one controlled environment room, whilst the pots sown on Day 3 were placed into a second controlled environment room (both at 20/15 °C day/night, 12 h cycles). The inclusion of the parent lines on each day (i.e. into each replicate within each batch) accounted for any day/room effects. Lines were randomised on each bench using CycDesigN 4.1 (2009). The design ensured that the same two lines did not occur in the same pot more than once on each bench and the randomisation of pots and plants was different for each batch.
The root zone treatments were imposed immediately after the initial harvest on Day 14 (refer below). In pots designated for aerated treatment, the nutrient solution continued to be bubbled with air for the duration of the experiment. Pots designated for the stagnant treatment were given a ‘hypoxic’ pre-treatment where pots were flushed with N2 for 3–4 h to decrease O2 in the solution to about one-tenth of that at air equilibrium. The pots were left unstirred overnight, and the following day (Day 15), the nutrient solution was replaced with de-oxygenated nutrient solution (flushed overnight with N2 gas) containing 0.1 % (w/v) agar. The inclusion of agar prevented convective movements in solution and simulated the changes in gas composition associated with soil waterlogging, i.e. impeded entry of O2 and impeded exit of ethylene and CO2 (Wiengweera et al. 1997). Solutions in the aerated pots were also replaced at this time with aerated nutrient solution (without agar). Pots were then returned to their allocated positions on the benches. All solutions were renewed every 7 days. If solution levels dropped, the pots were topped up with deionised water (aerated treatment) or deionised water containing 0.1 % w/v agar which had been de-oxygenated (stagnant treatment). After 21 days in the treatments (Day 35), plants were removed for final harvest measurements.
Plant growth measurements
Plant growth measurements were taken at two stages of the experiment. Initial harvest measurements were taken after 14 days growth, before treatments had been imposed and whilst all plants were maintained in aerated conditions. Final harvest measurements were taken on Day 35, at the conclusion of the 21-day treatment phase, where plants had been exposed to either an aerated or stagnant treatment.
At the initial harvest, one of the two plants in each replicate/treatment combination was removed and tiller number, shoot fresh/dry weight and root fresh/dry weight were recorded. Final harvest measurements, taken on the remaining plant, included the same parameters plus an additional measurement of longest adventitious root length. Root porosity measurements were also made on the remaining plant at the final harvest (described in next section).
Root porosity measurements
Root porosity (% gas-filled volume per unit tissue volume) was measured on samples of adventitious roots from each plant at the conclusion of the aerated or stagnant treatment phase by determining root buoyancy before and after vacuum infiltration of gas spaces in the roots with water (Raskin 1983) using equations modified by Thomson et al. (1990). To prepare the roots, the root mass was cut immediately below the crown with a scalpel, and approximately 15 adventitious roots, longer than 10 cm in length, were selected. Lateral roots were removed and the roots were lined up and cut at 5 cm (from the base which was previously attached to the crown) to provide a root sample of approximately 0.5 g (fresh weight) for porosity measurements. The balance was placed on a stand so that the pan hook beneath the balance could be accessed. A beaker containing approximately 900 ml deionised water was placed beneath the balance. Thus samples could be weighed normally on top of the balance (in air) or held in a clamp, attached to the pan hook and weighed under water (submerged). The water level in the beaker was maintained at the same level during all measurements. The root samples were rinsed in three washes of deionised water to remove any loose lateral roots, gently blotted to remove surface water, and the fresh weight of the root sample (in air) was recorded. The roots were then weighed (submerged) before and after vacuum infiltration. Firstly, an empty stainless steel aquarium clamp was weighed submerged in the beaker of water beneath the balance. Secondly, the root sample was placed in the clamp, so roots were held together but not compressed, and the roots and clamp were weighed in the same submerged fashion. Thirdly, the roots and clamp were transferred to a small beaker of water (enough to cover the roots) and subject to vacuum infiltration (3 × 5 min) and, finally, the roots and clamp were weighed again (submerged). These data were used to determine root porosity values for each sample using the equations of Thomson et al. (1990). The process of weighing, vacuum infiltration and re-weighing samples was carried out in sets of 12, and a total of 96 samples were processed each day. This meant that all samples from one bench (48 pots × 2 genotypes) were processed in a single day, and the root porosity and final harvest measurements were carried out over three successive days for each ‘batch’ (see details under heading ‘Experimental design and root zone O2 treatments’).
Statistical analysis
The experimental design was a completely randomised block with two oxygenation treatments, three batches and three germination days within each of the batches. The design was generated using CycDesigN 4.1 (2009). For each plant growth measurement, a linear mixed model was applied to investigate the main effects and interaction of DH lines and oxygenation conditions. This mixed model accounted for the design and randomisation structure. The fixed effects were tested using an approximate F test, and the random effects were tested using a Chi-squared test based on the residual maximum likelihood ratio test. All data were analysed using ASReml-R (Gilmour et al. 2005) with R statistical programme (R Core Team 2012).
Genetic map construction
A genetic map with a length of 1,147 cM, consisting of 782 DArT (Diversity Assays Technology) and 76 SSR markers (Zhou et al. 2012a), was constructed using Joinmap 4.0 (Van Ooijen 2006). DArT assays were conducted by Triticarte Pty. Ltd. and performed as described by Wenzl et al. (2006). These SSR markers were selected from a published genetic map (Varshney et al. 2007).
QTL mapping
The software MapQTL 5.0 (Van Ooijen 2004) was used to scan the QTL for the root porosity trait in barley. The permutation test was performed with 10,000 iterations. The minimum LOD value for the genome-wide threshold was 2.2 and 3.0 with P values 0.05 and 0.01, respectively. Genotype and phenotype data of each individual line, and their genetic maps, were imported to the software. Interval mapping was applied to detect possible QTL across the whole genome. Genetic markers with LOD threshold values greater than 3.0 in each region were selected as cofactors, and multiple QTL model (MQM) analysis was performed. MQM analyses were performed until the cofactors remained constant. The percentage of variance explained by each QTL, and additive effect was obtained using MQM mapping.
Results
Screening barley varieties for variation in waterlogging tolerance and adventitious root porosity
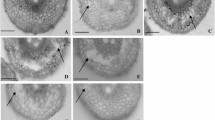
The seven varieties tested in the preliminary screening experiment differed in both waterlogging tolerance and adventitious root porosity. Waterlogging tolerance, screened in Tasmania, was rated from 0 (susceptible) to 10 (tolerant) based on leaf chlorosis and plant survival after 9 weeks continuous soil waterlogging (Fig. 1a, b). The varieties Franklin and Nasu Nijo, with ratings of 2.3 and 1.7, respectively, were significantly more susceptible to waterlogging than Yerong, TX9425, Yan89110, YSM1 and YYXT whose ratings ranged from 6.7 to 8.3. Root porosity values in Franklin and Nasu Nijo in stagnant conditions were also significantly lower than in the other varieties, 12.5 and 16.4 %, respectively (Fig. 2). The Chinese variety, YSM1, had the highest root porosity in stagnant conditions (22.8 %) followed by YYXT, Yerong, Yan89110 and TX9425 with root porosity values ranging from 19.5 to 20.4 %. Increased root porosity was significantly correlated with increased relative (i.e. % of control) root weights in both fresh (r = 0.886, P < 0.01) and dry (r = 0.909, P < 0.01) weights. There was also a significant positive correlation between soil waterlogging tolerance measured in Tasmania and root porosity measured in the stagnant deoxygenated agar nutrient solutions in Western Australia (r = 0.789, P < 0.05).

a Waterlogging tolerance scores of seven barley varieties following 9 weeks of continuous soil waterlogging (0 = susceptible, 10 = tolerant). b Barley lines with different waterlogging tolerance scores: A 10; B 9; C 3; D 8 (left) and 1 (right)

Root porosity values (% gas volume per unit root volume) of seven barley varieties following aerated or stagnant deoxygenated agar treatment in nutrient solution for 21 days
Plant growth in Franklin/YYXT population
Plant growth and development was evaluated in all lines on Day 14 before the treatments commenced (initial harvest). Pairwise comparisons between the groups of plants destined for aerated and stagnant treatments indicated that plant growth was similar, and there were no significant differences between the treatment groups for root and shoot fresh and dry weights (P values = 0.970 to 0.985). The average fresh weight for roots and shoots at initial harvest was approximately 1.05–1.10 g, whilst mean dry weights were 0.05 g (roots) and 0.10–0.11 g (shoots) (Table 1).
Plant growth and development was also evaluated on Day 35 at the conclusion of the 21-day treatment period (final harvest). Plant growth was reduced in all lines in the stagnant treatment compared with the aerated control (Table 1). The reductions were significant for all traits measured; tiller no. (P < 0.001); longest adventitious root length (P < 0.001); shoot fresh weight (P < 0.001); shoot dry weight (P < 0.001); root fresh weight (P < 0.005); and root dry weight (P < 0.05). Mean tiller number was reduced from 24 in the control to 14 in the stagnant treatment, and longest adventitious root lengths were also reduced, from 59.3 to 20.0 cm in Franklin and from 51.2 to 19.2 cm in YYXT. Longest root lengths in the DH population ranged from 26.8 to 78.4, with a mean of 55.0 cm in the aerated treatment, whilst in the stagnant treatment, longest root lengths ranged from 11.1 to 28.7 with a mean of 19.2 cm. These differences were reflected in root weights which were reduced by more than 50 % in plants exposed to stagnant conditions. In the aerated treatment, the mean fresh root weight over all lines was 28.9 g, whilst the in the stagnant treatment, the mean fresh root weight was 12.5 g. Dry root weights were reduced from 1.1 to 0.5 g. Shoot weights were similarly reduced with mean fresh weight dropping from 60.8 to 18.4 g and dry weights dropping from 5.2 to 1.9 g.
Adventitious root porosity in Franklin/YYXT population
Adventitious root porosity values were significantly higher following the stagnant treatment compared with the aerated treatment (P < 0.001). The mean root porosity value of all lines in the aerated treatment was 6.0 % compared with 11.4 % in the stagnant treatment (Table 1); see ‘Introduction’ for explanation of porosity in roots from aerated and low O2 conditions. There was considerable variation for root porosity within the Franklin/YYXT DH population (Fig. 3). In the aerated treatment, root porosity values ranged from 0.3 to 17.0 %, whilst in the stagnant treatment, values ranged from 5.0 to 21.4 %. The parental lines, YYXT and Franklin, did not differ significantly in the aerated treatment; mean root porosity was 7.0 % for YYXT and 5.8 % for Franklin (P = 0.106). In the stagnant treatment, however, root porosity values were significantly higher for YYXT, 12.6 %, compared with Franklin, 10.2 % (P < 0.001).

Frequency histogram of mean root porosity values (% gas volume per unit root volume) in a barley DH population of Franklin/YYXT following aerated or stagnant deoxygenated agar treatment in nutrient solution for 21 days
Increasing porosity values were significantly correlated with increases in absolute root fresh (r = 0.243, P < 0.01) and dry (r = 0.217, P < 0.02) weights and absolute shoot fresh (r = 0.183, P < 0.05) and dry (r = 0.223, P < 0.02) weights in the stagnant treatment. Increasing porosity values were also significantly correlated with increases in relative (i.e. % of control) root fresh (r = 0.252, P < 0.005) and dry (r = 0.232, P < 0.01) weights in stagnant conditions. Correlations were not significant between root porosity and relative shoot fresh (r = 0.084, P < 0.5) and dry (r = 0.142, P < 0.2) weights.
QTL identification in Franklin/YYXT population
A significant QTL was detected for adventitious root porosity in the barley genome and was mapped in the same position on 4H in both aerated and stagnant conditions. The nearest marker was EBmac0701. The QTL, with LOD values of 12.1 and 13.5, explained 35.7 and 39.0 % of phenotypic variation in aerated and stagnant conditions, respectively (Table 2). The parent YYXT contributed the allele for increased root porosity.
Significant QTL were also identified for other traits including root and shoot fresh and dry weights, tiller number and the length of the longest adventitious root. The positions of the QTL associated with root porosity and plant growth traits are indicated on a genetic map (Fig. 4). The various QTL accounted for between 9.0 and 16.7 % of phenotypic variation and were identified on 1H, 2H, 3H, 4H, 5H and 7H (Table 2). A number of QTL were identified on 1H for shoot/root weight and longest root length. A QTL for shoot weight was identified at 1.47–3.3 cM with closest markers bPb-1781 and bPb-5064. The QTL explained 11.6 and 16.0 % of variation for shoot fresh weight in aerated and anoxic conditions, respectively. The same QTL also explained 12.5 % of variation in shoot dry weight. A second QTL for shoot weight was located at 40.5 cM with closest marker GBM1042. This QTL accounted for 8.7 % of the variation in shoot fresh weight in aerated conditions and 11.5 % of variation in shoot dry weight in stagnant conditions. A QTL for root weight was located at 58 cM with closest marker EBmac0501. It explained 10.3 % of the variation in root fresh weight in stagnant conditions and 11.0 % of variation in root dry weight in both aerated and stagnant conditions. Two QTL for longest root length were identified at 17 (aerated conditions) and 35 cM (stagnant conditions) which explained 9.0 and 11.2 % of the variation in longest root length, respectively.

Genetic map of barley Franklin/YYXT DH population showing QTL identified following stagnant deoxygenated agar treatment in nutrient solution for 21 days. Arrows indicate QTL for RFW (root fresh weight), RDW (root dry weight), SFW (shoot fresh weight), SDW (shoot dry weight), LRL (longest root length) tiller number and root porosity in stagnant conditions
Other QTL for longest root length were identified on 2H at 156 cM (stagnant conditions) and 7H at 50.2 cM (aerated conditions) and explained 10.7 and 13.9 % of variation in longest root length, respectively. Two QTL for tiller number were identified on 3H, at 24.0 and 35.9 cM which explained 15.5–16.7 % of the variation in tiller number, and a third QTL for tiller number was identified on 4H. It explained 14.0 % variation in stagnant conditions, but it was not in the same region as the root porosity QTL. The tiller number QTL was located at 6.8 cM with closest marker bPb-1469, whilst the root porosity QTL was located at 116 cM. A second QTL for root weight was identified on 5H which explained 13.4 % of the variation in root fresh weight in stagnant conditions. Although plant growth traits were not mapped to the same location as the root porosity 4H QTL at LOD threshold value 3.0, reducing the LOD value (threshold LOD value 2.2 at P = 0.05) revealed a significant QTL for root fresh weight at the same location. The QTL was only identified in stagnant conditions and explained 7.6 % of the variation in root fresh weight at LOD 2.71 (data not shown).
Discussion
Oxygen deficiency increased root porosity and severely reduced plant growth and development in barley. Mean root porosity over all lines increased from 6.0 % in the aerated nutrient solution to 11.4 % in the stagnant deoxygenated agar nutrient solution; the increased gas-filled volume in roots of plants from the stagnant treatment resulted from the formation of lysigenous aerenchyma. The more tolerant parental line, YYXT, had greater root porosity (12.6 %) in the stagnant treatment compared with Franklin (10.2 %). In the aerated treatment, these two parent genotypes were not significantly different. This is similar to the preliminary screening experiment where significant differences in adventitious root porosity were also observed between YYXT and Franklin when in the stagnant treatment but not in the aerated treatment, although the genotypic differences in root porosity were greater in the earlier experiment (see “Results”).
A QTL was detected for root porosity on 4H which explained 35.7 and 39.0 % of phenotypic variation in aerated and stagnant conditions, respectively. The nearest marker to the QTL was EBmac0701. The Franklin/YYXT DH population had also been screened for waterlogging tolerance in a soil system (Zhou et al. 2012b). Plants were screened in stainless steel tanks filled with soil from a waterlogging prone area in Tasmania. Plants were waterlogged at the three-leaf stage and waterlogging was maintained for 9 weeks until susceptible lines died. Lines were rated at various stages after waterlogging for ‘plant health’, a combined score of leaf chlorosis and plant survival after waterlogging. Four significant QTL were identified on 2H, 3H, 4H and 6H which explained between 6.2 and 30.1 % of phenotypic variation. The QTL on 4H, Q WL .YyFr.4H (bPb-4244, 121 cM), accounted for 7.0 % of phenotypic variation and was located at the same position as the root porosity QTL identified in the present study (EBmac0701, 116 cM) indicating that the QTL for root porosity on 4H also contributed to waterlogging tolerance in soil. The fact that the 4H QTL explained more variation in the present study than in the earlier soil screening experiment of Zhou et al. (2012b) may be due to large differences between the two experiments. The soil screening experiment was designed to screen for whole-plant tolerance (plant health) in a simulated field environment following an extended period of waterlogging (up to 9 weeks). In contrast, the present study was run for a much shorter time in highly controlled conditions, using growth rooms and hydroponic systems, to focus on detailed measurements of the root porosity trait.
The correlation between waterlogging tolerance scores in the Franklin/YYXT DH population screened in soil in Tasmania and root porosity values in stagnant deoxygenated agar nutrient solution in the present study is positive but not strong (r = 0.148, P < 0.1). The relatively weak correlation may be due to the fact that the 4H QTL only explained 7.0 % of the variation in waterlogging tolerance in the Franklin/YYXT population, whilst the other three QTLs accounted for 52 % of the variation. When the soil waterlogging tolerance ratings and root porosity data from Franklin/YYXT were analysed together using MapQTL software, with the porosity data as a covariate, the QTL on 4H for waterlogging tolerance became non-significant, whilst other QTL remained unchanged, confirming that the 4H QTL is responsible for increased root porosity and improved waterlogging tolerance in barley.
Phenotypic data on waterlogging tolerance were also collected on a Franklin/Yerong DH population (Zhou 2011) using the same soil tank screening protocol as described in Zhou et al. (2012b). After 9 weeks of soil waterlogging, four significant QTL were detected with a major QTL on 4H (Q WL .YeFr.4H) accounting for 23.9 % of phenotypic variation. The same QTL was also detected at earlier stages of waterlogging. This QTL was located at 110 cM after waterlogging tolerance was mapped in the Franklin/Yerong population (Zhou 2011). Given that the closest marker for the root porosity QTL identified in the present study, EBmac0701, was located at 112 cM in the barley consensus map (Wenzl et al. 2006), it is likely that Yerong and YYXT share the same gene(s) for waterlogging tolerance on chromosome 4H.
Aerenchyma is a significant adaptive trait for waterlogging tolerance. Higher root porosity increases the movement of O2 into roots (Armstrong 1979), with benefits to cellular energy status (e.g. Zea mays roots, Drew et al. 1985), which in turn enhances root growth and tissue survival in anaerobic media, as well as root functioning in nutrient uptake and translocation to the shoot (reviewed by Colmer and Greenway 2011). In wheat, the importance of aerenchymatous adventitious roots for nutrient uptake from severely hypoxic medium has been demonstrated (Wiengweera and Greenway 2004), and positive correlations between aerenchyma amount and plant growth under waterlogged conditions have been observed (Huang et al. 1994; Setter et al. 1999). In the field study of Setter et al. (1999), wheat yields were positively correlated with the percentage of aerenchyma in adventitious roots under conditions of intermittent waterlogging. In the same study, relative grain yield for barley under waterlogging was not correlated with the amount of aerenchyma in adventitious roots. In the present study, however, increased porosity in adventitious roots was positively correlated with greater absolute root and shoot biomass, and relative root biomass, for barley grown in stagnant deoxygenated conditions.
In their review on waterlogging and germplasm improvement in wheat and barley, Setter and Waters (2003) stress the importance of environmental characterisation when screening for waterlogging tolerance as the timing, duration and intensity of waterlogging can vary significantly in different environments. Depending on the environment, the best strategy for plants may be to grow during waterlogging, or simply survive, then recover rapidly when waterlogging ceases. However, when waterlogging is for an extended period, they argue that there is evidence in support of aerenchyma as an adaptive trait in wheat and other temperate cereals for both continuous and intermittent waterlogged environments. This has been demonstrated in the present study where the putative root porosity allele (i.e. QTL on chromosome 4H) has also contributed to improved waterlogging tolerance in soil in both the Franklin/YYXT (Zhou et al. 2012b) and Franklin/Yerong populations (Zhou 2011). The fact that the QTL accounted for 7.0 and 23.9 %, respectively, of phenotypic variance in the two populations may reflect environmental differences as well as differing genetic backgrounds in the two populations.
Most of the other QTL identified in this study for traits such as root and shoot fresh/dry weights and tiller number do not align with other waterlogging tolerance QTL that have been identified in the Franklin/YYXT or Franklin/Yerong populations when phenotyped in the soil tank system (Zhou 2011; Zhou et al. 2012b). The major QTL for waterlogging tolerance in these populations is presented on a genetic map (Zhou et al. 2012b). Of the four QTL identified in the Franklin/YYXT population, the 4H QTL was located in a similar position to the root porosity QTL in this study. A putative root fresh weight QTL was also identified at the same location on 4H in the present study when a reduced LOD value of 2.71 was applied. The other three soil waterlogging tolerance QTL from Franklin/YYXT were located on 2H, 3H and 6H at 76.1, 5.2 and 78.4 cM (Zhou et al. 2012b), respectively, and were not located near any QTL identified in the present study.
Four major QTL controlling waterlogging tolerance on 2H, 3H and 4H were also identified in the Franklin/Yerong population (Zhou 2011). As described earlier, the 4H QTL for waterlogging tolerance in the Franklin/Yerong population was in a similar location to the 4H QTL identified in Franklin/YYXT for root porosity (present study) and soil waterlogging tolerance (Zhou et al. 2012b). Additionally, one of the QTL identified on 2H in the Franklin/Yerong population, Q WL .YeFr.2H.2, was located in a similar position to a QTL identified for longest adventitious root length in stagnant conditions in the present study. This 2H QTL for root length explained 10.7 % of variation and was located at 156 cM with closest marker bPb-3220. In the Franklin/Yerong population, the 2H QTL explained 17.2 % of variation for waterlogging tolerance (Zhou 2011). Thus results from the present study can be used to link traits to QTL that have been identified in other populations.
The potential to develop molecular markers for waterlogging tolerance in barley is promising. In the study involving the Franklin/Yerong population, Zhou (2011) demonstrated that the markers linked to the four QTL were effective in screening for waterlogging tolerance. When the marker for the 4H QTL was selected, the average score of the lines was 6.8 (rating 0–10 with 10 the most tolerant) which was significantly better than the average score of 4.8 from lines lacking the marker. When markers for all four QTL were selected, the average score increased to 7.6. In contrast when none of the markers were present, most of the lines were very susceptible with an average score of 1.4.
We explored possible synteny between the root porosity QTL identified in the present study and QTL associated with waterlogging/submergence tolerance in other species, in particular maize (Zea mays) and rice (Oryza sativa L.). In maize, there has been an effort to introduce aerenchyma, and flooding tolerance from the wild relative, teosinte, and QTL controlling aerenchyma formation was identified in an F2 population from a maize x teosinte cross. Four QTL for aerenchyma formation under non-flooded conditions were identified on two regions of chromosome 1, and on one region each of chromosomes 5 and 8; together these explained 46.5 % of the phenotypic variation (Mano et al. 2007). In rice, identification of the Sub1 QTL has enabled a breeding programme resulting in significant improvements in submergence tolerance of lowland rice. The locus was first identified in 1996 (Xu and Mackill 1996), and it contains a cluster of three ethylene responsive factor (ERF) genes, Sub1A, Sub1B and Sub1C with a variant in Sub1A being the key gene that confers a high degree of tolerance to complete submergence in rice. The Sub1A-1 gene has now been introgressed into at least eight lowland rice ‘mega varieties’, via marker-assisted backcrossing, and these submergence-tolerant varieties have been released in several countries in South Asia and Southeast Asia (Collard et al. 2013; Septiningsih et al. 2012).
The rice Sub1A-1 gene sequence (GenBank: FR720458.1) (Niroula et al. 2012) was used to blast the barley genome sequences using Viroblast (http://webblast.ipk-gatersleben.de/barley/viroblast.php). The results showed that one gene in Morex_contig_1565327 was localised to the same region as the root porosity QTL on 4H and was homologous to the Sub1A-1 gene on rice chromosome 9. The SSR marker GBM1220 next to EBmac0701 was anchored to the FPC_contig_43829. Morex_contig_1565327 and FPC_contig_43829 were anchored in the same region. This contig from Morex was then used to blast the maize genome, and its homologous region was anchored to short arm of chromosome 1 in maize where the QTL, Qaer1.02-3, for aerenchyma formation was mapped (Mano et al. 2007).
The comparative mapping revealed that the 4H root porosity QTL we identified in barley is syntenic with both the rice Sub1A-1 gene and the maize QTL, Qaer1.02-3. Both loci affect ethylene biosynthesis and responsiveness but in seemingly different ways. The presence of the Sub1A-1 gene in the submergence-tolerant rice genotypes suppresses the perception and production of ethylene via the induction of the Slender Rice-1 (SLR1) and SLR Like-1 (SLRL1) genes (Fukao and Bailey-Serres 2008). Accumulation of SLR1 and SLRL1 transcripts and proteins lead to the inhibition of GA-mediated shoot elongation and conservation of energy, a ‘quiescence’ strategy. This conserved energy helps the plant to survive and re-grow upon de-submergence. In contrast, the formation of lysigenous aerenchyma in crops such as rice and maize is generally promoted by the accumulation of endogenous ethylene which is stimulated by hypoxia (Justin and Armstrong 1991; Drew et al. 2000). In maize, there is also evidence that changes in ethylene sensitivity can affect aerenchyma formation, a response consistent with the possible involvement of an ERF gene. He et al. (1992) found that aerenchyma could be induced under well-oxygenated conditions in nutrient solution by the transient shortage of either an N or P source. They found the rate of ethylene synthesis declined, but that there was an increase in sensitivity to ethylene in the root tissue which stimulated aerenchyma formation. In a recent study on genes controlling aerenchyma formation in maize roots, Rajhi et al. (2011) identified a number of genes that related to a range of molecular functions including Ca2+ signalling, cell wall modification and the generation or scavenging of reactive oxygen species (ROS). The genes were up-regulated and down-regulated in root cortical cells under waterlogged conditions with evidence indicating their expression was regulated by ethylene. It is therefore conceivable that an ERF would be involved in the ethylene regulation of lysigenous aerenchyma formation in roots.
Data from the present study indicate that increased adventitious root porosity, presumably associated with development of aerenchyma, is positively correlated with root and shoot biomass in barley maintained in stagnant deoxygenated conditions. The 4H QTL identified in this study is located in the same region as waterlogging tolerance QTL identified from screening barley DH populations in a soil waterlogging tank system in Tasmania (Zhou 2011; Zhou et al. 2012b). This is the first report of a QTL for root porosity in barley.
References
Armstrong W (1979) Aeration in higher plants. Adv Bot Res 7:225–332
Arnell N, Liu C (2001) Hydrology and water resources. In: McCarthy JJ, Canziani OF, Leary NA, Dokken DJ, White KS (eds) Climate change 2001: impacts, adaptation and vulnerability, contribution of working group II to the third assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, pp 191–234
Collard BCY, Septiningsih EM, Das SR, Carandang JJ, Pamplona AM, Sanchez DL, Kato Y, Ye G, Reddy JN, Singh US, Iftekharuddaula KM, Venuprasad R, Vera-Cruz CN, Mackill DJ, Ismail AM (2013) Developing new flood-tolerant varieties at the International Rice Research Institute (IRRI). SABRAO J Breed Genet 45:42–56
Colmer TD (2003) Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots. Plant, Cell Environ 26:17–36
Colmer TD, Greenway H (2011) Ion transport in seminal and adventitious roots of cereals during O2 deficiency. J Exp Bot 62:39–57
Colmer TD, Voesenek LACJ (2009) Flooding tolerance: suites of plant traits in variable environments. Funct Plant Biol 36:665–681
CycDesigN 4.1 (2009) A package for the computer generation of experimental designs. CySoftware Ltd. http://www.vsni.co.uk/software/cycdesign
Drew MC, Saglio PH, Pradet A (1985) Larger adenylate energy charge and ATP/ADP ratios in aerenchymatous roots of Zea mays in anaerobic media as a consequence of improved internal oxygen transport. Plant Physiol 165:51–58
Drew MC, He CJ, Morgan PW (2000) Programmed cell death and aerenchyma formation in roots. Trends Plant Sci 5:123–127
FAO (2002) http://www.fao.org/waicent/FAOINFO/AGRICULT/agl/agll/gaezlnav.html on 18 March 2002
Fukao T, Bailey-Serres J (2008) Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellins responses in rice. Proc Natl Acad Sci USA 105:16814–16819
Gilmour AR, Cullis BR, Gogel B, Welham SJ, Thompson R (2005) ASReml, user guide. Release 2.0. VSN International Ltd, Hemel Hempstead
He CJ, Morgan PW, Drew MC (1992) Enhanced sensitivity to ethylene in nitrogen-starved or phosphate starved of Zea mays L. during aerenchyma formation. Plant Physiol 98:137–142
Huang BR, Johnson JW, Nesmith DS, Bridges DC (1994) Root and shoot growth of wheat genotypes in response to hypoxia and subsequent resumption of aeration. Crop Sci 34:1538–1544
Justin SHFW, Armstrong W (1987) The anatomical characteristics of roots and plant response to soil flooding. New Phytol 106:465–495
Justin SHFW, Armstrong W (1991) Evidence for the involvement of ethene in aerenchyma formation in adventitious roots of rice (Oryza sativa L.). New Phytol 118:49–62
Li H, Vaillancourt R, Mendham N, Zhou M (2008) Comparative mapping of quantitative trait loci associated with waterlogging tolerance in barley (Hordeum vulgare L.). BMC Genom 9:401
Mano Y, Omori F, Takamizo T, Kindiger B, Bird RM, Loaisiga CH, Takahashi H (2007) QTL mapping of root aerenchyma formation in seedlings of a maize x rare teosinte “Zea nicaraguensis” cross. Plant Soil 295:103–113
Mano Y, Omori F, Kindiger B, Takahashi H (2008) A linkage map of maize x teosinte Zea luxurians and identification of QTLs controlling root arenchyma formation. Mol Breed 21:237–337
Niroula RK, Pucciariello C, Ho VT, Novi G, Fukao T, Perata P (2012) SUB1A-dependent and -independent mechanisms are involved in the flooding tolerance of wild rice species. Plant J 72:282–293
Pang J, Zhou M, Mendham N, Shabala S (2004) Growth and physiological responses of six barley genotypes to waterlogging and subsequent recovery. Aust J Agric Res 55:895–906
Ponnamperuma F (1984) Effects of flooding on soils. In: Kozlowski T (ed) Flooding and plant growth. Academic Press, New York, pp 9–45
Qiu JD, Ke Y (1991) A study of determination of wet tolerance of 4572 barley germplasm resources. Acta Agric Shanghai 7:27–32
Rajhi I, Yamauchi T, Takahashi H, Nishiuchi S, Shiono K, Watanabe R, Mliki A, Nagamura Y, Tsutsumi N, Nishizawa NK, Nakazono M (2011) Identification of genes expressed in maize root cortical cells during lysigenous aerenchyma formation using laser microdissection and microarray analyses. New Phytol 190:351–368
Raskin I (1983) A method for measuring leaf volume, density, thickness and internal gas volume. HortScience 18:698–699
R Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, http://www.R-project.org/
Septiningsih EM, Sanchez DL, Singh N, Sendon PMD, Pamplona AM, Heuer S, Mackill DJ (2012) Identifying novel QTLs for submergence tolerance in rice cultivars IR72 and Madabaru. Theor Appl Genet 124:867–874
Setter T, Waters I (2003) Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil 253:1–34
Setter TL, Burgess P, Waters I, Kuo J (1999) Genetic diversity of barley and wheat for waterlogging tolerance in Western Australia. In: Proceedings of the 9th Australian barley technical symposium, Melbourne, Australia, 12–16 September 1999, pp 2.17.1–2.17.7
Shabala S (2011) Physiological and cellular aspects of phytotoxicity tolerance in plants: the role of membrane transporters and implications for crop breeding for waterlogging tolerance. New Phytol 190:289–298
Takeda K, Fukuyama T (1986) Variation and geographical distribution of varieties for flooding tolerance in barley seeds. Barley Genet Newsl 16:28–29
Thomson CJ, Armstrong W, Waters I, Greenway H (1990) Aerenchyma formation and associated oxygen movement in seminal and nodal roots of wheat. Plant, Cell Environ 13:395–404
Van Ooijen JW (2004) MapQTL 5.0 Software for the mapping of quantitative trait loci in experimental populations. Kyazma BV, Wageningen
Van Ooijen JW (2006) JoinMap 4.0 Software for the calculation of genetic linkage maps in experimental populations. Kyazma BV, Wageningen
Varshney RK, Marcel TC, Ramsay L, Russell J, Roder MS, Stein N, Waugh R, Langridge P, Niks RE, Graner A (2007) A high density barley microsatellite consensus map with 775 SSR loci. Theor Appl Genet 114:1091–1103
Wenzl P, Li HB, Carling J, Zhou MX, Raman H, Paul E, Hearnden P, Maier C, Xia L, Caig V, Ovesna J, Cakir M, Poulsen D, Wang JP, Raman R, Smith KP, Muehlbauer GJ, Chalmers KJ, Kleinhofs A, Huttner E, Kilian A (2006) A high-density consensus map of barley linking DArT markers to SSR, RFLP and STS loci and agricultural traits. BMC Genom 7:206
Wiengweera A, Greenway H (2004) Performance of seminal and nodal roots of wheat in stagnant solution: K+ and P uptake and effects of increasing O2 partial pressures around the shoot on nodal root elongation. J Exp Bot 55:2121–2129
Wiengweera A, Greenway H, Thomson CJ (1997) The use of agar nutrient solution to simulate lack of convection in waterlogged soils. Ann Bot 80:115–123
Xu K, Mackill DJ (1996) A major locus for submergence tolerance mapped on rice chromosome 9. Mol Breed 2:219–224
Zhou M (2011) Accurate phenotyping reveals better QTLs for waterlogging tolerance in barley. Plant Breed 130:203–208
Zhou MX, Li HB, Mendham NJ (2007) Combining ability of waterlogging tolerance in barley. Crop Sci 47:278–284
Zhou GF, Johnson P, Ryan PR, Delhaize E, Zhou MX (2012a) Quantitative trait loci for salinity tolerance in barley (Hordeum vulgare L.). Mol Breed 29:427–436
Zhou MX, Johnson P, Zhou GF, Li CD (2012b) Lance R (2012b) Quantitative trait loci for waterlogging tolerance in a barley cross of Franklin × YuYaoXiangTian Erleng and the relationship between waterlogging and salinity tolerance. Crop Sci 52:2082–2088
Acknowledgments
Funding support from the Australian Grains Research and Development Corporation through project DAW00187 ‘DAFWA-TIAR China Barley Collaboration’ is gratefully acknowledged. We would also like to thank Sharon Westcott and Vera Limadinata for assisting with root porosity and plant growth measurements.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Broughton, S., Zhou, G., Teakle, N.L. et al. Waterlogging tolerance is associated with root porosity in barley (Hordeum vulgare L.). Mol Breeding 35, 27 (2015). https://doi.org/10.1007/s11032-015-0243-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11032-015-0243-3