
Abstract
It has become evident that AKT inhibitors have great potential in cancer treatment. In this study, we investigate the anticancer activity of MK-2206, a novel AKT inhibitor, on HepG2 hepatocellular carcinoma cell, and to show whether MK-2206 enhances the apoptosis-inducing potential of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). The cell growth inhibition was evaluated by MTT assay and colony formation assay. Cell cycle distribution was assessed by propidium iodide flow cytometry. Apoptosis was determined by AnnexinV-FITC/PI double staining assay and caspase-9, casapse-7, caspase-3, and PARP cleavage. The results of present study showed that MK-2206-induced G1-phase arrest was associated with a marked decrease in the protein expression of cyclin D1 with concomitant induction of p21 and p27. MK-2206-induced apoptosis was characterized by cleavage of a pro-caspase in a concentration-dependent manner. Moreover, the MAP family kinases p38 kinase and JNK were activated by exposure to MK-2206. SB203580, an p38-specific inhibitor, partially blocked MK-2206-induced death of HepG2 cells and caspase activation. A combination of MK-2206 with TRAIL significantly inhibited growth of TRAIL resistant HepG2 cells. Taken together, our findings provide a new insight to better understand anticancer mechanisms of MK-2206, at least in HepG2 cell. Using of MK-2206 as a potent sensitizer to TRAIL-induced apoptotic cell death offers a promising means of enhancing the efficacy of TRAIL-based HCC treatments.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Hepatocellular carcinoma (HCC) accounts for up to 70 % of cancer deaths in Eastern Asia and Central Africa, being the most common malignant hepatobiliar disease [1, 2]. Nearly 85 % of these cases occur in less developed countries, with China alone accounting for more than 50 % of the total [3]. The main cause of HCC is the cirrhotic liver, which associates with chronic hepatitis B virus (HBV) or hepatitis C virus (HCV) infections and the consumption of alcohol. Dysregulation of cellular proliferation and apoptosis are frequent events related with malignant phenotype and poor responsiveness of HCC toward chemotherapy [4]. Therefore, developing effective pharmacological therapy to these dysregulation is a valid target for HCC clinical treatment.
The phosphatidylinositol3-kinase (PI3K)/AKT pathway is activated in the majority of human cancers [5, 6]. AKT, also known as PKB, is a serine/threonine protein kinase, which comprises three related isoforms AKT1, AKT2, AKT3, and functions as a key regulator of cell proliferation, survival, and oncogenesis [7, 8]. The initiation and development of cancer are the results of a disturbance in the balance between cell proliferation and apoptosis. Actually, PI3K/AKT signaling is associated with both of these events, and plays an important role both in tumor growth and in the potential response of a tumor to cancer treatment [9]. Recent studies have indicated that unregulated activation of the PI3K/AKT pathway is a prominent feature of many human cancers, and AKT is considered as an attractive target for cancer therapy [10]. Several small molecules designed to specifically target PI3K/AKT have been developed. MK-2206, an oral small molecule allosteric inhibitor of AKT, is being tested in preclinical settings as an anticancer agent. The activation of MK-2206 against glioma cells has been confirmed in vitro [11, 12].
As a member of the tumor necrosis factor (TNF) superfamily, tumor necrosis factor-related apoptosis inducing ligand (TRAIL), is considered as one of the most promising candidates for the treatment of cancers, as it is able to selectively induce apoptosis in tumor cells without affecting normal cells [13]. However, previous work indicated that tumors cells, including HCC cells, were found to show resistance to TRAIL-induced apoptosis [14]. Recent studies have shown that chemotherapeutic agents can increase the sensitivity to TRAIL by increasing the activation of caspase or by downregulating antiapoptotic proteins in a range of tumors, suggesting that a synergistic antitumor effect may be achieved by using combination therapies to increase the apoptotic effects of TRAIL [15, 16]. Treatment of TRAIL-resistant tumors with inhibitor of PI3K/AKT pathway restores the sensitivity to TRAIL-induced apoptosis [17].
The purpose of the present study is to investigate the anti-cancer effects of MK-2206 on human HCC HepG2 cell proliferation and apoptosis, and to explore the molecular mechanisms. Also, we tested whether MK-2206 potentiated TRAIL-induced HepG2 cell apoptosis. We found that combined treatment of HepG2 cell with MK-2206 and TRAIL induced marked cell apoptosis.
Materials and methods
Reagents
MK-2206 was obtained from Selleck Chemicals, and dissolved with DMSO. TRAIL was purchased from Peprotech (Rocky Hill, NJ). SP600125 and SB203580 were purchased from Calbiochem (La Jolla, CA, USA). 3-(4,5-dimethylthiazol-2yl)-2,5-diphenyl tetrazolium bromide (MTT) and Propidium iodide (PI) were purchased from Sigma-Aldrich (St. Louis, MO, USA). The Annexin V-FITC/PI apoptosis detection kit was purchased from KeyGEN Biotech (Nan Jing, China).
Antibodies specific to β-actin was obtained from Sigma-Aldrich. Rabbit anti-AKT, p-AKT, caspase-9, caspase-7, caspase-3, PARP antibodies were purchased from Cell Signaling Technology (Shanghai, China). Mouse anti-cyclinD1, p38, and rabbit anti-cyclinB1 antibodies were purchased from Santa cruz Technology (CA, USA). Mouse anti-p21, p27, and p-JNK antibodies were purchased from BD Biosciences (California, USA). Rabbit anti-JNK antibody was purchased from Signalway Antibody (Shanghai, China).
Cell culture
HepG2 cell line was purchased from the Cell Bank of the Chinese academy of Sciences (Shanghai, China), and cultured in RPMI 1640 medium supplemented with 10 % fetal bovine serum (GIBCO, USA) at 37 °C containing 5 % CO2.
MTT assay
Cells were seeded in 96-well plates overnight, and then incubated with different concentrations of MK-2206 for 24 h. Each drug concentration was performed in six replicate wells. A negative control group without drugs was also established. At the end of the treatment, 20 μL MTT solution (5 mg/mL) was added to each well and incubated at 37 °C for a further 4 h. Then 150 μL of DMSO was added to each well after the medium was removed and mixed by agitation at room temperature for 20 min. The optical density (OD) values were measured at 490 nm on a scanning multiwall spectrophotometer (BioRad Model 550, USA).
Clone formation assay
The cells were seeded at 4 × 103cells/well in 6-well plates and incubated at 37 °C for 14 days. After washing twice with PBS and fixing in 4 % paraformaldehyde fixative for 10 min, cells were stained with crystal violet for 10 min. Colonies with ≥50 cells were counted under an inverted microscope.
Cell cycle evaluation
Cells were plated onto 6-well tissue culture plates. After culturing for 24 h, the medium was changed and the drugs were added. Cells were incubated for an additional 24 h before they were harvested. Control and treated cells were collected into flow cytometry tubes and centrifuged at 1,000 rpm for 5 min to obtain cell pellets. The cells were washed in PBS, then fixed in 75 % ethanol for 24 h at 4 °C. The fixed cells were washed three times, and resuspended in PBS containing 10 μg/mL of RNase A for 30 min, and then incubated with 10 μg/mL ropidium iodide (PI) for 15 min in the dark. Subsequently, the samples were acquired in a FACScalibur system by cellquest software. For each measurement, at least 20,000 cells were acquired. Analysis was performed by Modfit software.
Flow cytometry detection of apoptotic cells
The total number of cells, both adherent and suspended, were collected and washed twice with PBS. Annexin V-FITC/PI apoptosis detection kit was used according to the manufacturer’s protocol. After the Annexin V-FITC was added into the labeled tube 5 min, PI was also added and incubated for 10 min in the dark. Samples were analyzed with the FACScan flow cytometry and CellQuest analysis software (Becton–Dickinson). Data analysis was performed using Cellquest software.
Western blot analysis
After 24 h of MK-2206 treatment (0–20 μM), cells were harvested and washed twice with PBS. Total protein was obtained by lysing in ice-cold RIPA buffer for 20 min, and the supernatant containing proteins were collected by centrifugation at 13,000×g for 30 min. The protein concentration of the supernatant was determined using the BCA protein assay. For western blot analysis, 30 μg of total protein sample was loaded over a SDS-PAGE and transferred to polyvinylidine difluoride filter membranes (Millipore, Bedford, MA). The membranes were incubated overnight at 4 °C with the specific primary antibodies. Horseradish peroxidase (HRP)-conjugated secondary antibody was diluted in 1 % BSA/Tween-TBS. Detection of the target proteins on the membranes was performed using the ECL Western Blot Detection Reagents (Thermo Scientific Pierce, USA).
Statistical analysis
Statistical analysis was performed using SPSS 16.0 software. Results are shown as the mean ± standard deviation (SD) and P values <0.05 was considered statistically significant. Each experiment consisted of at least three replicates per condition.
Results
MK-2206 downregulates the phosphorylation of AKT in HepG2 cell
Since MK-2206 is designed to be an AKT inhibitor, we performed western blot analysis to confirm the inhibitory effect of this compound on the phosphorylation of AKT in HepG2 cells. As shown in Fig. 1, MK-2206 reduced the levels of S473p-AKT in a dose-dependent manner. No significant change of total AKT protein level was observed.

Western blot assay determines the expression of p-AKT and total AKT levels in HepG2 cells. Cells were treated with the increasing concentrations of MK-2206 for 24 h, after which protein lysates were prepared and subjected to protein gel blot analysis using the indicated antibodies. The expression of p-AKT and AKT were assessed by western blot analysis in HepG2 cells. β-Actin was used as an internal standard
MK-2206 inhibits hepatocellular carcinoma HepG2 cell proliferation
MTT results indicated that the cell viability of the MK-2206 group showed a significant decrease after HepG2 cells were incubated with MK-2206 for 24 h (Fig. 2a). To test the inhibitory effect of MK-2206 on growth kinetics of HepG2 cells, the colony formation assay was performed. Cells were treated with increasing concentration of MK-2206 (1–10 μM) as the treatment group and treated with DMSO as the control group. After 14 days of culture, we statistically accounted the percentage of colony formation. From Fig. 2b, we found that the colony formation of HepG2 cell line was dramatically suppressed upon MK-2206 treatment.

MK-2206 inhibits HepG2 cell growth. a MK-2206 reduces the viability of HepG2 cells. Cells were seeded a in 96-well culture plates at 37 °C with 5 % (v/v) CO2. After 24 h of incubation, cells were incubated with indicated concentrations of MK-2206 for 24 h and then processed for MTT assay. b MK-2206 inhibits the colony formation ability of HepG2 cells. After treatment with MK-2206 for 2 weeks, the cells were washed with PBS for two times, fixed with 4 % paraformaldehyde fixative for 10 min, stained with crystal violet for 10 min, washed again, and photographed. c MK-2206 treatment results in the cell cycle arrest of G1. HepG2 cells were exposed to different concentrations of MK-2206 for 24 h, then cell cycle analysis was determined by flow cytometry. d MK-2206 down-regulates cyclinD1 level and up-regulates p21 and p27 expression in HepG2 cells. Cells were cultured for 24 h in the presence of increasing doses of MK-2206
A variety of cytotoxic anticancer drugs are known to affect cell proliferation by disturbing cell cycle progression, so the effect of MK-2206 on HepG2 cell cycle distribution was examined after propidium iodide staining. As shown in Fig. 2c, compared with control group, cell cycle arrest in G1 phase was more pronounced by treatment with MK-2206 (52.76 % for control, 60.65 % for 1 μM MK-2206, 71.79 % for 2.5 μM MK-2206, and 72.97 % for 5 μM MK-2206), indicating concentration-dependent G1 phase arrest.
The protein levels of certain cyclins and cyclin-dependent kinase inhibitors (CKIs) family members, crucially required for the regulation of G1 phase progression, were determined by western blot analysis. Results showed that MK-2206 downregulated the expression of the CDK regulator cyclinD1, and upregulated the expression of p21 and p27 in a dose-dependent manner (Fig. 2d). In contrast, the cyclinB1 expression levels remained unchanged in MK-2206-treated HepG2 cells.
MK-2206 induces HepG2 cell apoptosis by activation of a cascade of caspase
To investigate whether the growth inhibitory effect was mediated through the induction of apoptosis, AnnexinV-FITC/PI double staining assay was used to assess the apoptosis-inducing effect of MK-2206. We found that HepG2 cells treated with MK-2206 displayed much higher apoptosis rate than the control group.
To explore whether the apoptotic effect of MK-2206 was through activation of a cascade of caspase, the cleavage of caspase-9, caspase-7, caspase-3, and PARP was detected by western blot analysis. The results showed that MK-2206 increased the cleavage of caspase-9, caspase-7, and caspase-3 in a concentration-dependent manner (Fig. 3b). The cleavage of PARP, a sensitive substrate of active caspase-3, was also observed with parallel activation of caspases. These data suggest that MK-2206 is a potent inducer of apoptosis in HepG2 cancer cells.

MK-2206 induces apoptosis in HepG2 cells. a HepG2 cells were incubated with the indicated concentrations of MK-2206 for 24 h, the percentage of dead cells was determined using the Annexin V staining assay. b MK-2206 treatment induced pro-caspase-9, pro-caspase-7, pro-caspase-3, and PARP cleavage. Equal amounts of whole-cell lysates were subjected to 15 % SDS-PAGE and immunoblotted with various antibodies, as indicated. β-Actin was used as an internal standard
Activation of p38 MAPK is required for MK-2206-reduced cell viability
Mitogen-activated protein kinases have been implicated in a variety of cellular processes, including cell cycle control and apoptosis. The role of MAPKs signal transduction events induced by MK-2206 that could contribute to apoptosis were investigated in this study. We found that MK-2206 induced dose-related activation of JNK and p38 MAPK in HepG2 cells (Fig. 4a).

MK-2206 increases the activation of JNK and p38 in HepG2 cells. a HepG2 cells were exposed to MK-2206 at the indicated concentrations (0, 1, 5, 10, 20 μM) for 24 h, and p-JNK, p-p38, JNK, and p38 were identified by western blot analysis, as described in "Materials and methods" section. b HepG2 cells were exposed to 20 μM of MK-2206 for 24 h and MTT analysis was performed in the presence or absence of SB203580 (20 μM) and SP600125 (20 μM). * P < 0.05 compared with the control group. c To investigate the effect of the p38 inhibitor on CCT128930-induced caspase cleavage, HepG2 cells were pre-treated as before with 20 μM SB203580. After 24 h incubation with MK-2206 (20 μM), protein extracts of HepG2 cells were analyzed for caspases activation using western blot analysis
To address the role of activated JNK and p38 in MK-2206-reduced cell viability, HepG2 cells were treated with 20 μM MK-2206 in the presence or absence of SP600125 and SB203580 for 24 h. Results showed that MK-2206-reduced viability and caused cell death was significantly blocked in the pretreatment of SB203580, an p38-specific inhibitor (Fig. 4b). Moreover, MK-2206-induced cleavages of apical pro-caspase-3, pro-caspase-9, and PARP into the characteristic activate fragments were partially reduced in the presence of SB203580 (Fig. 4c). All of these data suggest that MK-2206 mediates the proliferation and apoptosis of HepG2 cells via the p38 MAPK pathway.
MK-2206 sensitizes HepG2 cells to TRAIL-mediated apoptosis
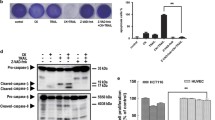
To investigate whether the combination treatment of MK-2206 and TRAIL has an effect on HepG2 cell survival, we cotreated HepG2 cells with MK-2206 and TRAIL for 24 h. As shown in Fig. 5a, cotreatment with MK-2206 significantly enhanced TRAIL-induced cytotoxicity in HepG2 cells, as evaluated by MTT assays. To confirm that the enhanced cytotoxic effect by MK-2206 was due to increased apoptosis, we detected the apoptotic response using fluorescein labeled Annexin V. When cells were analyzed by flow cytometry, it became evident that the combined TRAIL and MK-2206 treatment induced apoptotic cell death of HepG2 cells; whereas when the TRAIL or MK-2206 was used alone, much lower effects were observed (Fig. 5b).

MK-2206 enhances TRAIL-induced cytotoxicity in HepG2 cells. a Cells were cotreated with 100 ng/mL TRAIL and 15 μM MK-2206 for 24 h, then cell viability was determined by MTT assay, as described in "Materials and methods" section. Data are mean ± SD and representative of three independent experiments. * P < 0.05 compared with the control group. # P < 0.05 compared with the MK-2206 group. b HepG2 cells were treated with TRAIL alone or together with MK-2206 for 24 h. After treatment, apoptosis was detected by flow cytometry. The number of apoptotic cells was expressed as % of total cell number. c HepG2 cells were treated with TRAIL with or without MK-2206 for 24 h. Whole-cell extracts were prepared and analyzed by western blot using antibodies against PARP, caspase-3, and caspase-9. β-Actin was served as a loading control
Caspase activation is associated with the synergistic antitumor effect of chemotherapeutic agents and TRAIL. Therefore, we examined whether caspases are activated during the induction of apoptosis in HepG2 cells by the combined treatment with TRAIL and MK-2206. We found that although TRAIL and MK-2206 alone had little effect on the activation of caspase-9, caspase-3, and on the cleavage of PARP, their combination significantly induced the cleavage of caspase-9, caspase-3, and PARP (Fig. 5c).
Discussion
The PI3K/AKT signaling pathway has been well documented as playing a major role in carcinogenesis and drug resistance in HCC cells [18]. Studies have shown that tumors accompanied by the activation of PI3 K/AKT signaling become more aggressive and are associated with poor prognosis in patients with HCC [19]. MK-2206, a highly selective inhibitor of all AKT isoforms, is currently undergoing clinical trials for the treatment of solid tumors [20]. Previous studies have found that MK-2206 is effective in several human cancer models by inducing autophagy and apoptosis [21, 22]. However, the inhibitory effects and detailed molecular mechanisms by which MK-2206 contributes to HepG2 cells have not been fully illustrated.
Cell cycle arrest in cancer cells has been known as a major indicator for anticancer effect, the loss of cell cycle control has been implicated in tumor development and proliferation [23]. In this study, MK-2206 induced a significant cell-cycle arrest in G1 phase. Cyclin D1, a cyclin required for G1 to S transition, which was found to be reduced upon MK-2206 treatment could be the main cause for the G1 block. Our data also suggested that the levels of p21 and p27, two members of the Cip/Kip family of CDK inhibitors, were upregulated by MK-2206 treatment.
Apoptosis, a naturally occurring process important for clearing damaged or infected cells, is the induction of cell suicide and can be triggered by either intrinsic cues or activation of the relevant signaling pathways. From apoptosis assessment assays and western blot analysis, we showed that the expression levels of cleaved caspase-3, -7, and -9 proteins, activated forms, were dramatically increased in MK-2206-treatment cells. Since the cleavage of PARP is one of the markers observed during apoptosis, we also examined the protein state of PARP. As shown in Fig. 3b, PARP mainly existed as full length form in control cells, while cleaved PARP fragments were detected in cells treated with MK-2206.
Signal transduction pathways involving the mitogen-activated protein kinases (MAPKs) including the ERKs, JNKs, and p38 MAPK, have been shown to differentially contribute to pro- and anti-apoptotic pathways [24, 25]. To gain further insight in the mechanism of MK-2206-induced death of HepG2 cells, we examined the possible involvement of members of the MAPK subfamily. We observed that both p38 MAPK and JNK were activated in the HepG2 cells treated with MK-2206. In the presence of the p38 MAPK inhibitor SB203580, not the presence of JNK inhibitor SP600125, MK-2206-induced downregulation of cell viability, and caspase cleavage were inhibited (Fig. 4b), implying that MK-2206-induced JNK activation may be involved in other biological processes in hepatoma cells. Moreover, further experiments are needed to know how p38 MAPK gives a signal to regulate the MK-2206-induced apoptosis of HepG2 cell.
TRAIL, a member of the TNF-superfamily, was originally reported to induce apoptosis in a wide range of tumor cell types but not in normal cells, and thus represents a promising anticancer cytokine [26, 27]. However, a number of in vitro studies have convincingly shown that some types of cancer cells, including hepatoma cell lines, are resistant to the apoptotic effects of TRAIL [28, 29]. But combinatorial approaches based on TRAIL and different chemotherapeutic agents, such as small molecule inhibitors and drugs, have been developed to overcome the resistance of cancer cells to TRAIL [30]. A recent report has highlighted that TRAIL sensitivity of acute myelogenous leukemia (AML) cells could be considerably increased by cotreatment with the AKT inhibitor, perifosine [31]. Therefore, we investigate whether the novel AKT inhibitor MK-2206 exhibits synergistic activity against the HepG2 cell when combined with TRAIL. From the Fig. 5, we found that MK-2206 sensitized HepG2 cells to TRAIL-induced apoptosis. The combined therapy of MK-2206 and TRAIL appears to be a more powerful modality compared with treatment using a single agent against HCC.
In summary, we have demonstrated that AKT inhibitor MK-2206 has important effects on HepG2 cells by inhibiting cell growth and by inducing cell apoptosis. This was mediated by the activation of caspase and p38-dependent cell death. In addition, the combination of MK-2206 and TRAIL could represent a novel strategy for overcoming TRAIL resistance, and combined treatment with MK-2206 and TRAIL may be an effective strategy for human liver cancer therapy.
References
Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D (2011) Global cancer statistics. CA Cancer J Clin 61(2):69–90. doi:10.3322/caac.20107
Jemal A, Center MM, DeSantis C, Ward EM (2010) Global patterns of cancer incidence and mortality rates and trends. Cancer Epidemiol Biomarkers Prev 19(8):1893–1907. doi:10.1158/1055-9965
El-Serag HB (2011) Hepatocellular carcinoma. N Engl J Med 365(12):1118–1127. doi:10.1056/NEJMra1001683
Carbajo-Pescador S, Steinmetz C, Kashyap A, Lorenz S, Mauriz JL, Heise M, Galle PR, González-Gallego J, Strand S (2013) Melatonin induces transcriptional regulation of Bim by FoxO3a in HepG2 cells. Br J Cancer 108(2):442–449. doi:10.1038/bjc.2012.563
Altomare DA, Khaled AR (2012) Homeostasis and the importance for a balance between AKT/mTOR activity and intracellular signaling. Curr Med Chem 19(22):3748–3762. doi:CMC-EPUB-20120607-24
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS, Anderson MJ, Arden KC, Blenis J, Greenberg ME (1999) Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 96(6):857–868. doi:S0092-8674(00)80595-4
Choudhury GG, Mahimainathan L, Das F, Venkatesan B, Ghosh-Choudhury N (2006) c-Src couples PI3 kinase/Akt and MAPK signaling to PDGF-induced DNA synthesis in mesangial cells. Cell Signal 18(11):1854–1864. doi:10.1016/j.cellsig.2006.02.003
Cheng GZ, Park S, Shu S, He L, Kong W, Zhang W, Yuan Z, Wang LH, Cheng JQ (2008) Advances of AKT pathway in human oncogenesis and as a target for anti-cancer drug discovery. Curr Cancer Drug Targets 8(1):2–6
Osaki M, Oshimura M, Ito H (2004) PI3 K-Akt pathway: its functions and alterations in human cancer. Apoptosis 9(6):667–676. doi:10.1023/B:APPT.0000045801.15585.dd
Lindsley CW (2010) The Akt/PKB family of protein kinases: a review of small molecule inhibitors and progress towards target validation: a 2009 update. Curr Top Med Chem 10(4):458–477
Cheng Y, Ren X, Zhang Y, Patel R, Sharma A, Wu H, Robertson GP, Yan L, Rubin E, Yang JM (2011) eEF-2 kinase dictates cross-talk between autophagy and apoptosis induced by Akt Inhibition, thereby modulating cytotoxicity of novel Akt inhibitor MK-2206. Cancer Res Cancer Res 71(7):2654–2663. doi:10.1158/0008-5472.CAN-10-2889
Jin R, Nakada M, Teng L, Furuta T, Sabit H, Hayashi Y, Demuth T, Hirao A, Sato H, Zhao G, Hamada JI (2013) Combination therapy using notch and akt inhibitors is effective for suppressing invasion but not proliferation in glioma cells. Neurosci Lett 534:316–321. doi:10.1016/j.neulet.2012.12.008
Lan Y, Liu X, Zhang R, Wang K, Wang Y (2013) Hua ZC (2013) Lithium enhances TRAIL-induced apoptosis in human lung carcinoma A549 cells. Biometals. doi:10.1007/s10534-012-9607-x
Carlisi D, D’Anneo A, Angileri L, Lauricella M, Emanuele S, Santulli A, Vento R, Tesoriere G (2011) Parthenolide sensitizes hepatocellular carcinoma cells to TRAIL by inducing the expression of death receptors through inhibition of STAT3 activation. J Cell Physiol 226(6):1632–1641. doi:10.1002/jcp.22494
Charette N, De Saeger C, Horsmans Y, Leclercq I, Stärkel P (2013) Salirasib sensitizes hepatocarcinoma cells to TRAIL-induced apoptosis through DR5 and survivin-dependent mechanisms. Cell Death Dis 4:e471. doi:10.1038/cddis.2012.200
Woo JS, Kim SM, Jeong CH, Ryu CH, Jeun SS (2013) Lipoxygenase inhibitor MK886 potentiates TRAIL-induced apoptosis through CHOP- and p38 MAPK-mediated up-regulation of death receptor 5 in malignant glioma. Biochem Biophys Res Commun 431(2):354–359. doi:10.1016/j.bbrc.2012.11.134
Wang S (2008) The promise of cancer therapeutics targeting the TNF-related apoptosis-inducing ligand and TRAIL receptor pathway. Oncogene 27(48):6207–6215. doi:10.1038/onc.2008.298
Cully M, You H, Levine AJ, Mak TW (2006) Beyond PTEN mutations: the PI3K pathway as an integrator of multiple inputs during tumorigenesis. Nat Rev Cancer 6(3):184–192. doi:10.1038/nrc1819
Jung KH, Zheng HM, Jeong Y, Choi MJ, Lee H, Hong SW, Lee HS, Son MK, Lee S, Hong S, Hong SS (2013) Suppression of tumor proliferation and angiogenesis of hepatocellular carcinoma by HS-104, a novel phosphoinositide 3-kinase inhibitor. Cancer Lett 328(1):176–187. doi:10.1016/j.canlet.2012.08.005
Somnay Y, Simon K, Harrison AD, Kunnimalaiyaan S, Chen H, Kunnimalaiyaan M (2013) Neuroendocrine phenotype alteration and growth suppression through apoptosis by MK-2206, an allosteric inhibitor of AKT, in carcinoid cell lines in vitro. Anticancer Drugs 24(1):66–72. doi:10.1097/CAD.0b013e3283584f75
Liu R, Liu D, Trink E, Bojdani E, Ning G, Xing M (2011) The Akt-specific inhibitor MK2206 selectively inhibits thyroid cancer cells harboring mutations that can activate the PI3K/Akt pathway. J Clin Endocrinol Metab 96(4):E577–E585. doi:10.1210/jc.2010-2644
Sangai T, Akcakanat A, Chen H, Tarco E, Wu Y, Do KA, Miller TW, Arteaga CL, Mills GB, Gonzalez-Angulo AM, Meric-Bernstam F (2012) Biomarkers of response to akt inhibitor MK-2206 in breast cancer. Clin Cancer Res 18(20):5816–5828. doi:10.1158/1078-0432.CCR-12-1141
Liu Y, Cao Y, Zhang W, Bergmeier S, Qian Y, Akbar H, Colvin R, Ding J, Tong L, Wu S, Hines J, Chen X (2012) A small-molecule inhibitor of glucose transporter 1 downregulates glycolysis, induces cell-cycle arrest, and inhibits cancer cell growth in vitro and in vivo. Mol Cancer Ther 11(8):1672–1682. doi:10.1158/1535-7163.MCT-12-0131
Widmann C, Gibson S, Johnson GL (1998) Caspase-dependent cleavage of signaling proteins during apoptosis. A turn-off mechanism for anti-apoptotic signals. J Biol Chem 273(12):7141–7147
Chun J, Joo EJ, Kang M, Kim YS (2013) Platycodin D induces anoikis and caspase-mediated apoptosis via p38 MAPK in AGS human gastric cancer cells. Cell Biochem 114(2):456–470. doi:10.1002/jcb.24386
Siegelin MD (2012) Utilization of the cellular stress response to sensitize cancer cells to TRAIL-mediated apoptosis. Expert Opin Ther Targets 16(8):801–817. doi:10.1517/14728222.2012.703655
Hellwig CT, Rehm M (2012) TRAIL signaling and synergy mechanisms used in TRAIL-based combination therapies. Mol Cancer Ther 11(1):3–13. doi:10.1158/1535-7163.MCT-11-0434
Maksimovic-Ivanic D, Stosic-Grujicic S, Nicoletti F, Mijatovic S (2012) Resistance to TRAIL and how to surmount it. Immunol Res 52(1–2):157–168. doi:10.1007/s12026-012-8284-8
Abou El Naga RN, Azab SS, El-Demerdash E, Shaarawy S, El-Merzabani M, Ammar el-SM (2013) Sensitization of TRAIL-induced apoptosis in human hepatocellular carcinoma HepG2 cells by phytochemicals. Life Sci 92(10):555–561. doi:10.1016/j.lfs.2013.01.017
Wang G, Wang X, Yu H, Wei S, Williams N, Holmes DL, Halfmann R, Naidoo J, Wang L, Li L, Chen S, Harran P, Lei X, Wang X (2013) Small-molecule activation of the TRAIL receptor DR5 in human cancer cells. Nat Chem Biol 9(2):84–89. doi:10.1038/nchembio.1153
Tazzari PL, Tabellini G, Ricci F, Papa V, Bortul R, Chiarini F, Evangelisti C, Martinelli G, Bontadini A, Cocco L, McCubrey JA, Martelli AM (2008) Synergistic proapoptotic activity of recombinant TRAIL plus the Akt inhibitor Perifosine in acute myelogenous leukemia cells. Cancer Res 68(22):9394–9403. doi:10.1158/0008-5472.CAN-08-2815
Acknowledgments
This study was supported by Grants from the National Natural Science Foundation of China (No. 81272683).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jiao, P., Zhou, YS., Yang, JX. et al. MK-2206 induces cell cycle arrest and apoptosis in HepG2 cells and sensitizes TRAIL-mediated cell death. Mol Cell Biochem 382, 217–224 (2013). https://doi.org/10.1007/s11010-013-1737-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11010-013-1737-0