
Abstract
The Id1 protein is critical for endothelial cell angiogenesis, and this function is particularly relevant to cancer development, cardiovascular disease, and wound healing. We hypothesized that Id1 enhanced migration and tubulogenesis by controlling the expression and function of p53. In this study, we examined cell migration following Id1 overexpression and silencing endothelial cells. The results showed that overexpression of Id1 enhanced cell migration and increased beta1-integrin expression, but inhibition of beta1-integrin blocked motility even in clones overexpressing Id1, suggesting that Id1 regulated motility through beta1-integrin. Further analysis revealed that p53, whose expression and distribution is regulated by Id1, was critical for cell migration, and may be involved in regulating the expression of beta1-integrin. Inhibiting p53 function using PFT-α, a functional inhibitor of p53, increased the expression of beta1-integrin and promoted cell migration even in Id1-silencing endothelial cells, demonstrating that the Id1 knockdowns induced inhibition of endothelial cell migration and the expression of beta1-integrin were controlled by p53. In addition, Id1–p53 pathway regulated the cytoskeleton formation and tubulogenesis. These results demonstrate that Id1-induced beta1-integrin expression in endothelial cells and the function of Id1 in cell migration and tubulogenesis are dependent on p53.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The inhibitor of differentiation 1 protein (Id1) is a transcription factor with helix-loop-helix (HLH) structure that is essential for DNA binding and transcriptional regulation. Activation of Id1 is critical for cell growth and differentiation in many different cell types and tissues [1], and Id1 is a well-established therapeutic target to suppress proliferation and angiogenesis in cancerous and ischemic tissues [2]. Moreover, selective inhibition of Id1 may reduce endothelial progenitor cells proliferation and so may be a novel strategy to inhibit tumor angiogenesis and growth [3, 4].
Angiogenesis is a complex organizational event, and the earliest stages involve endothelial cell attachment to the extracellular matrix (ECM) and ECM proteolysis. Subsequently, endothelial cells coordinate the spatially selective formation and degradation of ECM and expression of adhesion receptors of the integrin family to cross-tissue boundaries [5]. Angiogenesis also requires coordinated endothelial proliferation and migration [6, 7]. Moreover, reorganization of the F-actin cytoskeleton and cell–matrix adhesion play crucial roles in endothelial cell (EC) adhesion, migration, and morphogenesis during angiogenesis [8]. The expression of Id1 has been implicated in the migration of mature endothelial cells [2]. Although the effect of Id1 on the migration has been explored, the molecular mechanism and signaling pathways controlled by the Id1 protein have not been clearly defined.
Accumulating evidences suggested that the beta1-integrin play a role in angiogenesis and the angiogenesis process may be controlled by beta1-integrin through the regulation of cell adhesion, migration, survival, and morphogenesis [9, 10]. In addition, beta1-integrin is also critical for vessel formation through establishing endothelial cell polarity [11] and vascular patterning during embryonic development [12]. Furthermore, angiogenesis inhibitors, such as Maspin [13] affect cell adhesion and cytoskeleton reorganization through integrin signaling. Although these results show that beta1-integrin is critical for endothelial cells angiogenesis, the detailed mechanisms by which beta1-integrin functions in the process of Id1-mediated angiogenesis remain unclear.
In this study, we examined the role of Id1 overexpression in endothelial cell migration and contraction. We investigated Id1-regulated beta1-integrin expression by inhibiting the activity of p53 protein. These results demonstrated that activation of the p53-hypoxia inducible factor (HIF) pathway was required for these endothelial cell migration and contraction.
Materials and methods
Materials and cell culture
Human umbilical vascular endothelial cells (HUVECs) lines were obtained from American Type Culture Collection (ATCC, Manassas, VA), and maintained in Roswell Park Memorial Institute (RPMI 1640) supplemented with 10% fetal bovine serum (FBS) at 37°C in 5% CO2. Antibodies against Id1; p53; β-actin, and beta1-integrin (for beta1-integrin blocking antibody, and beta1-integrin-FITC for Flow Cytometer) were purchased from Santa Cruz. The beta1-integrin antibody used in western blotting was purchased from Abcam. The HIF-1α antibody was purchased from Wuhan Boster Biological Technology (China).
Generation of Id1-t and si-Id1
The HUVECs-pIRES (con-t) and HUVECs-Id1 transfectants (Id1-t) were established as described previously [14]. Stable transfectants were selected with 800 μg/ml geneticin (G418, Sigma-Aldrich, St. Louis, MO, 63112, USA) after transfection. The surviving colonies were picked after 3 weeks.
A stable Id1 siRNA vector was generated using a previously described method [15]. Briefly, the nucleotide sequence coding a short hairpin RNA targeting the Id1 mRNA were cloned into the pSuppressor-Retro vector to generate the siRNA expression vector. The sequences of the si-Id1 nucleotide sequence as follow: si-Id1-F, TCG AGG CTG TTA CTC ACG CCT CAA GGA AGC TTG CTT GAG GCG TGA GTA ACA GCC TTT TT; si-Id1-R, CTA GAA AAA GGC TGT TAC TCA CGC CTC AAG CAA GCT TCC TTG AGG CGT GAG TAA CAG CC. The control vector was generated using the same procedures as the siRNA vector, and the short hairpin RNA sequence was replaced with non-sense sequences that are not homologous to the human genome. The sequences of the control siRNA [15] were: si-con-F, TCG AGC GTA TTG CCT AGC ATT ACG TGA TGC TTG ACG TAA TGC TAG GCA ATA CGC TTT TT; si-con-R, CTA GAA AAA GCG TAT TGC CTA GCA TTS CGT CAA GCT TCA CGT AAT GCT AGG CAA TAC GC. The resulting vectors were then transfected into HUVECs using Lipofectin (Life Technologies) in accordance with the manufacturer’s instructions. Positive si-Id1 and si-con clones were then selected in Neomycin (200 μg/ml) and stable transfectants were isolated after 3 weeks drug selection.
Cell migration
To measure cell migration, confluent monolayers of HUVECs were “wounded” by scraping with a thin 10 μl pipette tip across the monolayer to produce lesions with constant diameter. The cells were washed three times with PBS to remove loose cells, and then cells were cultured in RIPA-1640 media with 2% FBS. Images were obtained using phase contrast microscopy (at ×40 mag.) at the time of initial wounding, and then again at 24 h post-wounding. Semiquantitative measurements were made of control and treated wounds with ImageTool software.
Collagen gel contraction
For collagen gel contraction assay [16], 600 μl cushions of 2 mg/ml collagen I was prepared in 24-well plates. Next, HUVECs (5 × 105 per well) were seeded in serum-free medium. And the medium contains 1 μg/ml beta1-integrin antibody or 10 μg/ml PFT-α follow the experiment needs. After all cells had attached, the medium was removed and cells were overlaid with fresh medium containing 600 μl of 2 mg/ml collagen I, which subsequently polymerized. Gel diameters were monitored with a dissecting microscope 2 days later.
Western blotting
Cells were washed three times with ice-cold PBS, and lysates were prepared in buffer containing 1% NP-40, 0.5% deoxycholate, 0.1% SDS, 10 mM Tris–HCl, 0.2 mM PMSF, and protease. The lysate was boiled for 5 min and then the supernatants were transferred to fresh microcentrifuge tubes and stored at −20°C. Equal amounts of protein were loaded onto each lane of a 12% SDS-PAGE gel. After blotting, polyvinylidene difluoride membranes were blocked for 2 h in 5% milk powder in TBS-Tween at room temperature. Membranes were then incubated with primary antibodies overnight at 4°C. The binding of secondary HRP antibodies was visualized by ECL or ECL Plus. Quantification of the expression of protein though the optical density by Quantity One (Bio-Rad, California, USA).
Flow cytometry
All labeling steps were performed on ice in 5 ml polypropylene round-bottom FACS tubes. The cell suspension was centrifuged at 1000 rpm for 5 min, and the cells resuspended in blocking buffer (49 ml RIPA-1640; 1 g bovine serum albumin, BSA; 1 ml FBS) on ice for 15 min. The cell suspension was transfer (2.0 × 105 cells per sample) into the FACS tubes for the controls required to set up the flow cytometer. Some cell pellets were treated with beta1-integrin antibody (2 μg/tube). The pellets were dispersed, mixed well, and incubated on ice for 1 h with agitation every 20 min to prevent cell sedimentation and clumping. The cells were washed by adding 3 ml of wash buffer (100 ml RIPA-1640, 1 g BSA) to the tubes, and centrifuged at 1000 rpm for 5 min at 4°C. The supernatant was aspirated, and the cell pellet dispersed by flicking the tube. These steps were repeated. After a final wash, the cells were resuspended in the sample tube to a final concentration of 3 × 106 cells/ml in wash buffer.
F-actin staining
HUVECs were washed three times with PBS, plated on glass cover slips, and fixed for 30 min in PBS with 3.7% formaldehyde. Fixed cells were then treated with 0.1% Triton X-100 for 10 min. The F-actin was directly stained with rhodamine-phalloidin (1:100) for 30 min after blocking in 1% BSA for 30 min at room temperature. Cell nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI, Beyotime Institute of Biotechnology, Shanghai, China) in the mounting medium. Cells were washed three times with PBS and photographed using a confocal microscope.
Immunofluorescence
HUVECs that had been plated the previous day on a slide glass for 24 h. After washing three times with PBS, the cells were fixed with 4% paraformaldehyde for 30 min and then permeabilized with 0.25% Triton X-100 in PBS for 10 min at room temperature. The cells were incubated in a blocking solution of 5% BSA in PBS for 1 h and stained with anti-p53 antibody (1:100 dilution in 5% BSA) overnight at 4°C, followed by the addition of Cy3-conjugated goat anti-rabbit secondary antibody (1:100 dilution in 5% BSA; Santa Cruz, California, USA). Nuclei were stained with DAPI. Stained cells were visualized by fluorescence microscopy (Olympus BX81; Olympus, Tokyo, Japan) after the sections were washed in PBS thrice for at least 20 min. To determine the cytoplasmic/nuclear ratio of p53, the integral optical density (IOD) of the fluorescence in the nuclear and cytoplasmic areas in each cell were averaged to calculate the cytoplasmic-to-nuclear ratio of immunofluorescence intensity, using the Image Pro Plus (Media Cybernetics, Washington, DC, USA). In selected experiments, the background fluorescence was determined by selecting the IOD in areas without cells. In each experiment, a minimum of 100 cells were randomly selected and examined.
In vitro tube formation
The in vitro tube formation assay was performed using a matrigel basement membrane matrix (BD Biosciences). Matrigel, kept on ice, was placed at 200 μl/well in 24-well culture plates. The plates were then incubated at 37°C for 30 min to allow matrigel to solidify. HUVECs were seeded at 104 cells per well on the top of the solidified matrigel, and the plate was incubated at 37°C for 24 h. Tube formation on matrigel was observed and analyzed under the microscope. The degree of angiogenesis was measured by multiplying the number of branch points by the total number of branches.
Statistical analysis
Data are given either as mean ± SD or as proportions. In cases of significant differences between the groups, multiple comparisons between groups were made by one way ANOVA. Comparisons between two groups were made by independent t test.
Results
Id1 is required for endothelial cell migration and contraction
To investigate whether Id1 is involved in regulating endothelial cell migration, we constructed two transfectant clones either overexpressing Id1 (Id1-t) or with Id1 knockdown using small interfering RNA (si-Id1). Results showed that Id1 transfected clones (Id1-t) expressed 150% of control Id1 protein, while Id1 knockdowns (si-Id1) expressed just 47% of control Id1 expression (Fig. 1a). We then quantified the cells migration in Id1-t, con-t, si-Id1, and si-con HUVECs. Compared to control cells (con-t), we observed a 20% increase in vitro wound healing in the Id1 overexpressing clones relative to control cells (con-t), but an almost 20% decrease in migration in Id1 knockdown clones, compared with the control siRNA clones (si-con) (Fig. 1b, c). In addition, endothelial cell proliferation was minimal in culture with RIPA-1640 media with 2% FBS during 24 h (data not shown). Thus, ectopic Id1 expression led to increased migration of HUVEC, while suppression of endogenous Id1 expression reduced migration.

Overexpression of Id1 promotes cell migration in HUVECs. a The expression of Id1 protein in HUVECs with Id1-t, con-t, si-con, and si-Id1 was examined by western blotting. The data are relative to β-actin in the same lane. Each lane was loaded with 30 μg proteins. b Cells migration assays though cell wound healing. c Quantification of migration for the clones Id1-t, con-t, si-con, and si-Id1. **P < 0.01, *P < 0.05 for Id1-t versus with con-t; ## P < 0.01, # P < 0.05 for si-Id1 versus with si-con
Id1 controls the expression of beta1-integrin
To evaluate whether Id1-induced cell migration depends on beta1-integrin, we first examined the effect of varying Id1 on the expression of beta1-integrin. Overexpression of Id1 led to the increased expression of beta1-integrin in HUVECs, while Id1 knockdown almost abolished the beta1-integrin expression (Fig. 2a, c). Recent studies demonstrated that the expression and function of beta1-integrin was regulated by transcriptional mechanisms dependent on HIF-1α [17]. Therefore, we examined the expression of HIF-1α in HUVECs overexpressing Id1 (Fig. 2a, b). In all, Id1 overexpression contributed to enhanced expression of HIF-1 in addition to beta1-integrin.

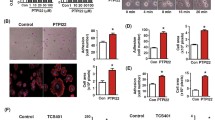
Overexpression of Id1 promotes expression of HIF-1 and beta1-integrin (ITGB1) in HUVECs. a Western blotting analysis of HIF-1 and beta1-integrin expression. b, c The densitometry values for HIF-1 and beta1-integrin normalized to β actin. **P < 0.01 for Id1-t versus with con-t; ## P < 0.01 for si-Id1 versus with si-con. d Quantification of migration of cells after treated or untreated with 1 μg/ml ITGB1 antibody. e Analysis of collagen gel contraction estimated as the area of the gels at the 48 h after treatment with 1 μg/ml ITGB1 antibody. *P < 0.05 for con-t versus with Id1-t; # P < 0.05 for Id1 cells treatment with anti-ITGB1 versus Id1 cells
To investigate the role of beta1-integrin in Id1-induced cell migration, we stimulated cells with a functional inhibitory beta1-integrin antibody. This antibody inhibited cell migration by 25%. Although Id1 overexpression increased EC migration, beta1-integrin antibody inhibited migration of ECs overexpressing Id1 on collagen I (Fig. 2d). It is well known that the force for cell migration can be generated by cellular contraction [18]. In addition, cell contraction was abrogated by beta1-integrin antibody even in HUVECs overexpressing Id1 (Fig. 2e). These results suggested that the enhanced cell migration and contraction observed in Id1-overexpressing HUVECs was beta1 integrin-dependent. Thus, Id1 expression controlled beta1-integrin expression and beta1-integrin mediated the concomitant increase in cell migration and contractility.
Id1 controls the expression and distribution of p53
Mutated p53 proteins can enhance beta1-integrin trafficking [19], so we assessed the role of p53 in the processes of Id1-mediated expression of beta1-integrin. The endothelial cell clones overexpressing ectopic Id1 showed almost no detectable p53 expression compared to the control transfectant clones. In contrast, Id1 knockdown clones demonstrated a fourfold greater p53 expression compared with si-con transfectant clones (Fig. 3c). In normal con-t and Id1-t cells, weak nuclear and cytoplasmic p53 staining was observed, while robust p53 protein expression was localized to the nucleus, with little or no cytoplasmic expression, in si-Id1 clones (Fig. 3a, b).

Overexpression of Id1 inhibits p53 nuclear translocation and expression in HUVECs. a HUVECs stably expressing Id1-t, con-t, si-con, and si-Id1 harvested for detecting p53 localization by indirect immunofluorescence. b Quantification of distribution of p53 after stably expressing Id1-t, con-t, si-con, and si-Id1. c Western blotting analysis of p53 expression and the densitometry values for p53 normalized to β-actin. **P < 0.01 for si-Id1 versus with si-con
p53 controls the expression of beta1-integrin
The expression and function of p53 can be regulated by Id1 and the beta1-integrin expression was regulated by Id1, strongly suggesting that p53 might also regulate beta1-integrin expression and function. To ascertain whether p53 played a role in regulating the expression of beta1-integrin and cell migration, we inhibited p53 function using PFT-α, a functional inhibitor of p53. We first examined the expression and distribution of p53 in cells pretreated for 24 h with 10 μg/ml PFT-α. After 24 h, the expression and the distribution of p53 showed no obvious change (data not shown). We then asked whether p53 could control the expression of HIF-1 and beta1-integrin. Indeed, Id1 knockdown severely abolished HIF-1α and beta1-integrin expression, while PFT-α led to the increased expression of beta1-integrin in Id1 knockdown clones (Fig. 4a, b, c). In addition, flow cytometry analysis indicated that the cells treated with PFT-α exhibited an induction of beta1-integrin protein compared with untreated Id1 knockdown clones (Fig. 4d).

The p53 inhibitor PFT-α increases the expression of beta1-integrin and promoted cell migration even in si-Id1 (knockdown) clones. a Western blotting analysis of HIF-1 and beta1-integrin (ITGB1) expression and the densitometry values (b, c) for HIF-1 and ITGB1 normalized to β-actin. d Flow cytometry detected the effect of PFT-α-regulated ITGB1 expression. e Quantification of migration of cells pretreated with 10 μg/ml PFT-α or untreated. f Analysis of collagen gel contraction estimated as the area of the gels at the 48 h after treatment with 10 μg/ml PFT-α. *P < 0.05 for si-Id1 versus with si-Id1 + PFT-α
Subsequently, the role of p53 in Id1-induced cell migration was examined by stimulating cells with PFT-α. This 10 μg/ml PFT-α increased cell migration by 16%. Although Id1 knockdown decreased ECs migration, PFT-α increased migration of Id1 knockdown ECs migration (Fig. 4e). In addition, cell contraction was increased by PFT-α even in Id1 knockdown cells (Fig. 4f). These results suggested that the decreased cell migration and contraction observed in Id1 knockdown HUVECs was p53 dependent. Thus, Id1–p53 pathway mediated the concomitant cell migration and contractility.
Id1–p53 pathway regulates the cytoskeleton formation and tubulogenesis
From unpublished data, we have evidences that Id1 is vital for the disassembly of actin filaments in the cytoskeleton, and also actin polymerization connected with angiogenesis [20], so we examined if p53 might also regulate the organization of the actin cytoskeleton and tubulogenesis. Preincubation with 10 μg/ml PFT-α for 24 h induced more cytoplasmic F-actin filaments and actin stress fibers than those observed in untreated si-Id1 clones and appeared to significantly increase the peripheral rim of F-actin (Fig. 5a).

The p53 inhibitor PFT-α regulates Id1-dependent cytoskeleton rearrangement and tubulogenesis. a The cytoskeleton for clones si-con and si-Id1 were pretreated for 24 h with PFT-α (10 μg/ml) or untreated. b Treatment of si-con and si-Id1 cells with PFT-α promoted capillary tube formation. c Quantification of the total capillary tube length per field was shown in b. *P < 0.05 for si-Id1 + PFT-α versus si-Id1
Cell migration and cytoskeleton formation play critical roles in the angiogenic process. To determine whether Id1–p53 pathway was required for HUVECs tubulogenesis, we cultured si-Id1, and si-con clones on Matrigel in RIPA-1640 media with 10% FBS, and with or without 10 μg/ml PFT-α 24 h later. Compared with si-con cells, there was almost no tube formation in si-Id1 clones, while there was an increase of tube formation in PFT-α-treated groups (Fig. 5b, c).
Discussion
The purpose of this study was to investigate the effects of the Id1 protein in tubulogenesis through endothelial cell migration and contraction as well as the underlying molecular signaling pathways. In a series of studies, we demonstrated that Id1 promoted endothelial cell migration by increasing beta1-integrin expression, and that the regulation of beta1-integrin by Id1 was controlled by p53. Previous studies focused mainly on the role of Id1 in cell proliferation and survival, while this study demonstrated that Id1 expression enhanced cell migration and contraction while suppression of Id1 expression resulted in reduced migration. Furthermore, Id1 expression was correlated with beta1-integrin expression; however, the enhancement of migration was blocked by a beta1-integrin antibody, indicating that the effects of Id1 were mediated by beta1-integrin. Inhibition of p53 enhanced migration, even in Id1 knockdown clones, suggesting that the motility induced by Id1 was actually controlled by p53.
Although numerous studies have demonstrated the role of Id1 in angiogenesis, few studies examined the molecular signaling pathways involved. Id1 has been implicated in tumor cell migration and invasion [21], our study directly demonstrated that Id1 overexpression promoted endothelial cells migration. Furthermore, Id1 enhanced cell migration through an HIF-1/beta1 integrin-dependent pathway. Beta1-integrin is necessary for cell migration on two-dimensional substrates or even in 3D microenvironments [22]. The essential role of beta1-integrin in migration of HUVECs was demonstrated by the inhibitory effect of a beta1-integrin antibody on cell migration and contraction on clones overexpressing Id1. Thus, inhibition of integrin signaling repressed the migration of endothelial cell even in cells with high Id1 expression.
Cell contraction is necessary for cell migration and beta1-integrin is critical in this process by serving as a link between the ECM and the cytoskeleton. Collagen contraction and crosslinking that facilitate the transfer of contractile forces at focal adhesions is integrin-dependent [23]. This tension triggers the integrin switch that couples cytoskeletal and extracellular matrix mechanics for adhesion-dependent motility [24]. In addition, HIF-1 has been shown to increase cell contraction, beta1-integrin expression [17] and angiogenesis [25]. Id1 enhanced cell contraction through an HIF-1/beta1 integrin-dependent pathway.
The tumor suppressor protein p53 is a critical transcription factor for cell proliferation, survival, and cell cycle progression. Recent studies demonstrated that p53 can also affect other cellular functions, such as cell migration [26]. Furthermore, a loss-of-function p53 mutation contributed to migration directionality in cancer cells and metastasis by enhancing integrin trafficking [19]. Given that p53 can downregulate the expression of HIF-1 [27, 28]; we speculate that p53 exerts its detrimental effects, at least in part, by down-regulating beta1-integrin. Taken together, these research results suggest that p53 may be required for Id1-mediated regulation of beta1-integrin expression. This hypothesis is further supported by a series of experiments in this study.
Pifithrin-α is a reversible inhibitor of p53 and so generally considered to be useful compound to elucidate p53-mediated events in a variety of cells [29]. If Id1-induced expression of beta1-integrin in endothelial cells was mediated by upregulating p53 signaling, then PFT-α should upregulate beta1-integrin, even when Id1 was downregulated. We focused first on the influence of the PFT-α on the expression and nuclear translocation of p53 in endothelial cells. Results indicated that PFT-α does not affect the expression and nuclear translocation of p53, just as 10–100 μM PFT-α did not affect the expression or nuclear translocation of p53 in cancer cells [30]. Inhibition of p53 signaling promoted the migration of endothelial cells, and also induced the expression of HIF-1 and beta1-integrin.
Although our results clearly indicated that Id1 regulated the expression and function of beta1-integrin through p53, it remains to be determined whether Id1 binds directly to the beta1-integrin gene promoter. Furthermore, cells were grown on 2-dimensional (2-D) culture plates, so studies on 3-dimensional (3-D) ECM scaffolds or in vivo studies employing transgenic animal models are warranted.
Our findings suggested that Id1-induced migration is associated with corresponding changes in beta1-integrin, and also demonstrated that p53 suppressed Id1-induced beta1-integrin expression. These results may be of value in the management of tubulogenesis related diseases like heart attack and stroke. Another implication of our study is that Id1-induced beta1-integrin expression and migration may be connect with the HIF-1 pathway. This study may lay the foundation for new rational therapeutic strategies to control tubulogenesis related cardiovascular diseases.
Abbreviations
- Id1:
-
Inhibitors of differentiation
- HLH:
-
Helix-loop-helix
- ECM:
-
Extracellular matrix
- HIF:
-
Hypoxia inducible factor
- HUVEC:
-
Human umbilical vascular endothelial cells
- RPMI 1640:
-
Roswell Park Memorial Institute 1640
- FBS:
-
Fetal bovine serum
- BSA:
-
Bovine serum albumin
- DAPI:
-
6-Diamidino-2-phenylindole
References
Norton JD (2000) ID helix-loop-helix proteins in cell growth, differentiation and tumorigenesis. J Cell Sci 113(Pt 22):3897–3905
Nishiyama K, Takaji K, Kataoka K, Kurihara Y, Yoshimura M, Kato A, Ogawa H, Kurihara H (2005) Id1 gene transfer confers angiogenic property on fully differentiated endothelial cells and contributes to therapeutic angiogenesis. Circulation 112:2840–2850
Gao D, Nolan DJ, Mellick AS, Bambino K, McDonnell K, Mittal V (2008) Endothelial progenitor cells control the angiogenic switch in mouse lung metastasis. Science 319:195–198
Wang H, Yu Y, Guo RW, Shi YK, Song MB, Chen JF, Yu SY, Yin YG, Gao P, Huang L (2010) Inhibitor of DNA binding-1 promotes the migration and proliferation of endothelial progenitor cells in vitro. Mol Cell Biochem 335:19–27
Cheresh DA, Stupack DG (2008) Regulation of angiogenesis: apoptotic cues from the ECM. Oncogene 27:6285–6298
Adams RH, Alitalo K (2007) Molecular regulation of angiogenesis and lymphangiogenesis. Nat Rev Mol Cell Biol 8:464–478
Riedel K, Koellensperger E, Ryssel H, Riedel F, Goessler UR, Germann G, Kremer T (2008) Abrogation of TGF-beta by antisense oligonucleotides modulates expression of VEGF and increases angiogenic potential in isolated fibroblasts from radiated skin. Int J Mol Med 22:473–480
Sun X, Li C, Zhuang C, Gilmore WC, Cobos E, Tao Y, Dai Z (2009) Abl interactor 1 regulates Src-Id1-matrix metalloproteinase 9 axis and is required for invadopodia formation, extracellular matrix degradation and tumor growth of human breast cancer cells. Carcinogenesis 30:2109–2116
Mettouchi A, Meneguzzi G (2006) Distinct roles of beta1 integrins during angiogenesis. Eur J Cell Biol 85:243–247
Carlson TR, Hu H, Braren R, Kim YH, Wang RA (2008) Cell-autonomous requirement for beta1 integrin in endothelial cell adhesion, migration and survival during angiogenesis in mice. Development 135:2193–2202
Zovein AC, Luque A, Turlo KA, Hofmann JJ, Yee KM, Becker MS, Fassler R, Mellman I, Lane TF, Iruela-Arispe ML (2010) Beta1 integrin establishes endothelial cell polarity and arteriolar lumen formation via a Par3-dependent mechanism. Dev Cell 18:39–51
Lei L, Liu D, Huang Y, Jovin I, Shai SY, Kyriakides T, Ross RS, Giordano FJ (2008) Endothelial expression of beta1 integrin is required for embryonic vascular patterning and postnatal vascular remodeling. Mol Cell Biol 28:794–802
Qin L, Zhang M (2010) Maspin regulates endothelial cell adhesion and migration through an integrin signaling pathway. J Biol Chem 285:32360–32369
Chen L, Qiu J, Yang C, Yang X, Chen X, Jiang J, Luo X (2009) Identification of a novel estrogen receptor beta1 binding partner, inhibitor of differentiation-1, and role of ERbeta1 in human breast cancer cells. Cancer Lett 278:210–219
Ling MT, Lau TC, Zhou C, Chua CW, Kwok WK, Wang Q, Wang X, Wong YC (2005) Overexpression of Id-1 in prostate cancer cells promotes angiogenesis through the activation of vascular endothelial growth factor (VEGF). Carcinogenesis 26:1668–1676
Hoang MV, Whelan MC, Senger DR (2004) Rho activity critically and selectively regulates endothelial cell organization during angiogenesis. Proc Natl Acad Sci USA 101:1874–1879
Keely S, Glover LE, MacManus CF, Campbell EL, Scully MM, Furuta GT, Colgan SP (2009) Selective induction of integrin beta1 by hypoxia-inducible factor: implications for wound healing. FASEB J 23:1338–1346
Sheetz MP (1994) Cell migration by graded attachment to substrates and contraction. Semin Cell Biol 5:149–155
Muller PA, Caswell PT, Doyle B, Iwanicki MP, Tan EH, Karim S, Lukashchuk N, Gillespie DA, Ludwig RL, Gosselin P, Cromer A, Brugge JS, Sansom OJ, Norman JC, Vousden KH (2009) Mutant p53 drives invasion by promoting integrin recycling. Cell 139:1327–1341
Pyatibratov MG, Tolkatchev D, Plamondon J, Xu P, Ni F, Kostyukova AS (2010) Binding of human angiogenin inhibits actin polymerization. Arch Biochem Biophys 495:74–81
Bhattacharya R, Kowalski J, Larson AR, Brock M, Alani RM (2010) Id1 promotes tumor cell migration in nonsmall cell lung cancers. J Oncol 2010:856105
Caswell PT, Chan M, Lindsay AJ, McCaffrey MW, Boettiger D, Norman JC (2008) Rab-coupling protein coordinates recycling of alpha5beta1 integrin and EGFR1 to promote cell migration in 3D microenvironments. J Cell Biol 183:143–155
Levental KR, Yu H, Kass L, Lakins JN, Egeblad M, Erler JT, Fong SF, Csiszar K, Giaccia A, Weninger W, Yamauchi M, Gasser DL, Weaver VM (2009) Matrix crosslinking forces tumor progression by enhancing integrin signaling. Cell 139:891–906
Friedland JC, Lee MH, Boettiger D (2009) Mechanically activated integrin switch controls alpha5beta1 function. Science 323:642–644
Huang L, Zhang Z, Zhang S, Ren J, Zhang R, Zeng H, Li Q, Wu G (2011) Inhibitory action of Celastrol on hypoxia-mediated angiogenesis and metastasis via the HIF-1alpha pathway. Int J Mol Med 27:407–415
Gadea G, de Toledo M, Anguille C, Roux P (2007) Loss of p53 promotes RhoA-ROCK-dependent cell migration and invasion in 3D matrices. J Cell Biol 178:23–30
Yamakuchi M, Lotterman CD, Bao C, Hruban RH, Karim B, Mendell JT, Huso D, Lowenstein CJ (2010) P53-induced microRNA-107 inhibits HIF-1 and tumor angiogenesis. Proc Natl Acad Sci USA 107:6334–6339
Sano M, Minamino T, Toko H, Miyauchi H, Orimo M, Qin Y, Akazawa H, Tateno K, Kayama Y, Harada M, Shimizu I, Asahara T, Hamada H, Tomita S, Molkentin JD, Zou Y, Komuro I (2007) p53-induced inhibition of Hif-1 causes cardiac dysfunction during pressure overload. Nature 446:444–448
Carlisi D, Vassallo B, Lauricella M, Emanuele S, D’Anneo A, Di Leonardo E, Di Fazio P, Vento R, Tesoriere G (2008) Histone deacetylase inhibitors induce in human hepatoma HepG2 cells acetylation of p53 and histones in correlation with apoptotic effects. Int J Oncol 32:177–184
Murphy PJ, Galigniana MD, Morishima Y, Harrell JM, Kwok RP, Ljungman M, Pratt WB (2004) Pifithrin-alpha inhibits p53 signaling after interaction of the tumor suppressor protein with hsp90 and its nuclear translocation. J Biol Chem 279:30195–30201
Acknowledgments
This research program was supported by grants from the National Natural Science Foundation of China (30970721) and the Fundamental Research Funds for the Central Universities (CDJXS10 23 22 10).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Qiu, J., Wang, G., Hu, J. et al. Id1-induced inhibition of p53 facilitates endothelial cell migration and tube formation by regulating the expression of beta1-integrin. Mol Cell Biochem 357, 125–133 (2011). https://doi.org/10.1007/s11010-011-0882-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11010-011-0882-6