
Abstract
Context
For many organisms, responses to climate change (CC) will be affected by land-use and land-cover changes (LULCC). However, the extent to which LULCC is concurrently considered in climate change vulnerability assessments (CCVAs) is unclear.
Objectives
We identify trends in inclusion of LULCC and CC in vulnerability assessments of species and the direction and magnitude of their combined effect on biodiversity. Further, we examine the effect size of LULCC and CC in driving changes in “currencies” of response to CC, such as distribution, abundance and survival.
Methods
We conducted a systematic literature review of articles published in the last 30 years that focused on CCVA and accounted for impacts of both CC and LULCC.
Results
Across 116 studies, 34% assumed CC and LULCC would act additively, while 66% allowed for interactive effects. The majority of CCVAs reported similar effect sizes for CC and LULCC, although they affected different CCVA currencies. Only 14% of the studies showed larger effects of CC than of LULCC. Another 14% showed larger effects of LULCC than CC, specifically for dispersal, population viability, and reproduction, which tend to be strongly affected by fragmentation and disturbance. Although most studies found that LULCC and CC had negative effects on species currencies, in some cases effects were neutral or even positive.
Conclusions
CCVAs that incorporate LULCC provided a better account of drivers of vulnerability, and highlight aspects of drivers that are generally more amenable to on-the-ground management intervention than CCVAs that focus on CC alone.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Climate change (CC) is a major driver of biodiversity change, alongside concurrent impacts of land-use and land-cover change (LULCC), invasive species, pollution and overexploitation (IPBES 2019). CC and LULCC shape niches and geographical distributions of species, their persistence and extinction probability, and the processes through which species interact with their environment and with each other (Pacifici et al. 2017). Climate change vulnerability assessments (CCVAs) typically define vulnerability as a function of exposure, sensitivity, and adaptive capacity (Williams et al. 2008; Foden and Young 2016). CCVAs in general have been applied to many different areas (mountain ranges, biomes, countries, states, conservation areas, etc.), sectors (agriculture, water, energy, etc.), while varying in the level of biological and ecological detail considered (ecosystems, communities, species). For CCVAs focused on aspects of biodiversity, exposure is defined as the amount of environmental change the species could experience; sensitivity as the species’ response when exposed to such change; and adaptive capacity as the ability of the species to adjust or cope with the impacts, for example through adaptive evolution, migration, or behavioral change (Watson et al. 2013).
Although CC is expected to become a major driver of species’ endangerment, currently the most threat to species arises from LULCC (IPBES 2019). LULCC will likely continue to be a dominant driver of vulnerability into the future (Marques et al. 2019). LULCC can affect vulnerability independently of CC, but it can also interact with CC to exacerbate or ameliorate endangerment (Mantyka-Pringle et al. 2015). Indeed, a recent global analysis predicts that LULCC alone by 2070 could result in a loss of 8% of terrestrial vertebrates (Powers and Jetz 2019), and together with CC could lead to a cumulative loss of about 38% of terrestrial vertebrates (Newbold 2018). As a result, CCVAs that ignore current and anticipated LULCC risk mischaracterizing the vulnerability of species (Foden et al. 2013; Pacifici et al. 2015). LULCC can be incorporated into CCVAs under the assumption that it interacts with CC additively (CC + LULCC) or interactively (CC * LULCC). For example, CC and LULCC each directly shape species’ physiological tolerances in terrestrial (Chuine 2010) and aquatic environments (Thackerey et al. 2016), phenology (Blois et al. 2013), interspecific interactions, movement patterns (Tucker et al. 2018), resource availability (Walther et al. 2002), and energy requirements (Gallagher et al. 2017). Climate change and LULCC also interact to affect vulnerability. LULCC determines the incidence, type, and effectiveness of macro- and microclimatic refugia (Morelli et al. 2012, 2020; Stralberg et al. 2018), alters landscape connectivity important for climate-driven range shifts (Meier et al. 2012), and facilitates spread of invasive species that benefit from climate change.
Several factors may hinder the ease of inclusion of LULCC in CCVA: lack of data, difficulty in quantifying the manner in which CC and LULCC interact, and uncertainty (Sirami et al. 2017). Incorporating CC and LULCC requires data on both current and anticipated future states of both drivers, which are not often available (especially for LULCC). While the climate modeling community has self-organized to produce widely-agreed upon sets of scenarios (e.g., the Intergovernmental Program on Climate Change’s Representative Concentration Pathways), data sources for current and projected LULCC are more fragmented and less standardized (but see Schulp et al. 2019).
Here we report on a systematic literature review to assess biodiversity-focused CCVAs for their inclusion/integration of LULCC and CC, and evaluate how these factors affect the responses of biodiversity. CCVAs use many types of “responses” to characterize vulnerability (geographical distributions, survival rates, and other species traits), which, following a previous global review (Pacifici et al. 2015), we call “currencies.” First, we assess trends in the use of various currencies in CCVAs that include both CC and LULCC. Second, we evaluate the frequency and manner in which LULCC is incorporated into historical and future (predictive) CCVAs. Then, we assess representation of taxonomic groups and type of impact of LULCC on currencies (positive/negative/neutral). Finally, we use this information to propose ways forward for enhanced integration of CC and LULCC in CCVA.
Methods
Trends in CCVA currencies
CCVAs measure or estimate responses to a variety of currencies, including: 1) absolute changes in distribution or population size, 2) probability of extinction, or 3) indices or relative measurements of changes in distribution, population, or extinction risk (Pacifici et al. 2015). These currencies can be assessed by CCVA frameworks that rely on different methodological approaches (Pacifici et al. 2015; Wheatley et al. 2017). Many assessments (especially of large groups of species for which data availability is low) utilize a scoring-system based on species’ auto-ecological and life‐history traits expected to mediate climate impacts (Foden et al. 2013). Other assessments use statistical or mechanistic models to project future geographical distributions of species or population dynamics. Both approaches can be combined in “hybrid” assessments that score traits and use model projections for multidimensional assessment of the different currencies (Pacifici et al. 2015; Smith et al. 2016).
We performed a literature search to determine how are currencies utilized in CCVA assessments. Our systematic review examined whether or not 1) LULCC was assumed to act independently or interact with CC to determine vulnerability; 2) LULCC was assumed to be static or dynamic through time; 3) LULCC and CC affected the same or different currencies; and 4) LULCC and CC had the same or opposing effects on overall vulnerability, and their relative magnitude (positive or negative). We conducted the search on 27.03.2018 following recommendations for conducting systematic reviews (Gurevitch et al. 2018). Journal articles were located using Web of Science (WoS) and PubMed. In this initial step, no time restrictions were implemented for the search. For WoS, terms were searched under “Topic” (title, abstract, key terms, key terms plus). For PubMed terms were searched under “title, abstract, key terms”. To define search terms that could correspond to currencies in CCVA, we did a screening of relevant texts in ecology, conservation biology, animal behavior, etc. (See Appendix I for a brief search on the importance of CCVA and the details on the search method). Acknowledging that some search terms or concepts do not necessarily correspond to a single currency (e.g., “species distribution” and “dispersal”), a quick scoping search using WoS and PubMed only for those concepts showed how often these cited are in the same papers or used interchangeably. We retrieved 86 search terms, which are found in Appendix I (Table I.2.). To simplify the search, we defined three search strings, i.e., sets of key terms (Table I.3), and the resulting records are displayed in Table I.4 in Appendix I (note that the search described in Sect. 2.1. is not included). We opted not to review the references within the papers that we retrieved from the search engines.
Papers were then filtered to retain studies directly related to CCVA. First, we excluded studies that (i) focused on vulnerability of human populations or human infrastructure and activities, (ii) examined climate impacts occurring outside the period 1900–2100, (iii) were reviews or meta-analyses, (iv) examined only effects of LULCC or of CC, or how CC affects LULCC but not biodiversity, (v) were context-specific or at fine spatial scales (i.e., very local studies or in very small study areas likely not representative of species’ entire distributions), (vi) or did not clearly identify the studied types of impacts in the title or abstract. A total of 131 papers were retained (Appendix I, Table I.3). A coding procedure was then implemented to extract relevant information from these papers. With this step, we systematically retrieved information on (i) taxonomic group, (ii) currency(ies) (distribution, abundance, survival, dispersal, composition, interactions, diversity, and reproduction; Pacifici et al. 2015), (iii) whether the impact was observed (historical) or projected (future), (iv) the aspect(s) of LULCC studied, (v) the aspect(s) of CC studied, (vi) the manner in which LULCC and CC are assumed to affect vulnerability (additively/independently: CC + LULCC, versus interactively: CC*LULCC), (vii) methods for including LULCC and CC in CCVA, (viii) direction of the impact by CC and LULCC (negative when the combined drivers result in a decrease in the currency connoting increased vulnerability, positive when the combined drivers result in an increase in the currency connoting less vulnerability, mixed when the drivers have positive and negative effects together, or neutralizing when the positive or negative effects of the drivers counteract each other), and (ix) effect sizes and main findings.
Results
Trends in CCVA currencies
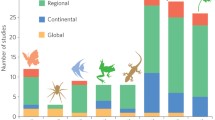
Our systematic review identified 116 studies that formally included both LULCC and CC into assessments of climate change vulnerability (Fig. 1). Of these, 66% allowed for interactions between LULCC and CC (CC * LULCC: n = 77), with the remainder (n = 39) assuming they acted independently of one another to affect vulnerability (CC + LULCC). Throughout the timeframe of the studies we analyzed, we found that potential interactions between LULCC and CC (CC * LULCC) were considered more after ~ 2009 than before (33% before and 67% thereafter 2009). Sirami et al. (2017) also found a similar increase around 2007. LULCC was assumed to be static (i.e., unchanging with time) in 82% of the studies (n = 95) and dynamic in the rest. Most studies were attempting to predict future vulnerability, except for studies examining abundance, which tended to focus on historical trends. Plants were the most commonly studied taxonomic group, followed by birds then mammals. The majority of studies reported negative impacts on currencies (increased vulnerability; 67%, n = 78), but for some currencies, such as survival and community composition, mixed impacts were frequently observed. CCVAs that incorporated LULCC used a diversity of inferential methods (e.g., traits, correlative and process-based modeling), but interpreting differences among the prevalence of methods was not straightforward because some of the currencies could only be estimated using specific inferential methodology (e.g., through correlative or mechanistic and process-based modeling).

Frequency of studies on CCVA currencies that included CC and LULCC additively or interactively (CC + LULCC and CC * LULCC), historical (empirical observations and modeled trends) and prospective analyses (model projections), per taxonomic group and with reported positive, negative, mixed and neutral impacts. (Graphic symbols designed by Dr. Dovi Kacev)
Distribution: Distributional changes were reported for all taxonomic groups, with a slightly higher number of studies focusing on plants, followed by birds and mammals. The reported distributional changes were mostly predictive; only 15% were historical (n = 17). The majority of the studies that reported distributional changes allowed for interactions between CC and LULCC (CC * LULCC; n = 55). A few studies reported negative effects of either CC or LULCC on dispersal (n = 7), mostly assessed using distribution models of terrestrial plants and mammals. An almost even number of studies that reported on dispersal ability used CC + LULCC or CC * LULCC (Fig. 1). The most frequently described distributional changes were range contractions and range shifts.
Population parameters: Most studies using abundance (including population growth rates) as a currency reported negative effects of exposure of CC and LULCC. These studies covered a wide range of taxonomic groups (Fig. 1). Contrary to studies focusing on distributional changes, population changes were more commonly the focus of historical CCVAs rather than predictive CCVAs. An even larger proportion of the studies using abundance currencies included CC * LULCC. We find that 15% (n = 17) of the studies reported shifts in abundance. A few studies reported decreases in survival rates (n = 3), most of which were on birds and ascertained using either observations or models that assumed CC + LULCC.
Extinction probability: Surprisingly, none of the studies reported changes in extinction probability, likely because of the short time span of historical studies and the uncertainty with which future extinctions can be predicted. Nonetheless, some studies mentioned that range contractions coupled with distribution shifts towards higher latitudes and elevations tend to result in high local extinctions.
Other ecological currencies: Eight percent (n = 9) of the papers considered the impact of CC and LULCC on other ecological processes, such as species interactions, community composition, and diversity. These studies focused on plants, fungi, insects, and fish, and included LULCC * CC.
CC and LULCC in CCVA
Most studies did not explicitly report the aspect of climate which was assumed to most affect vulnerability (e.g., drought, extreme storms, heat waves, rising temperatures, etc.), although studies included a range of variables that could reflect or could be correlated with specific climatic stressors (Fig. 2a). Similarly, several aspects of LULCC were utilized as “drivers” of potential change (Fig. 2b). Many proxies were used to describe land use, the most common one being a metric of land use change, followed by static descriptors of land cover type or of land use type or habitat quality. More infrequent proxies of LULCC were descriptors of fragmentation, connectivity and deforestation, or of human disturbance and population density.

CC and LULCC and CC phenomena addressed by the selected studies: a CC and b LULCC (SLR—sea level rise; N/A—not applicable i.e. when the study did not attribute the vulnerability to a specific phenomenon). Most studies do not make an attribution of changes in CCVA currencies with one or more dimensions of LULCC or CC. For LULCC, deforestation, agriculture and fragmentation were the most common topics, with reforestation being only mentioned in the recent publications. For CC, the dominant dimension was global warming, and only very few recent studies addressed other dimensions
Inferential methodologies were varied (Fig. 3), with a greater variety of methods used to assess distribution and abundance as compared to other currencies. Species distribution modeling (SDM) was the most common method used, as geographic distribution changes were the most common types of studies, but also regression analyses were also common. Some studies proposed novel integration of methodologies, such as for example SDM and population viability analysis (Tian et al. 2014), metapopulation evaluation (García-Valdés et al. 2015) or demographic analysis (Hunt et al. 2017), SDM and graph theory (Giannini et al. 2015), environmental niche factor analysis and stochastic demographic models (Fordham et al. 2012), and even machine learning methods for assessing vulnerability (Heubes et al. 2013).

Methodologies used to integrate LULCC into CCVA for each of the currencies. SDM—Species distribution models (includes Maxent, logistic regression, etc.), ENFA—Environmental Niche Factor Analysis, GAMs—Generalized Additive Models, PVA—Population Viability Analysis, GLMM—Generlized Linear Mixed Models, SVMs—Support Vector Machines. When two methods are reported, the study integrated them
Direction of impact and effect size
Most studies reported negative impacts (enhanced vulnerability; Fig. 4); it was not possible to determine whether they only assessed negative impacts or whether they contained a reporting bias against positive impacts. Negative impacts occurred irrespective of the taxonomic group. Several studies (19%) modeled or conceptualized a hierarchical interaction between LULCC and CC to assess whether LULCC had an effect on the ability of species to occupy potential ranges predicted on the basis of climate. The majority of studies (66%; 77 out of 116) reported negative effects of LULCC, which frequently exacerbated the negative effects of CC (n = 43, CC* LULCC). LULCC and CC tended to either affect different currencies or operated at different spatial and temporal scales, with CC being longer-term and broader-scale than LULCC. In 34% of cases LULCC and CC affected currencies in opposing directions, which led to a zero-sum net effect of LULCC and CC for 12% of studies. Finally, we found that a small fraction of studies (14%; 16 out of 116 studies) reported that CC had a markedly greater effect than LULCC when examining distribution or abundance, while a similar fraction of studies reported the opposite with a greater effect from LULCC than CC for both distribution and dispersal. Inclusion of LULCC became unimportant in two particular cases: at high latitudes where much LULCC has yet to occur (Kéry et al. 2006) and in extremely built-up places where additional LULCC was predicted to be restricted (Bombin and Reed 2016) or where most of it had already occurred in the past (Argent et al. 2018).

CC and LULCC impacts and effect sizes on the different currencies. Effect sizes for studies by currency (N at the top for each currency), Plus sign indicates a positive effect, negative sign indicates a negative effect, plus/minus indicate drivers in different directions and 0 indicates no net effect. Greyed out symbols indicate no studies. (Graphic symbols designed by Dr. Dovi Kacev)
Discussion
We set out to find the degree to which LULCC acted independently or interactively with CC in climate change vulnerability assessments, how each affects different currencies, and whether or not the inclusion of LULCC exacerbates or opposes effects of CC. Two-thirds of the studies allowed for interactive effects (CC*LULCC), with proportionately more studies doing so since 2009 that before (Oliver and Morecroft 2014; Sirami et al. 2017), perhaps because of the growing awareness of LULCC as the strongest driver of current biodiversity loss (IPBES 2019). Most studies (66%) found (or assumed) only negative effects of LULCC on vulnerability, although in 12% of studies their opposing impacts effectively neutralized one another. The number of studies in which the effects of CC overshadowed the effects of LULCC or vice versa was the same (14% in both cases). LULCC and CC were generally assumed to affect different currencies, and the diversity of currencies considered broadened through time. All together, we see these trends as promising signs because they likely reflect the real manner in which CC and LULCC affect biodiversity (i.e., interactively and in a multi-faceted manner). Nonetheless, our results highlight areas of potential improvement; the diversity of currencies used and the various ways in which LULCC was assumed to interact (or not) with CC suggests, that we have yet to develop a comprehensive understanding of how LULCC and CC act together to affect facets of vulnerability.
LULCC and CC effects on vulnerability
CC and LULCC are widely acknowledged to influence vulnerability, so accounting for both in the same assessment will likely lead to a higher estimated vulnerability. We found this indeed to be the case for most studies. We also found that most studies showed notable effects of both drivers, with no systematic differences in the magnitude or sign of effect. However, despite the fact that including LULCC enhanced estimated vulnerability, we did find that including LULCC could have mixed effects on CCVA currencies, i.e., positive, negative, or neutralizing effects, irrespective of taxonomic group. For example, one of the most commonly reported positive synergies was an increase in range size due to expansion of suitable habitat conditions arising from both CC and LULCC (we note that a “positive” impact at the species level is not necessarily a positive effect at other levels, if range shifts cause community reorganization or enhance invasiveness).
Despite the general consensus that CC and LULCC make species more vulnerable, a few CCVAs did not find any clear negative or positive effects. Similarly, negative effects on one currency could diminish positive effects in another. For example, for population parameter currencies, one of the studies we reviewed showed that due to rising temperatures, the mosquito Aedes notoscriptus may experience increased predation but, concurrently, shorter life cycles, which could therefore buffer higher mortality rates (Hunt et al. 2017). Surprisingly, only a small fraction (14%) of the studies reported a much stronger effect of CC, and an equivalent fraction (14%) reported a stronger effect of LULCC. Across studies, LULCC tended to be relatively unimportant in two cases: in landscapes where anthropogenic LULCC has yet to occur (Kéry et al. 2006) and in extremely built-up places where future LULCC is predicted to be restricted or have little additional impact (Argent et al. 2018).
We also found that CCVAs tend to incorporate LULCC such that it operates at different spatial and temporal scales than CC. This likely follows from the theoretical understanding that, in general, LULCC operates at local, shorter time scales and CC on broader and longer scales (Soberón 2007). In some cases, these differences in scale are reflected in the hierarchical nature of the framework used to assess impacts in which LULCC is used as a “filter” to assess whether species can, for instance, colonize newly climatically suitable habitat. Despite a general consensus that effects of CC operate at broader scales (Soberón 2007), comparisons are somewhat compromised when the spatial and temporal resolutions of land use and climate data are mismatched, which we found to be common in the studies we reviewed. Similarly, LULCC datasets with high thematic resolution (e.g., number of land cover classes) and broad temporal coverage are rare, making it more difficult to include LULCC in CCVAs. Relatedly, many potentially important aspects of LULCC are not reflected in available data sets, including habitat-relevant land cover classes, land use (versus simply cover), density of roads, power lines, etc.
LULCC and CC effects on currencies
The compounding effect of LULCC on impacts predicted from CC alone arises because LULCC and CC tend to affect different currencies (Thuiller et al. 2004), which occurs because they operate at different spatial and temporal scales. Most studies we reviewed report CC’s influence on distribution while LULCC tended to be assumed to affect dispersal, reproductive output, and population viability. If true, these differences could arise because changes in geographical distribution usually require longer time periods to respond to external drivers, making CC more suitable “predictor” of distributional shifts and thus more informative (Taheri et al. 2021). On the other hand, LULCC tends to have more immediate effects on survival—species loss with deforestation for instance, and dispersal—effects of barriers like roads, and thus LULCC affects aspects usually considered when assessing extinction probability. It is also possible that the immediate effects of CC are less straightforward to observe (but see Blois et al. 2013, and the emerging field of conservation physiology e.g., Thackerey et al. 2016). While some CCVA currencies may be more informative than others, our results echo conclusions by Pacifici et al. (2015), who also found that the use of more informative currencies is still relatively rare among CCVAs. For example, the focus on geographic distribution is warranted for understanding the potential future impacts of climate change on species' ranges (Taheri et al. 2021). However, realization of projected changes in geographical distribution is dependent on assumptions about dispersal, establishment, successful reproduction, and biotic interactions, which are less commonly assessed in CCVAs – but those that do indeed tend to frame LULCC and CC in a multi-tiered approach. We found studies that incorporated LULCC were more likely use the latter set of currencies. However, these currencies are difficult to assess because data on demographic parameters are less available than on broad-scale distribution.
We found that there is not only a diversity of methods used to include both LULCC and CC on the studies of the impacts of these global change drivers on CCVA currencies, but also that the type of proxy for LULCC (i.e., the “agent” of LULCC that was expected to affect vulnerability) was extremely diverse. This may be a strength—LULCC can manifest itself in multiple dimensions and is expected to affect many different aspects of ecological systems. However, this diversity of formulations of LULCC proxies can also be a weakness as it makes comparison between studies difficult and may reflect lack of knowledge about the most important drivers of changes induced by LULCC.
Moving forward on CCVAs
We believe that the first step necessary to improve the implementation CCVA is to better understand how currencies relate to each aspect of CC and LULCC. There are several initiatives for collecting data that could support better assessments of CCVA currencies and improve our understanding of the interacting effects of LULCC and CC (Fig. 5; Hudson et al. 2014). For reliable CCVAs we need long-term, large-area studies, but we generally have short-term, small-area data. For example, data relevant to biotic interactions (i.e., prey resources, competitive regimes, phenological timing, refugia, etc.) require detailed studies, often at local scales, focusing on a few selected target species, typically require a long timeframe and large teams to collect data. Movement data (i.e., activity time, movement patterns, phenology, prevailing winds, connectivity, introductions) also require detailed studies, which are costly, time consuming and logistically demanding, and therefore usually limited to a single species in a single region at a time. While tracking studies have a long tradition in ecology, systematic collection of such data is only now taking place, and will allow determining general patterns about activity times (Gaynor et al. 2018) and movement patterns (Tucker et al. 2018). Likewise, initiatives like the IUCN's Global Invasive Species database (http://www.iucngisd.org/gisd), assess movement by certain classes of organisms. Alternatively, coupled model/empirical efforts may help bridge this gap. For example, Gallagher et al. (2017) illustrate how to derive an “energy landscape” that serves as a proxy for prey resources which could in turn be modeled as a function of land cover and climate. Similarly, advances in understanding and measuring landscape thermal signatures may provide future data on landscape characteristics as a proxy for refugia (Meerdink et al. 2019). Other efforts such as GEOBON Essential Biodiversity Variables (https://geobon.org/ebvs/what-are-ebvs/) can serve to monitor many of the currencies over larger areas, in a systematic and consistent way. Finally, initiatives like NEON (https://www.neonscience.org/) or LTER (https://lternet.edu/) in the US are now working on operationalizing a workflow to link repeat satellite or airborne imagery with time-series plot data to track biodiversity and habitat conditions. Thus, there are many efforts underway that will allow for improved CCVAs relatively soon. Within the plethora of data sources and types that are available or soon will be available, we foresee two challenges. First, not all data are collected or available at the spatial or temporal scales most relevant to CCVAs or at which management action occurs. Second, data often reflect land cover, but not land use. A step forward could be to develop a common platform to address issues related to data availability; define a common set of modeling conventions for predicting future LULCC; document and database responses of species to CC (like the PREDICTS database for LULCC; Hudson et al. 2014); and share knowledge to increase efficiency and effectiveness.

Example of currently available initiatives that could be used in or improved by CCVA (see appendix II for a list of the references included in this figure, hyperlinks to data, extent, resolution and data type). This process is not without challenges relating to data collection and scenarios to be modelled in CCVA for the different currencies. (Graphic symbols designed by Dr. Dovi Kacev)
For inferring the effects of CC and LULCC on biodiversity it is necessary to rely on models that relate individual species or community characteristics (e.g., species richness or turnover) to particular environmental factors, which can be projected into future scenarios of LULCC and CC. These models, however, face many challenges, which relate to the quality and availability of (species and environmental) data and to the modelling approach used (Faurby and Araújo, 2018). Modelling changes in distributions due to climate change is founded on the assumption that species occupy a specific niche, which for many species, include land cover characteristics (e.g., Hunt et al. 2017; Santos et al. 2017). Hence, appropriate assessment of changes in the amount of area where niche requirements are met needs to include both CC and LULCC. However, the predictors used in species distribution modelling are often represented by simplified data that lack ecological realism (Leitão and Santos, 2019). For example, many LULCC products are categorical in nature (e.g., agriculture/forest) so do not differentiate within categories (e.g., kind of crop, primary vs. secondary forest, type or intensity of use, etc.). Nevertheless, animals and plants respond to differences in the same general type of habitat, and we lack information on LULCC that can be linked to the relevant biological processes rather than anthropocentric perspectives of land cover classes (but see Moeslund et al. 2019). We also lack longitudinal data going back in time to reconstruct LULCC over more than a couple of decades. Projects like HYDE (Goldewijk et al. 2017) aim at reconstructing past LULCC with historical and anthropological accounts and can serve as data sources that span longer time periods, albeit at coarse resolution.
Our second suggestion for advancing the implementation of CCVAs is to further include interactions, especially dynamic interactions, between LULCC and CC. Many CCVA assume a static environmental context where vulnerability is assessed at a single point in time with data or model projections from one or more periods, within which no change is assumed (or mean values are used—effectively assuming no change). In reality, the environment is constantly changing and so vulnerability estimates should, as well. However, so far, process-based biodiversity models are mostly only possible for functional types (Hickler et al. 2012). Likewise, the integration of process-based modeling related to dispersal, ecophysiology (Thackerey et al. 2016), population dynamics (Zurell et al. 2016), biotic interactions (van der Putten et al. 2010) and ecological traits (Brown et al. 2014), although still uncommon approaches, constitute promising avenues for the improvement of predictions on the effects of LULCC and CC on biodiversity. Some of the studies we reviewed reported or accounted for interactions between LULCC and CC effects. Increasingly, regional and global circulation models incorporate feedbacks between LULCC and climate (e.g., the Advanced Canopy-Atmosphere-Soil Algorithm ACASA; Marras et al. 2008). Land use type affects albedo and sensible and latent heat fluxes, which in turn could alter land cover or use. Despite advances, not all relevant feedbacks between LULCC and CC have been incorporated into regional and global circulation models, as for example effects of deforestation and fire on water cycling and changes in temperature and albedo, effects of wetland removal on evapotranspiration and cooling, or effects of agricultural irrigation or urbanization on changes in albedo and cooling (Massad et al. 2018). At the global scale, integrated assessment models have been used to model development scenarios and simulate future LULCC (Doelman et al. 2018). In such models, the main drivers of LULCC are land use for agriculture, bioenergy, and carbon storage (Leclère et al. 2020). At the regional level several general LULCC models also allow for dynamic land cover types. Regional models include a set of rules to attribute which land use type will more likely be found in a grid cell at a given point in time. Many of these models have been used to assess future scenarios for energy and commodity development, as well as encroachment on ecologically valuable land (e.g., Verstegen et al. 2019). We do note opportunities for incorporating thematically-focused LULCC models into regional and global circulation models (e.g., Sleeter et al. 2019). For example, urban growth models could be better integrated into regional climate models to account for urban heat island effects and related effects of development on albedo and latent heat flux.
Third, we suggest we need to better include aspects of LULCC that are currently missing from CCVAs, such as land use intensity and ecological restoration. LULCC models need to better reflect disturbance (natural or anthropogenic), land use intensity, abandonment, and rejuvenation or restoration. Although most LULCC models do not explicitly account for abandonment or land use intensity, or for encroachment (legal or illegal) on natural ecosystems, some models do allow reversion to earlier states (e.g., forest becomes cropland which then later becomes forest again). However, intentional restoration is also typically ignored, likely because it would fit into the sensitivity and adaptive capacity component of a CCVA or be recommended in response to the outcome of a CCVA. Restoration should always be carefully considered to mediate or reduce LULCC and CC effects in CCVA studies (Strassburg et al. 2019). However, given the structural and functional losses found in restored ecosystems compared to undisturbed ecosystems (Moreno-Mateos et al. 2017), restoration should be incorporated into CCVAs since restored ecosystems may translate to reduced exposure and enhance adaptive capacity.
Finally, there is also a need to better assess all components of vulnerability, i.e., exposure, sensitivity and adaptive capacity. We find that the direct inclusion of LULCC on CCVA provides a more comprehensive picture of threats to species (as suggested by the wider diversity of currencies used by studies incorporating LULCC; Fig. 4). CC is typically assumed to affect exposure, whereas sensitivity and adaptive capacity to CC are presumed to be “inherent” to the species. In contrast, LULCC can interact with sensitivity and adaptive capacity (as well as affect exposure). Studies that use CC and LULCC in combination bring a broader understanding of the factors making species vulnerable. Surprisingly, only a small fraction of studies (12%) reported that CC is a stronger driver of change than LULCC. Incorporating LULCC in CCVA brings additional understanding as CC and LULCC mostly have compound effects, as different CCVA currencies are impacted by CC and LULCC. Although joint effects of LULCC and CC can be additive, multiplicative, or ameliorative, we found that most CCVAs concluded greater negative effects on biodiversity when LULCC was included. This is particularly important as it suggests that there will be a systematic bias towards underestimating these effects when LULCC is excluded. Additionally, LULCC and CC operate at different scales in space and time, which can help defining better conservation actions and management plans alongside with rethinking or optimizing the way CCVAs are currently implemented (Fig. 5).
Many CCVAs to date result from government-led initiatives and some of the academic literature has no explicit link to ongoing government programs or even to NGO lead initiatives (for example the IUCN; Foden and Young 2016). A better integration of these three types of institutional ways of addressing CCVAs may help streamline their content, comparability and the synthesis we aim to achieve about how species respond to and are impacted by global change drivers. For example, some locations are better at implementing CCVAs than others, e.g. California has just finished its fourth CCVA and in many European countries similar initiatives are ongoing at different paces. LULCC and CC are also hard to measure and model together, thus more and better data and methodological advances are necessary which can again benefit from a better integration across agencies that implement or study biological vulnerability. Further, part of this work will necessitate better understanding of biophysical feedbacks between CC and LULCC, and how these feedbacks affect biodiversity and responses of species to LULCC and CC (Williams and Newbold 2019), and herein we outline ongoing efforts and provide a roadmap and recommendations for improvement of CCVAs in the future.
Change history
16 October 2021
A Correction to this paper has been published: https://doi.org/10.1007/s10980-021-01332-5
References
Argent DG, Kimmel WG, Derek G (2018) Changes in the status of native brook trout on Laurel Hill, Southwestern Pennsylvania. NorthEas Nat 25:1–20. https://doi.org/10.1656/045.025.0101
Blois JL, Zarnetske PL, Fitzpatrick MC, Finnegan S (2013) Climate change and the past, present, and future of biotic interactions. Science 341:499–504
Bombin A, Reed LK (2016) The changing biodiversity of Alabama Drosophila: important impacts of seasonal variation, urbanization, and invasive species. Ecol Evol 6:7057–7069
Brown AM, Warton DI, Andrew NR, Binns M, Cassis G, Gibb H (2014) The fourth-corner solution – using predictive models to understand how species traits interact with the environment. Meth Ecol Evol 5:344–352
Chuine I (2010) Why does phenology drive species distribution? Trans Roy Soc London B 365:3149–3160
Doelman JC, Stehfest E, Tabeau A, van Meijl H, Lassaletta L, Gernaat DEHJ, Hermans K, Harmsen M, Daioglou V, Biemans H, van der Sluis S, van Vuuren DP (2018) Exploring SSP land-use dynamics using the IMAGE model: regional and gridded scenarios of land-use change and land-based climate change mitigation. Glob Env Ch 48:119–135
Faurby S, Araújo MB (2018) Anthropogenic range contraction bias species climate change forecasts. Nature Clim Ch 8:252–256
Foden WB, Young BE (2016) IUCN SSC Guidelines for Assessing Species’ Vulnerability to Climate Change. Version 1.0. Occasional Paper of the IUCN Species Survival Commission No. 59. Cambridge, UK and Gland, Switzerland: IUCN Species Survival Commission. x+114pp
Foden WB, Butchart SHM, Stuart SN, Vié J-C, Akçakaya R, Angulo A, DeVaniter LD, Gutsche A, Turak E, Cao L, Donner SD, Katariya V, Bernard R, Holland RA, Hughes AF, O’Hanlon SE, Garnett ST, Şekercioğlu ÇH, Mace GM (2013) Identifying the world’s most climate change vulnerable species: a systematic trait-based assessment of all birds, amphibians and corals. PlosONE 8:e65427. https://doi.org/10.1371/journal.pone.0065427
Fordham DA, Watts MJ, Delean S, Brook BW, Heard LMB, Bull CM (2012) Managed relocation as an adaptation strategy for mitigating climate change threats to the persistence of an endangered lizard. Glob Ch Biol 18:2743–2755
Gallagher A, Creel S, Wilson RP, Cooke SJ (2017) Energy landscapes and the landscape of fear. Tr Ecol Evol 32:88–96
García-Valdés R, Svenning J-C, Zavala MA, Purves DW, Araújo MB (2015) Evaluating the combined effects of climate and land-use change on tree species distribution. J Appl Ecol 52:902–912
Gaynor KM, Hojnowski CE, Carter NH, Brashares JS (2018) The influence of human disturbance on wildlife nocturnality. Science 360:1232–1235
Gianinni TC, Tambosi LR, Acosta AL, Jaffé R, Saraiva AM, Imperatriz-Fonseca VL, Metzger JP (2015) Safeguarding ecosystem services: a methodological framework to buffer the joint effect of habitat configuration and climate change. PLoS ONE 10(6):e0129225. https://doi.org/10.1371/journal.pone.0129225
Goldewijk KK, Beusen A, Doelman J, Stehfast E (2017) Anthropogenic land use estimates for the Holocene—HYDE 3.2. Earth Syst Sci Data 9:927–953
Gurevitch J, Koricheva J, Nakagawa S, Stewart G (2018) Meta-analysis and the science of research synthesis. Nature 555:175–182
Heubes J, Schmidt M, Stuch B, Marquez JRG, Wittig R, Zizka G, Thiombiano A, Sinsin B, Schaldach R, Hahn K (2013) The projected impact of climate and land use change on plant diversity: an example from West Africa. J Arid Env 96:48–54
Hickler T, Vohland K, Feehan J, Miller PA, Smith B, Costa L, Giesecke T, Fronzek S, Carter TR, Cramer W, Kühn I, Sykes MT (2012) Projecting the future distribution of European potential natural vegetation zones with a generalized, tree species-based dynamic vegetation model. Glob Ecol Biogeog 21:50–63
Hudson LN, Newbold T, Contu S, Hill SLL, Lysenko I, De Palma A, Phillips HRP, Senior RA, Bennett DJ, Booth H, Choimos A, Correia DLP, Day J et al (2014) The PREDICTS database: a global database of how local terrestrial biodiversity responds to human impacts. Ecol Evol 4:4701–4735
Hunt SK, Galatowitsch ML, McIntosh AR (2017) Interactive effects of land use, temperature, and predators determine native and invasive mosquito distributions. Freshw Biol 62:1564–1577
IPBES (2019) Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. In: Díaz S, Settele J, Brondízio ES, Ngo HT, Guèze M, Agard J, Arneth A, Balvanera P, Brauman KA, Butchart SHM, Chan KMA, Garibaldi LA, Ichii K, Liu J, Subramanian SM, Midgley GF, Miloslavich P, Molnár Z, Obura D, Pfaff A, Polasky S, Purvis A, Razzaque J, Reyers B, Chowdhury R, Shin YJ, Visseren-Hamakers IJ, Willis KJ, Zayas CN (eds) IPBES secretariat, Bonn, Germany. 56 pp. https://doi.org/10.5281/zenodo.3553579
Kéry M, Madsen J, Lebreton J-D (2006) Survival of Svalbard pink-footed geese Anser brachyrhynchus in relation to winter climate, density and land-use. J an Ecol 75:1172–1181
Leclère D, Obersteiner M, Barret M, BUtchard SHM, Chaudhary A, De Palma A, DeClerk FAJ, Di Marco M, Doelman JC, et al (2020) Bending the curve of terrestrial biodiversity needs an integrated strategy. Nature 585:551–556. https://doi.org/10.1038/s41586-020-2705-y
Leitão PJ, Santos MJ (2019) Improving models of species ecological niches: a remote sensing overview. Front Ecol Evol 7:9. https://doi.org/10.3389/fevo.2019.00009
Mahmood R, Pielke RA Sr, Hubbard KG, Niyogi D, Dirmeyer PA, McAlpine C, Carleton AM, Hale R, Gameda S, Beltrán-Przekurat A, Baker B, McNider R, Legates DR, Shepherd M, Du J, Blanken PD, Frauenfeld OW, Nair US, Fall S (2014) Land cover changes and their biogeophysical effects on climate. Int J Clim 34:929–953
Mantyka-Pringle CS, Martin TG, Rhodes JR (2012) Interactions between climate and habitat loss effects on biodiversity: a systematic review and meta-analysis. Glob Ch Biol 18:1239–1252
Mantyka-Pringle CS, Visconti P, Di Marco M, Martin TG, Rondinini C, Rhodes JR (2015) Climate change modifies risk of global biodiversity loss due to land-cover change. Biol Conserv 187:103–111
Marques A, Martins IS, Kastner T, Plutzar C, Theurl MC, Eisenmenger N, Huijbregts MAJ, Wood R, Stadler K, Bruckner M, Canelas J, Hilbers JP, Tukker A, Erb K, Pereira HM (2019) Increasing impacts of land-use on biodiversity and carbon-sequestration driven by population and economic growth. Nature Ecol Evol 3:628–637
Marras S, Spano DEI, Sirca CB, Duce P, Snyder RL, Pyles RD, Paw UKT (2008) Advanced-Canopy-Atmosphere-Soil Algorithm (ACASA model) for estimating mass and energy fluxes. Ital J Agron 3:793–794
Massad RS, Lathière J, Strada S, Perrin M, Personne E, Stéfanon M, Stella P, Szopa S, Noblet-Ducoudré N (2018) Reviews and syntheses: influences of landscape structure and land uses on local to regional climate and air quality. Biogeosc 16:2369–2408
Meerdink S, Roberts D, Hulley G, Gader P, Pisek J, Adamson K, King J, Hook SJ (2019) Plant species’ spectral emissivity and temperature using the hyperspectral thermal emission spectrometer (HyTES) sensor. Rem Sens Environ 224:421–435
Meier ES, Lischke H, Schmatz DR, Zimmermann NE (2012) Climate, competition and connectivity affect future migration and ranges of European trees. Glob Ecol Biogeog 21:164–178
Moeslund JE, Zlinszky A, Ejrnæs R, Brunbjerg AK, Bøcher PK, Svenning JC, Normand S (2019) LIDAR explains diversity of plants fungi lichens and bryophytes across multiple habitats and large geographic extent. BioRxiv. https://doi.org/10.1101/509794
Moreira F, Vaz P, Catry F, Silva JS (2009) Regional variations in wildfire susceptibility of land-cover types in Portugal: implications for landscape management to minimize fire hazard. Int J Wildl Fire 18:563–574
Morelli TL, Smith AB, Kastely CR, Mastroserio I, Moritz C, Beissinger SR (2012) Anthropogenic refugia ameliorate the severe climate-related decline of a montane mammal along its trailing edge. Proceed Roy Soc London B 279:4279–4286
Morelli TL, Barrows CW, Ramirez AR, Cartwright JM, Ackerly DD, Eaves TD, Ebersole JL, Krawchuk MA, Letcher BH, Mahalovich MF, Meigs GW, Michalak JL, Millar CI, Quiñones RM, Stralberg D, Thorne JH (2020) Climate-change refugia: biodiversity in the slow lane. Front Ecol Evol 5:228–234
Moreno-Mateos D, Barbier EB, Jones PC, Jones HP, Aronson J, López-López JA, McCrackin ML, Meli P, Montoya D, Benayas JMR (2017) Anthropogenic ecosystem disturbance and the recovery debt. Nature Comm 8:14163. https://doi.org/10.1038/ncomms14163
Newbold T (2018) Future effects of climate and land-use change on terrestrial vertebrate community diversity under different scenarios. Proceed Roy Soc B. https://doi.org/10.1098/rspb.2018.0792
Oliver TH, Morecroft MD (2014) Interactions between climate change and land use change on biodiversity: attribution problems, risks, and opportunities. Wiley Interd Rev - Clim Ch 5:317–335
Pacifici M, Foden WB, Visconti P, Watson JEM, Butchart SHM, Kovacs KM, Scheffers BR, Hole DG, Martin TG, Akçakaya HR, Corlett RT, Huntley B, Brickford D, Carr JA, Hoffmann AA, Midgley GF, Pearce-Kelly P, Pearson RG, Williams SE, Willis SG, Young B, Rondinini C (2015) Assessing species vulnerability to climate change. Nature Clim Ch 5:215–225
Pacifici M, Visconti P, Butchart SHM, Watson JEM, Cassola FM, Rondinini C (2017) Species’ traits influenced their response to recent climate change. Nature Clim Ch 7:205–208
Powers RP, Jetz W (2019) Global habitat loss and extinction risk of terrestrial vertebrates under future land-use-change scenarios. Nature Clim Ch 9:323–329
Santos MJ, Smith AB, Thorne JH, Moritz C (2017) The relative roles of changing vegetation and climate on elevation range dynamics of small mammals. Clim Change Resp 4:7. https://doi.org/10.1186/s40665-017-0035-6
Schulp CJE, Levers C, Kuemmerle T, Tieskens KF, Verburg PH (2019) Mapping and modelling past and future land use change in Europe’s cultural landscapes. Land Use Pol 8:332–344
Sirami C, Caplat P, Popy S, Clamens A, Arlettaz R, Jiguet F, Brotons L, Martin J-L (2017) Impacts of global change on species distributions: obstacles and solutions to integrate climate and land use. Glob Ecol Biogeog 26:385–394
Sleeter BM, Marvin DC, Cameron DR, Selmants PC, Westerling AL, Kreitler J, Daniel CJ, Liu J, Wilson TS (2019) Effects of 21st-century climate, land use, and disturbances on ecosystem carbon balance in California. Glob Ch Biol 25:3334–3353
Smith AB, Long QG, Albrecht MA (2016) Shifting targets: spatial priorities for ex situ plant conservation depend on interactions between current threats, climate change, and uncertainty. Biodiv Conserv 25:905–922
Soberón J (2007) Grinnellian and Eltonian niches and geographic distributions of species. Ecol Lett 10:1115–1123
Stralberg D, Carrol C, Wisley CB, McKenney DW, Nielsen SE (2018) Macrorefugia for North American trees and songbirds: Climatic limiting factors and multi-scale topographic influences. Glob Ecol Biogeog 27:690–703
Strassburg BBN, Beyer HL, Crouzeilles R, Iribarrem A, Barros F, de Siqueira MF, Sánchez-Tapia A, Balmford A, Sansevero JBB, Brancalion PHS, Broadbent EN, Chazdon RL, Filho AO, Gardner TA, Gordon A, Latawiec A, Loyola R, Metzger JP, Mills M, Possingham HP, Rodrigues RR, Scaramuzza CAM, Scarano FR, Tambosi L, Uriarte M (2019) Strategic approaches to restoring ecosystems can triple conservation gains and halve costs. Nature Ecol Evol 3:62–70
Stuart-Smith RD, Edgar GJ, Barrett NS, Kininmonth SJ, Bates AE (2015) Thermal biases and vulnerability to warming in the world’s marine fauna. Nature 528:88–92
Taheri S, Naimi B, Rahbek C, Araújo MB (2021) Improvements in reports of species redistribution under climate change are required. Sci Adv. https://doi.org/10.1126/sciadv.abe1110
Thackerey SJ, Henrys PA, Hemming D, Bell JR, Botham MS, Burthe S, Helaouet P, Johns DG, Jones ID, Leech DI, Mackay EB, Massimino D, Atkinson S, Bacon PJ, Brereton TM, Carvalho L, Clutton-Brock TH, Duck C, Edwards M, Elliott JM, Hall SJG, Harrington R, Pearce-Higgins JW, Høye TT, Kruuk LEB, Pemberton JM, Sparks TH, Thompson PM, White I, Winfield IJ, Wanless- Show S (2016) Phenological sensitivity to climate across taxa and trophic levels. Nature 535:241–245
Thuiller W, Araújo MB, Lavorel S (2004) Do we need land-cover data to model species distributions in Europe? J Biogeog 31:53–361
Tian Y, Wu J, Wang T, Ge J (2014) Climate change and landscape fragmentation jeopardize the population viability of the Siberian tiger (Panthera tigris altaica). Lands Ecol 29:621–637
Tucker MA, Böhning-Gaese K, Fagan WF, Fryxell JM, Van Moorter B, Alberts SC, Ali AH, Allen AM, Attias N, Avgar T, Bartlam-Brooks H, Bayarbaatar B, Belant JL et al (2018) Moving in the Anthropocene: global reductions in terrestrial mammalian movements. Science 359:466–469. https://doi.org/10.1126/science.aam9712
van der Putten WH, Macel M, Visser ME (2010) Predicting species distribution and abundance responses to climate change: why it is essential to include biotic interactions across trophic levels. Phil Transac Roy Soc B: Biol Sci 365:2025–2034
Verstegen JA, van der Laan C, Dekker SC, Faaij APC, Santos MJ (2019) Recent and projected impacts of land use and land cover changes on carbon stocks and biodiversity in East Kalimantan, Indonesia. Ecol Ind 103:563–575
Walther G-R, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:389–395
Watson JEM, Iwamura T, Butt N (2013) Mapping vulnerability and conservation adaptation strategies under climate change. Nature Clim Ch 3:989–994
Wheatley CJ, Beale CM, Bradbury RB, Pearce-Higgins JW, Critchlow R, Thomas CD (2017) Climate change vulnerability for species - assessing the assessments. Glob Ch Biol 23:3704–3715
Williams JJ, Newbold T (2019) Local climatic changes affect biodiversity responses to land use: A review. Divers Distrib. https://doi.org/10.1111/ddi.12999
Williams SE, Shoo LP, Isaac JL, Hoffmann AA, Langham G (2008) Towards an integrated framework for assessing the vulnerability of species to climate change. Plosbiology 6:e325. https://doi.org/10.1371/journal.pbio.0060325
Zurell D, Thuiller W, Pagel J, Cabral JS, Münkemüller T, Gravel D, Dullinger S, Normand S, Schiffers KH, Moore K, Zimmermann NE (2016) Benchmarking novel approaches for modelling species range dynamics. Glob Ch Biol 22:2651–2664
Acknowledgements
MJS was funded by the University Research Priority Program in Global Change and Biodiversity and the Department of Geography at the University of Zurich. ABS was supported by the Alan Graham Fund in Global Change. PJL was funded by the 2015-2016 BiodivERsA COFUND, with the national funder German Federal Ministry of Education and Research (BMBF; project GreenFutureForest, Grant 01LC1610A). NMH was supported by the Carlsberg Foundation and the European Union’s Horizon 2020 research and innovation program under the Marie Sklodowska-Curie grant agreement no. 746334, and acknowledges support from the Danish National Research Foundation for support to the Center for Macroecology, Evolution and Climate (Grant no. DNRF96). DMM was funded by the Spanish Ministry of Economy and Competitiveness through Societal Challenge Program (Grant CGL2015-70452-R) and María de Maeztu excellence accreditation MDM-2017-0714. We would like to thank Dr. Dovi Kacev for the design of the icons used in the paper. This is a contribution on behalf of the IUCN’s Climate Change Specialist Group, Species Survival Commission.
Funding
Open Access funding provided by Universität Zürich.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Maria J. Santos and Adam B. Smith should be considered joint first author; other authors are listed by alphabetical order of the last name.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Santos, M.J., Smith, A.B., Dekker, S.C. et al. The role of land use and land cover change in climate change vulnerability assessments of biodiversity: a systematic review. Landscape Ecol 36, 3367–3382 (2021). https://doi.org/10.1007/s10980-021-01276-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-021-01276-w