
Abstract
Context
Since 2005, unconventional gas development has rapidly altered forests across the Marcellus-Utica shale basin in the central Appalachian region of the eastern United States, an area of high conservation value for biodiversity. Much is still unknown about ecological impacts of associated land cover change.
Objectives
Our goal was to identify threshold responses among bird species and habitat guilds to (1) overall forest loss and fragmentation in affected landscapes, and (2) distance from anthropogenic disturbance, both related and unrelated to shale gas.
Methods
We conducted 2589 bird surveys at 190 sites across this region, and quantified community-level and species-specific thresholds relating to forest cover and distance to anthropogenic disturbance, using Threshold Indicator Taxa Analysis (TITAN).
Results
Forest interior species decreased abruptly in abundance and frequency of occurrence above a threshold of 17.0% overall forest loss, while early successional and synanthropic species increased abruptly above 30.5–36.5% forest loss, respectively. Broad quantile intervals around responses to distance from anthropogenic disturbance suggest these were not sharp threshold responses, but more gradual or linear responses. Among forest interior species evaluated, 48.1% increased in abundance farther from shale gas development, while 55.6% of early successional and synanthropic species decreased.
Conclusions
We found evidence of avian threshold responses to overall forest loss and fragmentation in affected landscapes across the Marcellus-Utica shale region. Our results suggest that efforts to avoid shale gas development in regional core forests—particularly those still retaining ≥ 83% forest cover—can reduce negative effects on area-sensitive, forest interior dependent species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The extraction of natural gas from deep, low-permeability shale formations has increased dramatically since the early 2000s, when the combined use of horizontal drilling and high-volume hydraulic fracturing technologies (i.e., fracking) increased the economic viability of previously untapped shale resources. These advances in ‘unconventional’ drilling practices opened new regions to shale gas development, including the Marcellus-Utica shale region in the eastern United States, which holds one of the largest and most productive shale gas reserves in North America (USEIA 2017, 2020). The Marcellus-Utica region also encompasses large expanses of core forest habitat of high conservation value (Pickering et al. 2002; Anderson et al. 2012). Emerging patterns of land use change due to shale gas development suggest the industry is driving substantial forest disturbance in the central Appalachian region, raising concerns about potential ecological consequences for biodiversity (Kiviat 2013; Souther et al. 2014; Brittingham et al. 2014). The number of reported wells drilled in the Marcellus-Utica region increased by two orders of magnitude from 115 wells in 2005 to 17,888 by 2018 (MCOR 2019). Improving our understanding of the impacts of this relatively recent form of energy development on wildlife and species diversity is key to the development of effective and sustainable land management and conservation strategies.
Energy sector development often occupies a relatively small footprint on landscapes, but results in a disproportionate amount of habitat fragmentation due to the increase in associated linear infrastructure (i.e., roads, transmission lines and pipelines; Bayne and Dale 2011). The resulting proliferation of edge habitats reduces suitability of impacted landscapes for area-sensitive species, as evidenced by declines in edge-avoiding birds following development of seismic lines in boreal forests (Bayne et al. 2005; Machtans 2006), and oil and gas well pads and roadways in sagebrush steppe (Gilbert and Chalfoun 2011; Mutter et al. 2015) and grasslands (Thompson et al. 2015; Nenninger and Koper 2018). Unconventional shale gas development is still relatively new to the eastern USA, and much is still unknown about the biological impacts on forest ecosystems. However, localized studies from the region suggest that forest disturbance associated with Marcellus-Utica shale gas development is fragmenting forests (Langlois et al. 2017), facilitating plant invasions (Barlow et al. 2017), reducing forest canopy volume (Young et al. 2018), and restructuring forest bird communities (Barton et al. 2016; Farwell et al. 2016, 2019).
Theoretical models and a growing body of research support the existence of ecological thresholds (i.e., tipping points or change points), beyond which ecological processes show a rapid, non-linear response to an incremental change in environmental pressures (Lande 1987; Betts et al. 2007; Bestelmeyer et al. 2011; Gutzwiller et al. 2015). The ecological effects of forest loss and fragmentation can lead to thresholds of landscape change beyond which additional losses can have disproportionate effects on biological communities (Fahrig 1999; Luck 2005). In fragmented landscapes, habitat generalists are likely to possess sufficient behavioral plasticity to persist in highly variable forest configurations (Leonard et al. 2008). However, there is evidence that area-sensitive species with narrow habitat requirements often exhibit non-linear threshold responses to changes in forest cover (Betts et al. 2007; Zuckerberg and Porter 2010; Rueda et al. 2013). At the local or patch scale, we might see species-specific thresholds in forest loss or fragmentation beyond which individuals cannot persist (Hanski and Hammond 1995; Radford et al. 2005). At a landscape or regional scale, there may be population thresholds in the proportion of forest habitat to the surrounding matrix at which point recolonizations can no longer compensate for emigrations and local extinctions (Hanski and Hammond 1995; Andren 1996; Boulinier et al. 2001). Failing to detect non-linear responses of species to habitat loss and fragmentation complicates management efforts and can lead to incorrect predictions of how species and communities will respond to continued development (Bestelmeyer 2006). Understanding and anticipating nonlinear threshold responses to landscape change are critical aspects of proactive, threshold-based adaptive management and conservation (Foley et al. 2015; van der Hoek et al. 2015).
Terrestrial birds are useful and effective indicators of habitat change and complex community response to human-modified landscapes (Canterbury et al. 2000; O’Connell et al. 2007). They are relatively easy to survey, and exhibit a wide range of space-use behaviors (Leonard et al. 2008) and habitat associations across multiple spatial scales (Warren et al. 2005; Mitchell et al. 2006). Many birds are also facing regional and global declines (Gaston et al. 2003; IUCN 2019), including several species of conservation concern whose core breeding ranges overlap the Marcellus-Utica shale region. In localized studies from this region, forest interior dependent birds decreased in abundance and richness with increasing proximity to shale gas infrastructure (Barton et al. 2016; Farwell et al. 2019) and increasing forest loss and edge density driven by shale gas development (Farwell et al. 2016). Conversely, some disturbance dependent and synanthropic birds appear to benefit from land cover change resulting from shale gas development, although responses vary by species and type of infrastructure (Barton et al. 2016; Farwell et al. 2016; Langlois 2017). However, these were all local-scale studies, and no published study has evaluated broad trends in avian response to shale gas development across the Marcellus-Utica region. Additionally, previous studies only tested for linear relationships between birds and shale gas development, despite growing interest in possible thresholds of forest disturbance beyond which major changes in species abundances and community assemblages may occur (Gutzwiller et al. 2015). Evidence of such thresholds among forest interior dependent species in the Marcellus-Utica shale region could be used to identify particularly vulnerable species and to inform preventive targets for landscape-level management and conservation (Huggett 2005).
Our objective was to assess whether birds exhibited non-linear threshold responses to anthropogenic forest disturbance across the Marcellus-Utica shale region. First, we tested for threshold responses to overall amount of forest loss and forest edge density in landscapes containing unconventional shale gas development. Second, we assessed potential threshold responses to distance from forest edges created by human development, and evaluated whether distance thresholds differed between forest edges created by shale gas infrastructure compared with other forms of human development, unrelated to shale gas. We used Threshold Indicator Taxa Analysis (TITAN; Baker and King 2010, 2013) to identify change points in bird abundances across gradients of forest cover within 1-km radius landscapes surrounding avian sampling points, and at varying distances from anthropogenic forest edges. We calculated threshold responses for the overall avian community, within habitat guilds, and for individual species within each guild.
Methods
Study area and site selection
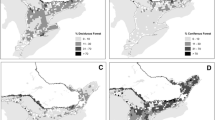
The study encompassed the Marcellus-Utica region (Fig. 1); sampling primarily occurred in areas experiencing shale gas development (hereafter, ‘shale gas’) in Pennsylvania, West Virginia, and eastern Ohio, with limited sampling of adjacent control areas in Maryland and New York. During 2014–2015, we sampled 120 sites impacted by shale gas and other types of human development (‘gas sites’) and 70 sites impacted only by human development unrelated to shale gas (‘non-gas sites’). Most gas sites were centered on a focal gas well pad or cluster of gas well pads; 10 of 120 gas sites did not contain well pads but were centered on other types of related infrastructure (e.g., pipelines, gas compressor stations). Non-gas sites had no unconventional shale gas development within a minimum 1-km radius of any point sampled, 90% had no shale gas within 2 km of any point sampled, and 79% had no shale gas within 3 km. We selected gas sites and non-gas sites from the same Level IV ecoregions (USEPA 2013) and across similar ranges of forest cover using a stratified, semi-random sampling approach (Brennan et al. 2002). Because our aim was to identify threshold responses of birds to development in predominantly forested landscapes (Zuckerberg et al. 2012; van der Hoek et al. 2013), most sites (167 of 190) fell at or above 60% forest cover (\(\overline{x}\) = 79%, range 25–99%).

Map of study region. Sites (n = 190) were surveyed in 5 states: Pennsylvania (n = 108), West Virginia (n = 60), Ohio (n = 17), Maryland (n = 3), and New York (n = 2). Black triangles represent shale gas sites (n = 120), white dots represent non-gas sites (n = 70). Extent of Marcellus shale formation shaded in gray
Land cover and distance metrics
We manually delineated land cover within broad land use categories based on temporally appropriate, 1-m resolution aerial imagery from the National Agriculture Imagery Program (NAIP 2016). NAIP digital orthoimagery is leaf-on and acquired at a 1-m ground sample distance, with a horizontal accuracy of ± 6.0 m (NAIP 2016). Primary land cover classes of interest included forest cover, human development associated with shale gas, and human development unrelated to shale gas. Non-gas development primarily included areas with some built areas but mostly vegetation, such as rural and exurban single-family properties, public parks, and other resource industries (e.g., surface mines, wind energy development). Within both gas and non-gas classes of human development, linear infrastructure (e.g., roads, railroads, powerlines and pipelines) is pervasive and can have a number of distinct negative effects on wildlife (van der Ree et al. 2015; Richardson et al. 2017). Because access roads and pipeline corridors are primary drivers of forest loss and fragmentation associated with shale gas in the Marcellus-Utica region (Farwell et al. 2016; Langlois et al. 2017), we were particularly interested in assessing whether breeding birds exhibit unique responses to linear forms of shale gas infrastructure as opposed to non-linear forms of shale gas development (i.e., drilling sites or ‘well pads’). We were also broadly interested in comparing bird responses to shale gas infrastructure (both linear and non-linear) with similar forms of human development unrelated to shale gas (both linear and non-linear). Thus, we classified human disturbance into four separate classes of development for analysis: areas developed for shale gas drilling sites (‘well pads’), developed areas unrelated to shale gas (‘non-gas development’), linear shale gas infrastructure (‘linear gas’), and linear infrastructure unrelated to shale gas (‘linear non-gas’).
We calculated distances from avian sampling points within forested areas to nearest anthropogenic disturbance, using the ‘near’ function in ArcGIS 10.4 (ESRI 2016). We separated points into those closest to well pads, non-gas development, linear gas, and linear non-gas. We ensured each class of development was the closest anthropogenic edge to points placed in each group. Thus, a linear gas point located 70 m from a pipeline had no other human development within a 70-m radius. For distance threshold analyses, we excluded points for which the closest edge was related to silviculture, since the impacts of forest disturbance due to regenerating timber harvests are expected to differ from the more permanent, maintained types of development that were the focus of this study (Schmiegelow et al. 1997; Marzluff and Ewing 2001; Lichstein et al. 2002).
We converted the vector land cover maps to raster grids, and calculated proportion of forest loss and forest edge density within 1-km radius buffers of each avian sampling point using Fragstats 4.2 (McGarigal et al. 2012). We selected a 1-km radius (314 ha) because this is an area large enough to capture multiple development features on the landscape, while also representing multiple ‘neighborhoods’ of breeding songbird territories (Desrochers et al. 2010) and an area feasibly visited during daily movements by songbirds (Krementz and Powell 2000; Lang et al. 2002). Although there is overlap among 1-km radius buffers, the avian survey data are independent and landscape metrics relate to avian data at each individual point (Zuckerberg et al. 2012; Becker et al. 2015). We did not focus exclusively on forest loss and edge density driven by shale gas but rather on total forest loss and edge density, because our goal was to test for potential thresholds, or ‘tipping points’, of overall forest disturbance within landscapes experiencing shale gas development. Thus, we restricted our forest cover threshold analyses to sampled landscapes containing shale gas (n = 1512).
Avian surveys and detection probability
We conducted avian surveys at 2589 sampling points, distributed among 190 sites across the study region (\(\overline{x}\) = 13.6 points; range = 8–20 points per site). At gas sites, we placed at least 2 sampling points adjacent to forest edges in areas directly disturbed by shale gas, and by other types of human development at non-gas sites. Remaining points were randomly distributed within surrounding forests ≥ 250 m apart to maintain statistical independence (Wood et al. 2006). Forest points were located at varying distances from edges of human development, and ≥ 50 m from natural canopy gaps to help minimize multiple edge effects (Paton 1994). Additionally, natural canopy gaps in our study sites were primarily tree-fall gaps or streams, which generally have fewer negative edge effects on forest birds than anthropogenic disturbances (Forsman et al. 2010).
We counted birds using standard 10-min avian point count surveys (Bibby et al. 1992; Ralph et al. 1995) during 12 May–3 July in 2014 and 15 May–6 July in 2015. Each sample point was surveyed once in either 2014 or 2015 by one of 13 observers experienced in regional bird identification and distance estimation. We conducted surveys between sunrise and 4 h after sunrise, when weather conditions did not interfere with audibility or visibility of birds. We recorded first detections of birds within 5 time intervals (0–2, > 2–4, > 4–6, > 6–8, > 8–10 min) and 4 distance bands (0–25, > 25–50, > 50–75, > 75–100 m), to allow for estimation of detection probability using a combined time removal and distance sampling approach (Sólymos et al. 2013). This approach provides conditional maximum likelihood estimates for the two components of detectability: availability (the probability that a bird will sing) and perceptibility (the probability that a bird will be detected). We included Julian date, time-since-sunrise, and quadratic terms of each as visit-level covariates potentially affecting singing rates (availability) and observer and forest cover as visit-level covariates potentially affecting perceptibility. We included forest cover within 100 m of sampled points as a detection covariate because foliage affects amplitude reduction, with sounds attenuating at shorter distances in forested settings (Huisman and Attenborough 1991; Yip et al. 2017). To calculate forest cover within 100 m of each survey point, we used the same raster land cover data described above. We only analyzed data for species with sufficient observations to run detection probability models; we found detection models converged for species with at least 50 detections. For species with ≥ 50 detections, we fitted 9 removal models and 4 distance sampling models, including intercept-only models (Online Appendix A), using the package ‘detect’ (Sólymos et al. 2016) in program R version 3.4.1 (R Core Team 2017). Using fitted models with lowest Akaike’s information criterion (AIC) values, we calculated custom offsets to account for detection probability and used adjusted abundances in subsequent analyses (Sólymos et al. 2013).
Threshold analysis
We used TITAN (Baker and King 2010, 2013) to analyze adjusted avian abundances, using the ‘TITAN2’ package (Baker et al. 2015) in R version 3.4.1 (R Core Team 2017). TITAN uses a combination of change-point analysis (King and Richardson 2003; Qian et al. 2003) and indicator species analysis (Dufrene and Legendre 1997) to identify abrupt changes in relative abundance and frequency of occurrence of species across an environmental gradient. Indicator species are organisms associated with a specific environmental condition, where a change in the occurrence or abundance of the species signals a change in that condition, and thus may be used as an indicator of the shifting health or integrity of an ecosystem (Carignan and Villard 2002). TITAN evaluates mid-points between values along a continuous environmental gradient as potential thresholds, iteratively partitioning observations into two groups and using change-point analysis to maximize a deviance reduction statistic that compares within-group and between-group dissimilarity. However, in place of the aggregate community dissimilarity response used in change-point analysis, TITAN uses taxon-specific indicator value scores to assess the strength of association between each taxon and the environmental gradient. Indicator value scores are the product of cross-group relative abundance and within-group frequency of occurrence; occurrence frequency within each group is used to weight the relative abundance of each taxon such that a large abundance within a sample group only results in a higher score if the taxon also frequently occurs within that group. Indicator value scores are normalized on a scale from 0 to 100%, allowing for cross-taxa comparisons. The relative strength of indicator value scores on either side of candidate change points indicates whether each taxon shows a negative response (Z–) or positive response (Z+) to the environmental gradient.
TITAN calculates the probability (p) of randomly obtaining indicator value scores equal to or larger than observed values, using 500 random permutations. To assess uncertainty around change-points, the original data is then bootstrapped (500 replicates) and change-points are recalculated with each permutation; uncertainty is expressed as quantiles of change-point distribution. Narrow intervals between upper and lower change-point quantiles (i.e., 5–95%) reflect sharp, nonlinear responses in taxon abundance, whereas broad quantile intervals indicate linear or more gradual responses. Two diagnostic indices of indicator response quality are obtained from bootstrap resampling: indicator purity and reliability. Purity is the percent of bootstrap replicates with the same change-point response directions (positive or negative) as the observed response. Reliability is the percent of bootstrapped change-point indicator value scores that consistently have p-values below defined probability levels (i.e., p ≤ 0.05). We considered taxa as indicators if their purity and reliability indices were ≥ 95%, and only included responses of indicator species in our analyses (Baker and King 2010). We calculated threshold responses for the overall avian community, within 3 habitat guilds likely to be affected by anthropogenic forest disturbance: forest interior, early successional, and synanthropic (or human-adapted), and for indicator species within each of these guilds (Barton et al. 2016; Farwell et al. 2016; Langlois 2017).
Results
Land cover and distance metrics
Within 1-km radius landscapes surrounding the 1512 avian sampling points that contained shale gas, there was a mean 24.3 ± 0.4% (± SE) of total forest loss (range 0.5–83.3%), with a mean forest edge density of 104.3 ± 1.1 m/ha (range 4.2–237.8 m/ha). In general, the footprint of shale gas infrastructure within these landscapes was relatively small, occupying a mean 3.5 ± 0.1% of land cover (range 0.001–26.0%). Among landscapes evaluated, 97% contained < 10% shale gas cover, and 75% contained < 5% shale gas cover.
Of all 2589 points sampled, 2483 fell closest to anthropogenic forest edges related either to shale gas infrastructure or non-gas development (timber harvests excluded). Of these, 216 points were closest to a well pad (\(\overline{x}\) = 85 m, range 0–414 m), 1008 points were closest to non-gas development (\(\overline{x}\) = 160 m, range 0–1273 m), 288 points were closest to linear gas infrastructure (\(\overline{x}\) = 95 m, range 0–750 m), and 971 points were closest to linear non-gas infrastructure (\(\overline{x}\) = 145 m, range 0–823 m).
Avian surveys
We detected 122 species within 100 m of our sampling points, but excluded species detected only once, non-breeding migrants, and species not reliably sampled using point counts (e.g., gamebirds and other non-passerines, predominantly aerial species, and far-ranging species such as raptors). Of the 91 remaining species of passerines and near-passerines observed during surveys, 60 had sufficient detections (n ≥ 50) for estimation of detection probability (29,426 total detections, range 56–3,469 per species). Based on natural histories of each species (Rodewald 2015) and previous regional studies of avian habitat guilds (Thomas et al. 2014; Barton et al. 2016; Farwell et al. 2016), we placed 27 species in the forest interior habitat guild, 8 in the early successional guild, and 10 in the synanthropic (or human-adapted) guild (Table 1). We included the remaining 15 species with no clear habitat guild in threshold analyses of the overall avian community, but excluded these from guild-specific analyses (“no-assigned guild” in Table 1).
Forest cover thresholds
Within landscapes containing shale gas development, indicator species within the overall avian community decreased in abundance and frequency of occurrence beyond a threshold of 17.0% total forest loss, with a relatively narrow quantile interval that indicates an abrupt change point (CP) between 11.2–21.2% forest cover (Table 2). The overall avian community also decreased in abundance above a threshold of 95.9 m/ha forest edge density, with a quantile interval ranging from 87.7 to 103.5 m/ha. Within the overall avian community and forest interior guild, negative responses to both percent forest loss and forest edge density had relatively narrow 5–95% quantile intervals, which indicates confidence in the existence of threshold responses among birds to forest loss. These abrupt, non-linear declines at relatively low levels of overall forest loss and fragmentation were driven by forest interior dependent species, in addition to chestnut-sided warblers, a young forest specialist included in our early successional guild (Rodewald 2015; Fig. 2).

Plots showing species-specific threshold responses of indicator species within each habitat guild to a percent forest loss and b forest edge density (m/ha). Solid circles represent negative responses (z −) with corresponding species on the left axis, and open circles represent positive responses (z +) with corresponding species on the right axis. Circles are sized based on z scores with larger circles indicating greater significance, and lines represent 5–95% bootstrap quantiles. See Table 1 for species codes
Generally, positive threshold responses to forest loss and forest edge density occurred at higher thresholds than negative responses (Table 2). The overall avian community increased in abundance and frequency of occurrence above thresholds of 34.1% forest loss and 122.0 m/ha forest edge density. These community-level positive responses to forest loss were primarily driven by species within the early successional and synanthropic guilds, although a few forest interior species also showed positive threshold responses to either percent forest loss or forest edge density, or both (Fig. 2).
Both the early successional and synanthropic guilds showed similar positive threshold responses to percent forest loss at 30.5% and 36.5%, and to forest edge density at 116.5 and 123.8 m/ha, respectively (Table 2). Aside from the negative response to forest loss and forest edge density within the early successional guild (driven by chestnut-sided warblers), no other indicator species from the early successional or synanathropic guilds showed positive threshold responses to either percent forest loss or forest edge density (Fig. 2).
Distance thresholds
Negative threshold responses to distance from all anthropogenic forest edges generally occurred closer to human development and had narrower quantile intervals than positive thresholds (Table 3). In other words, species that increased farther from human development showed threshold responses at greater distances than species that decreased farther from human development. However, many quantile intervals for distance thresholds were quite broad, indicating more gradual or linear responses, particularly among positive indicator species that increased in abundance farther from human development.
The forest interior guild showed positive threshold responses closer to well pads (63.7 m) and linear shale gas infrastructure (59.9 m) than non-gas development (116.4 m) and non-gas linear infrastructure (264.6 m). Thus, among forest interior birds, edge effects of non-gas development generally extended farther into adjacent forests than edge effects associated with shale gas. However, quantile intervals extended up to 94.0 m from well pads and 356.7 m from linear shale gas infrastructure for the forest interior guild, and much further for individual species (up to 319.0 m from well pads and 546.1 m from linear gas; Fig. 3a–d), revealing substantial variability in species-specific responses to distance from human development among forest interior birds. We also observed a negative threshold response within the forest interior guild to distance from non-gas development (37.5 m) and non-gas linear infrastructure (63.8 m; Table 3) that was driven by yellow-throated warblers, the only forest interior bird to increase in abundance closer to human-created forest edges (Fig. 3a, c). There were no negative indicator species for distance from either well pads or linear shale gas, within the forest interior guild.

Plots showing species-specific threshold responses of all indicator species to distance from four categories of human development: a non-gas, non-linear development, b gas well pad development, c non-gas linear infrastructure, and d linear gas infrastructure. Thresholds are shown for all indicator species, including both positive (z +) and negative (z −) threshold responses. Circles are sized based on z scores with larger circles indicating greater significance, and lines represent 5–95% bootstrap quantiles. See Table 1 for species codes
In contrast, there were no positive indicator species for distance from any category of human development within the early successional guild, reflecting a general lack of species that increased farther from human development, within this guild. Blue jays were the only synanthropic species that was a positive indicator for distance from well pads (Table 3; Fig. 3), which indicates there was some quality of well pad development that caused blue jays to occur in higher abundances at greater distance from well pads. While both the early successional and synanthropic guilds showed negative distance thresholds to all four categories of human development, thresholds within the synanthropic guild occurred much closer to linear shale gas infrastructure, in particular, compared with the other categories of human development.
Discussion
Land cover thresholds
Our land cover analyses identified threshold responses to total amounts of forest loss and edge density in landscapes experiencing unconventional shale gas development (Table 2). Some threshold responses were quite abrupt with narrow quantile intervals, including negative responses to forest loss within the overall avian community at 17.0% and within the forest interior guild at 17.2%. These threshold values suggest that area-sensitive, forest interior dependent birds decreased abruptly at relatively low levels of overall forest loss, within the landscapes we evaluated. Our findings are consistent with other studies from the central Appalachian region, which documented declines in abundance and richness of forest interior birds in response to anthropogenic forest disturbance at relatively low levels of forest loss (Suarez-Rubio et al. 2013; Becker et al. 2015; Farwell et al. 2016; Langlois 2017). In landscapes altered by mountaintop removal coal mining, forest interior birds declined at even lower thresholds of forest disturbance, showing a negative threshold response at 4.4% forest loss (Becker et al. 2015). However, the broad quantile interval (2.0–76.8% forest loss) around this threshold suggests this was a more gradual or linear response than the abrupt threshold we observed among forest interior birds at ~ 15.4–24.8% forest loss, in landscapes experiencing shale gas development.
We also observed negative threshold responses to increasing forest edge density within the overall avian community at 95.9 m/ha, and within the forest interior guild at 96.0 m/ha (Table 2). However, 3 of 27 (11.1%) forest interior species showed positive threshold responses to forest loss, and 6 of 27 (22.2%) showed positive threshold responses to forest edge density (Fig. 2). This suggests some forest interior birds can tolerate a degree of habitat fragmentation within predominantly forested landscapes (Fahrig 2003), and may partly reflect the attraction of some forest interior species to canopy openings within large areas of mature forest (Buchanan and Hart 2012; Perkins and Wood 2014; Farwell et al. 2019). However, a predominance of unfragmented forest habitat was still important for 17 of 27 (63.0%) forest interior, area-sensitive species (Banks-Leite et al. 2010; Fig. 2).
In contrast, in central Appalachian forests impacted by mountaintop removal mining, birds showed negative threshold responses at comparatively low levels of forest edge density at 50.8 m/ha for the overall avian community and 22.9 m/ha for forest interior birds (Becker et al. 2015). The observed differences in bird tolerance of forest edge density in landscapes altered by shale gas compared with mountaintop removal coal mining may largely reflect differences in the footprint and configuration of these two types of energy development. Surface coal mines tend to occupy large, sprawling areas on mountain ridgetops, whereas shale gas typically creates a network of relatively small forest perforations connected by narrow linear infrastructure (Farwell et al. 2016; Langlois et al. 2017). Thus, increasing forest edge density in mining regions may be driven by larger disturbances with greater potential impacts to overall avian abundance, while the creation of new forest edges associated with shale gas is largely driven by smaller-scale disturbances and linear corridors.
Conversely, positive responses to forest loss can be interpreted as positive responses to increasing anthropogenic disturbance and availability of non-forest habitat. Positive responses to forest loss and forest edge density consistently occurred at higher thresholds than negative responses (Table 2, Fig. 2). This indicates that forest birds were negatively impacted by forest loss before early successional and synanthropic species benefitted. Again, this mirrors avian threshold responses reported in landscapes altered by mountaintop removal coal mining (Becker et al. 2015).
Distance thresholds
Our results support the occurrence of some nonlinear avian responses to distance from anthropogenic edges among indicator species, although quantile intervals for distance thresholds were generally quite broad for the overall avian community, within habitat guilds, and among indicator species. This suggests that distance threshold responses were typically gradual rather than sharply nonlinear. This may be a reflection of the wide range of sizes, shapes, and configurations of human development we evaluated, as well as varying species responses to anthropogenic disturbance. Previous studies of energy sector development have found similar variability in tolerance of energy-related infrastructure among birds, with some species appearing to be largely unaffected by energy development while other species avoid infrastructure (Gilbert and Chalfoun 2011; Kalyn Bogard and Davis 2014; Farwell et al. 2019).
In general, we found that species that increased farther from human development showed threshold responses at greater distances than species that decreased farther from human development. This indicates that benefits of anthropogenic disturbance for edge-associated species do not extend as far into disturbed forests as negative edge effects extend for forest interior species. For certain area-sensitive, forest interior dependent species (e.g., blue-headed vireo, Canada warbler, least flycatcher, veery), negative effects of anthropogenic edges extended much farther into affected forests (> 600 m for non-gas development and > 500 m for linear gas infrastructure; Fig. 3). These findings exceed the 50–200 m depth of edge effects generally reported for birds (Ries et al. 2004), and in other studies of shale energy development impacts on birds. Forest interior birds in northcentral Pennsylvania decreased in abundance up to 150 m from shale gas pipelines (Langlois 2017) and up to 250 m from shale gas well pads (Barton et al. 2016); however, these were the farthest extents of fixed-distance surveys conducted in each study, respectively. Our results were more consistent with a study of mixed-grass prairies affected by shale oil development in North Dakota, where grassland birds generally increased in abundance ≥ 214 m from road edges and ≥ 378 m from well pads, while individual species increased up to ≥ 469 m from road edges and ≥ 485 m from well pads (Thompson et al. 2015).
We also saw differences in threshold responses depending on type of human development, particularly in response to forest edges created by shale gas (linear and non-linear) compared with edges associated with non-gas development. Species within the forest interior guild, in particular, increased in abundance closer to forest edges associated with shale gas relative to non-gas development. This may reflect the smaller overall footprint of shale gas infrastructure compared with the other categories of human development we evaluated. However, it is worth noting that the forest interior guild showed similar positive threshold responses to distance from linear gas infrastructure and from well pad development, even though linear shale gas impacts tend to be narrower than land cover impacts associated with well pads. This suggests that shale gas pipelines and access roads have comparable negative edge effects on forest interior species to well pads, which may be due in part to the higher overall density of new forest edges created by linear gas infrastructure compared with well pad development (Farwell et al. 2016; Langlois et al. 2017).
In contrast, the early successional guild decreased in abundance farther from well pad development compared with other types of human development. This suggests there may be specific qualities of edge effects associated with well pad development that disturbance-dependent birds are exploiting. These qualities are likely related to the size and recency of canopy disturbance associated with well pad development, and resulting changes in plant community structure, composition, and associated availability of food resources and nest sites (King and Schlossberg 2014). The synanthropic guild decreased in abundance at comparable distances from well pads, non-gas development and non-gas linear infrastructure, but decreased abruptly very close to edges associated with shale gas linear infrastructure, compared with other types of development evaluated. This suggests that synanthropic species did not penetrate as far into forest edges adjacent to access roads and pipelines associated with shale gas, and that they remained close to these disturbances.
Management implications, caveats and conclusions
Ultimately, shale gas development is relatively recent in this region (≤ 10 years old at the time of the study), and it is reasonable to assume we have not yet observed the full range of successional impacts to affected forests, or the full response of species to ongoing changes. Our results highlight the importance of continued monitoring of biodiversity in the central Appalachian region, as shale gas development continues to age and expand in regional forests. By evaluating potential threshold responses in birds to anthropogenic forest disturbance across the Marcellus-Utica shale region, we addressed a knowledge gap in a region experiencing a rapid expansion of unconventional shale gas infrastructure. We found thresholds in bird abundance and frequency of occurrence as functions of the total amount of forest loss and fragmentation within landscapes experiencing shale gas development, and to a lesser extent as functions of distance from forest edges associated with shale gas and non-gas development. Such thresholds are predicted to occur in theoretical models (Lande 1987; Fahrig 1998; With and King 1999) and have been supported by regional studies of impacts of exurban development (Suarez-Rubio et al. 2013) and mountaintop removal mining (Becker et al. 2015) on forested landscapes, but before our study had not been assessed in the context of unconventional shale gas.
These thresholds can be used by conservation practitioners and resource managers to help identify particularly sensitive or vulnerable species, and to help predict the point at which landscapes may be on the verge of rapid ecological change (van der Hoek et al. 2015). However, the wide range of threshold values we observed among individual species underscores the fallacy of searching for a single, ‘one-size-fits-all’ threshold value for land cover change and edge effects (With and King 1999; Lindenmeyer and Luck 2005; Zuckerberg and Porter 2010). Rather than managing for minimum threshold values at which point large numbers of sensitive species will be lost, preventive management efforts could focus on managing landscapes above thresholds that will support the most sensitive species and species of highest conservation concern (Robbins et al. 1989; Betts et al. 2007; van der Hoek et al. 2015). Landscapes maintained at these target levels are also likely to benefit species less sensitive to development. For example, we found that forested landscapes with less than 17.2% overall forest loss supported most forest interior species, although some sensitive species decreased in abundance and frequency of occurrence at lower thresholds ranging from 8.7 to 15.9% forest loss. In contrast, species in the early successional and synanthropic guilds did not increase in abundance and frequency of occurrence in landscapes with more than 30.5–36.5% forest loss, suggesting these species are not likely to benefit optimally from anthropogenic disturbance in heavily forested landscapes below these thresholds of forest loss. It is also important to note that not all species evaluated showed threshold responses, including habitat generalists such as chipping sparrows and mourning doves, but also some forest associated species such as brown creepers and Kentucky warblers. Our findings suggest that threshold responses to forest loss and fragmentation are common but not ubiquitous (Zuckerberg and Porter 2010).
Lastly, our analyses were based on survey data collected during a 2-year period, and were intended as a preliminary assessment of potential nonlinear responses in birds to relatively recent land cover disturbance associated with shale gas. Our results suggest multiple forest interior dependent species in the Marcellus-Utica region of central Appalachia in eastern USA are exhibiting threshold responses to forest loss and fragmentation associated with shale gas. These responses have important conservation implications given that development is ongoing, and a number of new pipelines that will pass through interior forests in the central Appalachian region have been approved or are currently under construction (FERC 2019). Our findings indicate that efforts to avoid shale gas development in relatively undisturbed landscapes with low proportions of forest loss (< 15.4–24.8% forest loss) while concentrating new development in areas already disturbed by humans can help mitigate negative impacts to forest interior dependent species while maximizing benefits for early successional species.
References
AMJV (Appalachian Mountains Joint Venture) (2015) Priority landbird species. http://amjv.org/documents/Priority_Landbird_Species.pdf. Accessed 6 Mar 2018
Anderson MG, Clark M, Sheldon AO (2012) Resilient sites for terrestrial conservation in the Northeast and Mid-Atlantic region. The Nature Conservancy, Eastern Conservation Science, Boston, p 168
Andren H (1996) Population responses to habitat fragmentation: statistical power and the random sample hypothesis. Oikos 76:235–242
Baker ME, King RS (2010) A new method for detecting and interpreting biodiversity and ecological community thresholds. Methods Ecol Evol 1:25–37
Baker ME, King RS (2013) Of TITAN and straw men: an appeal for greater understanding of community data. Freshw Sci 32:489–506
Baker ME, King RS, Kahle D (2015) TITAN2: threshold indicator taxa analysis. R package version 2.1
Banks-Leite C, Ewers RM, Metzger J-P (2010) Edge effects as the principal cause of area effects on birds in fragmented secondary forest. Oikos 119:918–926
Barlow KM, Mortensen DA, Drohan PJ, Averill KM (2017) Unconventional gas development facilitates plant invasions. J Environ Manage 202:208–216
Barton EP, Pabian SE, Brittingham MC (2016) Bird community response to Marcellus shale gas development. J Wildl Manage 80:1301–1313
Bayne EM, Dale BC (2011) Effects of energy development on songbirds. In: Naugle DE (ed) Energy development and wildlife conservation in Western North America. Island Press, Washington, D.C., pp 95–114
Bayne EM, Van Wilgenburg SL, Boutin S, Hobson KA (2005) Modeling and field-testing of Ovenbird (Seiurus aurocapillus) responses to boreal forest dissection by energy sector development at multiple spatial scales. Landsc Ecol 20:203–216
Becker DA, Wood PB, Strager MP, Mazzarella C (2015) Impacts of mountaintop mining on terrestrial ecosystem integrity: identifying landscape thresholds for avian species in the central Appalachians, United States. Landsc Ecol 30:339–356
Bestelmeyer BT (2006) Threshold concepts and their use in rangeland management and restoration: the good, the bad, and the insidious. Restor Ecol 14:325–329
Bestelmeyer BT, Ellison AM, Fraser WR, Gorman KB, Holbrook SJ, Laney CM, Ohman MD, Peters DPC, Pillsbury FC, Rassweiler A, Schmitt RJ, Sharma S (2011) Analysis of abrupt transitions in ecological systems. Ecosphere 2:129
Betts MG, Forbes GJ, Diamond AW (2007) Thresholds in songbird occurrence in relation to landscape structure. Conserv Biol 21:1046–1058
Bibby CJ, Burgess ND, Hill DA (1992) Bird census techniques. Academic Press, London
Boulinier T, Nichols JD, Hines JE, Sauer JR, Flather CH, Pollock KH (2001) Forest fragmentation and bird community dynamics: inference at regional scales. Ecology 82:1159–1169
Brennan JM, Bender DJ, Contreras TA, Fahrig L (2002) Focal patch landscape studies for wildlife management. In: Wu J, Taylor WW (eds) Optimizing sampling effort across scales: integrating landscape ecology into natural resource management. Cambridge University Press, Cambridge, pp 68–91
Brittingham MC, Maloney KO, Farag AM, Harper DD, Bowen ZH (2014) Ecological risks of shale oil and gas development to wildlife, aquatic resources and their habitats. Environ Sci Technol 48:11034–11047
Buchanan ML, Hart JL (2012) Canopy disturbance history of old-growth Quercus alba sites in the eastern United States: examination of long-term trends and broad-scale patterns. For Ecol Manage 267:28–39
Canterbury GE, Martin TE, Petit DR, Petit LJ, Bradford DF (2000) Bird communities and habitat as ecological indicators of forest condition in regional monitoring. Conserv Biol 14:544–558
Carignan V, Villard MA (2002) Selecting indicator species to monitor ecological integrity: a review. Environ Monit Assess 78:45–61
Desrochers A, Renaud C, Hochachka WM, Cadman M (2010) Area-sensitivity by forest songbirds: theoretical and practical implications of scale-dependency. Ecography 33:921–931
Dufrene M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr 67:345–366
ESRI (Environmental Research Systems Institute) (2016) ArcGIS Desktop: Release 10.4. Redlands, CA, USA
Fahrig L (1998) When does fragmentation of breeding habitat affect population survival? Ecol Model 105:273–292
Fahrig L (1999) Forest loss and fragmentation: which has the greater effect on persistence of forest-dwelling animals? In: Rochelle JA, Lehmann LA, Wisniewski J (eds) Forest fragmentation: wildlife and management implications. Brill Academic Publishing, Leiden, pp 87–95
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst 34:487–515
Farwell LS, Wood PB, Brown DJ, Sheehan J (2019) Proximity to unconventional shale gas infrastructure alters breeding songbird abundance and distribution. Condor 121:1–20
Farwell LS, Wood PB, Sheehan J, George GA (2016) Shale gas development effects on the songbird community in a central Appalachian forest. Biol Cons 201:78–91
Federal Energy Regulatory Commission (FERC) (2019) Approved major natural gas pipeline projects. https://www.ferc.gov/industries/gas/indus-act/pipelines/approved-projects.asp. Accessed 7 April 2020
Foley MM, Martone RG, Fox MD, Kappel CV, Mease LA, Erickson AL, Halpern BS, Selkoe KA, Taylor P, Scarborough C (2015) Using ecological thresholds to inform resource management: current options and future possibilities. Front Mar Sci 2:95
Forsman JT, Reunanen P, Jokimaki J, Monkkonen M (2010) The effects of small-scale disturbance on forest birds: a meta-analysis. Can J For Res 40:1833–1842
Gaston KJ, Blackburn TM, Goldewijk KK (2003) Habitat conversion and global avian biodiversity loss. Proc R Soc B 270(1521):1293–1300
Gilbert MM, Chalfoun AD (2011) Energy development affects populations of sagebrush songbirds in Wyoming. J Wildl Manage 75:816–824
Gutzwiller KJ, Riffell SK, Flather CH (2015) Avian abundance thresholds, human-altered landscapes, and the challenge of assemblage-level conservation. Landsc Ecol 30(1):2095–2110
Hanski I, Hammond P (1995) Biodiversity in boreal forests. Trends Ecol Evol 10(1):5–6
Huggett AJ (2005) The concept and utility of ‘ecological thresholds’ in biodiversity conservation. Biol Cons 124:301–310
Huisman WHT, Attenborough K (1991) Reverberation and attenuation in a pine forest. J Acoust Soc Am 90:2664–2677
IUCN (International Union for Conservation of Nature) (2019) The IUCN red list of threatened species, v. 2019-1. http://www.iucnredlist.org. Accessed 15 Aug 2019
Kalyn Bogard HJ, Davis SK (2014) Grassland songbirds exhibit variable responses to the proximity and density of natural gas wells. J Wildl Manage 78:471–482
King DI, Schlossberg S (2014) Synthesis of the conservation value of the early-successional stage in forests of eastern North America. For Ecol Manage 324:186–195
King RS, Richardson CJ (2003) Integrating bioassessment and ecological risk assessment: an approach to developing numerical water-quality criteria. Environ Manage 31:795–809
Kiviat E (2013) Risks to biodiversity from hydraulic fracturing for natural gas in the Marcellus and Utica shales. Ann N Y Acad Sci 1286:1–14
Krementz DG, Powell LA (2000) Breeding season demography and movements of eastern towhees at the Savannah River Site, South Carolina. Wilson Bull 112:243–248
Lande R (1987) Extinction thresholds in demographic models of territorial populations. Am Nat 130:624–635
Lang JD, Powell LA, Krementz DG, Conroy MJ (2002) Wood thrush movements and habitat use: effects of forest management for red-cockaded woodpeckers. Auk 119:109–124
Langlois LA (2017) Effects of Marcellus shale gas infrastructure on forest fragmentation and bird communities in northcentral Pennsylvania. Dissertation, Pennsylvania State University, State College, USA
Langlois LA, Drohan PJ, Brittingham MC (2017) Linear infrastructure drives habitat conversion and forest fragmentation associated with Marcellus shale gas development in a forested landscape. J Environ Manage 197:167–176
Leonard TD, Taylor PD, Warkentin IG (2008) Landscape structure and spatial scale affect space use by songbirds in naturally patchy and harvested boreal forests. Condor 110(3):467–481
Lichstein JW, Simons TR, Franzreb KE (2002) Landscape effects on breeding songbird abundance in managed forests. Ecol Appl 12(3):836–857
Lindenmayer DB, Luck G (2005) Synthesis: thresholds in conservation and management. Biol Conserv 124:351–354
Luck GW (2005) An introduction to ecological thresholds. Landsc Ecol 124:299–300
Machtans CS (2006) Songbird response to seismic lines in the western boreal forest: a manipulative experiment. Can J Zool 84:1421–1430
Marzluff JM, Ewing K (2001) Restoration of fragmented landscapes for the conservation of birds: a general framework and specific recommendations for urbanizing landscapes. Restor Ecol 9:280–292
McGarigal K, Cushman SA, Ene E (2012) FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. University of Massachusetts, Amherst. Amherst, MA. https://www.umass.edu/landeco/research/fragstats/fragstats.html. Accessed 12 July 2017
MCOR (Marcellus Center for Outreach and Research) (2019) Tri-state unconventional shale wells drilled by year (PA, OH, WV). https://www.marcellus.psu.edu/resources-maps-graphics.html. Accessed 7 April 2020
Mitchell MS, Rutzmoser SH, Bently Wigley T, Loehle C, Gerwin JA, Keyser PD, Lancia RA, Perry RW, Reynolds CJ, Thill RE, Weih R, White D, Wood PB (2006) Relationships between avian richness and landscape structure at multiple scales using multiple landscapes. For Ecol Manage 221:155–169
Mutter M, Pavlacky DC, Van Lanen NJ, Grenyer R (2015) Evaluating the impact of gas extraction infrastructure on the occupancy of sagebrush-obligate songbirds. Ecol Appl 25:1175–1186
NAIP (National Agriculture Imagery Program) (2016) U.S. Department of Agriculture Farm Service Agency: NAIP imagery. https://www.fsa.usda.gov/programs-and-services/aerial-photography/imagery-programs/naip-imagery/. Accessed 12 July 2017
Nenninger HR, Koper N (2018) Effects of conventional oil wells on grassland songbird abundance are caused by presence of infrastructure, not noise. Biol Conserv 218:124–133
O’Connell TJ, Bishop JA, Brooks RP (2007) Sub-sampling data from the North American Breeding Bird Survey for application to the Bird Community Index, an indicator of ecological condition. Ecol Ind 7:679–691
Paton PWC (1994) The effects of edge on avian nest success: how strong is the evidence? Conserv Biol 8:17–26
Perkins KA, Wood PB (2014) Selection of forest canopy gaps by male cerulean warblers in West Virginia. Wilson J Ornithol 126:288–297
Pickering J, Kays R, Meier A, Andrew S, Yatskievych K (2002) The Appalachians. In: Mittermeier RA, Mittemeier CG, Gil PR, Pilgrim J, Fonesca G, Brooks T, Konstant WR (eds) Wilderness: earth’s last wild places. Conservation International, Washington DC, pp 458–467
Qian SS, King RS, Richardson CJ (2003) Two methods for the detection of environmental thresholds. Ecol Model 166:87–97
R Core Team (2017) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. Accessed 12 July 2017
Radford JQ, Bennett AF, Cheers GJ (2005) Landscape-level thresholds of habitat cover for woodland-dependent birds. Biol Conserv 124:317–337
Ralph CJ, Droege S, Sauer JR (1995) Managing and monitoring birds using point counts: standards and applications. USDA Forest Service General Technical Report PSW-GTR-149. USDA Forest Service Pacific Southwest Research Station, Albany, pp 161–168
Richardson ML, Wilson BA, Aiuto DAS, Crosby JE, Alonso A, Dallmeier F, Golinksi GK (2017) A review of the impact of pipelines and power lines on biodiversity and strategies for mitigation. Biodivers Conserv 26:1801–1815
Ries L, Fletcher RJ, Battin J, Sisk TD (2004) Ecological responses to habitat edges: mechanisms, models, and variability explained. Annu Rev Ecol Evol Syst 35:491–522
Robbins CS, Dawson DK, Dowell BA (1989) Habitat area requirements of breeding forest birds of the Middle Atlantic States. Wildl Monogr 103:3–34
Rodewald PG (ed) (2015) The birds of North America. Cornell Laboratory of Ornithology, Ithaca, New York
Rueda M, Hawkins BA, Morales-Castilla I, Vidanes RM, Ferrero M, Rodriguez MA (2013) Does fragmentation increase extinction thresholds? A European-wide test with seven forest birds. Glob Ecol Biogeogr 22:1282–1292
Schmiegelow FKA, Machtans CS, Hannon SJ (1997) Are boreal birds resilient to forest fragmentation? An experimental study of short-term community responses. Ecology 78(6):1914–1932
Sólymos P, Matsuoka SM, Bayne EM, Lele SR, Fontaine P, Cummings SG, Stralberg D, Schmiegelow FKA, Song SJ (2013) Calibrating indices of avian density from non-standardized survey data: making the most of a messy situation. Methods Ecol Evol 4:1047–1058
Sólymos P, Moreno M, Lele SR (2016) detect: Analyzing wildlife data with detection error. R package version 0.4-0. Accessed 12 July 2017
Souther S, Tingley MW, Popescu VD, Hayman DTS, Ryan ME, Graves TA, Harth B, Terrell K (2014) Biotic impacts of energy development from shale: research priorities and knowledge gaps. Front Ecol Environ 12:330–338
Suarez-Rubio M, Wilson S, Leimgruber P, Lookingbill T (2013) Threshold responses of forest birds to landscape changes around exurban development. PLoS ONE 8:e67593
Thomas EH, Brittingham MC, Stoleson SH (2014) Conventional oil and gas development alters forest songbird communities. J Wildl Manage 78:293–306
Thompson SJ, Johnson DH, Niemuth ND, Ribic CA (2015) Avoidance of unconventional oil wells and roads exacerbates habitat loss for grassland birds in the North American great plains. Biol Conserv 192:82–90
USEIA (U.S. Energy Information Administration) (2017) Marcellus Shale play geology overview. https://www.eia.gov/maps/pdf/MarcellusPlayUpdate_Jan2017.pdf. Accessed 7 April 2020
USEIA (U.S. Energy Information Administration) (2020) Annual Energy Outlook. https://www.eia.gov/outlooks/aeo/. Accessed 7 April 2020
USEPA (U.S. Environmental Protection Agency) (2013) Level III and IV ecoregions of the continental United States: Corvallis, Oregon, U.S. EPA—National Health and Environmental Effects Research Laboratory, map scale 1:7,500,000. https://www.epa.gov/eco-research/level-iii-and-iv-ecoregions-continental-united-states. Accessed 12 July 2017
USFWS (U.S. Fish and Wildlife Service) (2008) Birds of conservation concern 2008. United States Department of Interior, Fish and Wildlife Service, Division of Migratory Bird Management, Arlington, Virginia. http://www.fws.gov/migratorybirds/. Accessed 6 Mar 2018
van der Hoek Y, Renfrew R, Manne LL (2013) Assessing regional and interspecific variation in threshold responses of forest breeding birds through broad scale analyses. PLoS ONE 8:e55996
van der Hoek Y, Zuckerberg B, Manne LL (2015) Application of habitat thresholds in conservation: considerations, limitations, and future directions. Glob Ecol Conserv 3:736–743
van der Ree R, Smith DJ, Grilo C (2015) The ecological effects of linear infrastructure and traffic. In: van der Ree R, Smith DJ, Grilo C (eds) Handbook of road ecology. Wiley, Chichester, pp 1–9
Warren TL, Betts MG, Diamond AW, Forbes GJ (2005) The influence of local habitat and landscape composition on cavity-nesting birds in a forested mosaic. For Ecol Manage 214:331–343
With KA, King AW (1999) Dispersal success on fractal landscapes: a consequence of lacunarity thresholds. Landsc Ecol 14:73–82
Wood PB, Bosworth S, Dettmers R (2006) Cerulean warbler abundance and occurrence relative to large-scale edge and habitat characteristics. Condor 108:154–165
Yip DA, Bayne EM, Solymos P, Campbell J, Proppe D (2017) Sound attenuation in forest and roadside environments: implications for avian point-count surveys. Condor 119:73–84
Young J, Maloney KO, Slonecker ET, Milheim LE, Siripoonsup D (2018) Canopy volume removal from oil and gas development activity in the upper Susquehanna River basin in Pennsylvania and New York (USA): an assessment using lidar data. J Environ Manage 222:66–75
Zuckerberg B, Desrochers A, Hochachka WM, Fink D, Koenig WD, Dickinson JL (2012) Overlapping landscapes: a persistent, but misdirected concern when collecting and analyzing ecological data. J Wildl Manage 76:1072–1080
Zuckerberg B, Porter WF (2010) Thresholds in the long-term responses of breeding birds to forest cover and fragmentation. Biol Conserv 143:952–962
Acknowledgements
This research was funded by the U.S. Fish and Wildlife Service in collaboration with the Appalachian Mountains Joint Venture, and by a dissertation fellowship from the Southern Regional Education Board. The WV Division of Natural Resources, PA Department of Conservation and Natural Resources, and PA Game Commission provided support in the field and access to field housing. We are grateful to the Western Pennsylvania Conservancy, WV Host Farms Program, the many landowners who granted access to private property, and especially to the technicians who collected field data and assisted with land cover digitization. T Fearer, JT Anderson, DJ Brown, BE McNeil, and GT Merovich provided guidance during project design. Two anonymous reviewers provided valuable comments that greatly improved this manuscript. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Farwell, L.S., Wood, P.B., Dettmers, R. et al. Threshold responses of songbirds to forest loss and fragmentation across the Marcellus-Utica shale gas region of central Appalachia, USA. Landscape Ecol 35, 1353–1370 (2020). https://doi.org/10.1007/s10980-020-01019-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-020-01019-3