
Abstract
Context
Identifying animals movement through the landscape and delineating key corridors is critical for effective management and conservation. Still, assessments of space-use patterns and landscape connectivity are subjected to many limitations, especially in large scales.
Objectives
The main objective of this study was to assess functional connectivity for four focal mammal species with varying dispersal abilities and diets, across protected areas in a transnational region where only scarce information on movement patterns exists.
Methods
We used models relying on circuit theory, multiple layers of spatial information and graph-theoretical analysis. We delineated potential pathways suitable for species movement and evaluated connectivity status of the protected areas network in the Balkan Peninsula, southeastern Europe. Models were parameterized by combining information from experts and scientific literature, or by applying allometric equations, while novel connectivity metrics were developed.
Results
We identified four key, suitable corridors within the transnational study region. The largest one crossed three countries, highlighting the need for international conservation efforts. For species with higher dispersal abilities, the network of protected areas appeared to be well connected and robust while for others it consisted of numerous isolated sites, raising the need for species-specific management plans.
Conclusions
Our study serves as an example of how to set monitoring and conservation priorities in data-poor regions. Our findings highlight the need to identify a number of ecological corridors that could facilitate movement of multiple species with different functional traits and habitat preferences. This information is proved to be critical for setting spatially explicit conservation plans at local, regional and transnational scales.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Habitat loss is listed as a major threat to biodiversity at local, regional and global scales (Pimm and Raven 2000; Haddad et al. 2015). Compounding the effects of pure habitat loss, fragmentation further alters the structure and configuration of the landscape mosaic, generating physical barriers to biological flows (i.e. genes, individuals, populations) (Baguette et al. 2013). As a result, an increasing number of evidence, originating from both empirical and theoretical studies, sought for the need to maintain or recreate linkages between groups of remnant habitat patches (Fahrig 2003).
Habitat corridors could offer safe passes for species, enhancing the exchange of individuals among isolated patches, and thus benefit population viability through the rescue effect (Cushman et al. 2013; Pinaud et al. 2018). Habitat corridors could allow the maintenance of ecosystem health, functionality and services by supporting ecological processes (e.g. recycling of nutrients, pollination) (Christie and Knowles 2015). Counting for the ongoing and dynamic changes at the landscape and community level, which are triggered by climate change, habitat corridors could offer the means for species to capture their ecological and climatic niche, through shifting their distributions and/or colonizing neighboring sites (Brodie et al. 2012; Whitmee and Orme 2013).
Nevertheless, there is still an ongoing scientific discussion on the most appropriate tools and methods to be used for detecting corridors that could maximize the benefits of ecological connectivity across the heterogeneous landscapes. Least cost path modeling (LCP) represents a popular method for assessing connectivity, relying on the transformation of the study area into a “cost” map, where cost values indicate the opposition of the landscape to animal movement (Etherington and Holland 2013). Connectivity among habitat patches is then calculated as the lowest cost path from one patch to the other (see Adriaensen et al. 2003). The circuit theory modeling (McRae et al. 2008) is an alternative method widely applied for assessing connectivity and delineating ecological corridors (Dickson et al. 2019). Similarly to the LCP, in circuit theory modeling, the landscape is transformed into a cost map (called resistance surface), but instead of producing a single path with the lowest cost, a metric called effective resistance, indicating landscape’s opposition to movement between two patches, is calculated through multiple pathways. Based on this modeling procedure, a new landscape raster is created (called current map), where every cell is assigned a value that reflects the probability of an animal passing through it while traveling from one habitat patch to another (McRae et al. 2008).
Some weaknesses of LCP modeling that are related to the cost map construction could be noted also in circuit theory (Sawyer et al. 2011). Circuit theory could be less sensitive to the number of pixels and the Euclidean distance between patches; still, it could be more sensitive to any form of data aggregation (Marrotte and Bowman 2017). While LCP modeling assumes that individuals have complete knowledge of the landscape composition (Adriaensen et al. 2003; Etherington 2016), circuit theory models assume that individuals move randomly, based on landscape resistance, without having any prior information for landscape composition (McRae 2006). Therefore, circuit theory-based connectivity models offer a metric of isolation and connectance, which is calculated through multiple pathways (McRae et al. 2008; Pelletier et al. 2014). This methodological advantage further allows to delineate insights for any potential movement corridor across the landscape.
Recognizing the necessity to identify, propose and design effective habitat corridors, information on functional and structural properties of the landscape and their association with organism’s decision on habitat use and movement needs to be collected and evaluated (Hagen et al. 2012; Abrahms et al. 2017). Such information is often collected through intense field observations and/or satellite telemetry (Sawyer et al. 2011; Abrahms et al. 2017). Data collected through these methods offer critical insights on the spatial properties of the landscape (i.e. structure, configurations, dynamics) that control the biological flows within and between habitat patches (Cooke et al. 2004; Nathan et al. 2008). In this way, it is possible to determine the scales and extent at which management and conservation actions should be taken (Webster et al. 2002). Still, collection of accurate data on habitat use and movement is highly demanding as it could be subjected to many practical constrains (e.g. time, effort, personnel, financial costs; Hebblewhite and Haydon 2010). As a result, for the vast majority of the species, there is a lack of any detailed spatial information on habitat use and movement patterns (Zeller et al. 2012). Even for species for which such information is available, the spatial cover and temporal scales of the data are limited (e.g. short term monitoring data focused mainly on a given number of individuals within a population and not the full species range; Richard and Amstrong 2010; Zeller et al. 2017).
Effective conservation requires the parallel protection of multiple species, with different functional traits [e.g. dispersal ability, foraging behavior, minimum area requirements (MAR)], which determine their habitat preferences and their movement patterns (Nathan et al. 2008; Pe’er et al. 2014). This global principle is fundamental for the design, selection and establishment of networks of protected areas (PAs) that constitute the cornerstone of biodiversity conservation. Protected areas purpose is to retain habitat preservation for a list of species by applying an increased level of protection within their geographical boundaries. Therefore, ensuring PAs connectivity across the landscape mosaic could significantly improve their efficiency (Saura et al. 2018). Still, in practice, the assesment of habitat connectivity requires a wealth of information on species dispersal patterns and behavior, in order to identify potential corridors that could support multiple species movement over heterogeneous landscapes (Rondinini et al. 2011; Brodie et al. 2015; Mimet et al. 2016; Santini et al. 2016; Dilkina et al. 2017; Sahraoui et al. 2017).
All limitations regarding available information on movement patterns and habitat use maximize in regions where different economic and sociopolitical conditions might affect the collection and compilation of ecological data (Amano and Sutherland 2013). As an example, the Balkan Peninsula, in the southeastern Europe, is shared by many nations that have different political priorities, follow incompatible environmental policies, perceive conservation priorities in various ways and contribute differently to conservation actions (Radenković et al. 2017). The Balkan Peninsula is a crossroad of Europe, Asia and Africa, that constitutes a European biodiversity hotspot with a very high endemism for many taxonomic groups (Banarescu 2004; Weiss and Ferrand 2007). As an encouraging message for conservation, the number of PAs in the region is increasing (UNEP-WCMC, IUCN, NGS 2018); still, it is not known how well these PAs are connected.
In the present study, we proposed and applied a multi-species landscape connectivity framework over the Balkan Peninsula, exploring connectivity patterns for a set of mammal species (Canis lupus, Capreolus capreolus, Vulpes vulpes, Ursus arctos). We investigated potential corridors among 1356 PAs which are hosted in a transnational region of more than 250,000 km2. Given that there is a scarcity of information on movement patterns across the broader region, with no prior knowledge on the potential importance of landscape patches acting as ecological corridors or stepping stones at this scale, we developed a series of landscape models by taking advantage of circuit theory (McRae et al. 2008). Model parameters were selected and assigned by combining information obtained through expert interaction and scientific literature, or were estimated by using allometric equations. Connectivity metrics were developed and a graph-theoretical analysis was applied to delineate the overall connectivity properties of the network of PAs hosted in the region. We highlight that transnational monitoring, scientific cooperation and a join set of priorities and targets for PAs are critical steps towards the efficient conservation of heterogeneous transnational landscapes.
Methods
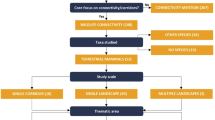
The methodology applied (Fig. 1) in the present study consists of seven steps: (a) define the study area, (b) select a number of focal species with different functional traits and ecological requirements, (c) define the circuit theory-based framework for modeling connectivity, (d) engage experts to select background layers and evaluate the potential impact of landscape features on animal movement, (e) construct resistance surfaces for modeling species, (f) demonstrate connectivity pathways for each species and produce consensus map by summarizing information from all species, (g) develop and apply a series of metrics to quantify connectivity properties at the landscape scale.

Flowchart representing the methodological steps undertook for the study
Study area
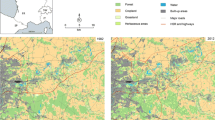
The study area expands over the boundaries of four countries (i.e. Greece, the Republic of North Macedonia, Albania and Bulgaria) located in the Balkan Peninsula, southeastern Europe. We used the World Database on Protected Areas (UNEP-WCMC, IUCN 2018) to identify the PAs hosted within the study area. Initially, a total of 2206 PAs were identified. Protected areas which overlap with each other were spatially merged, resulting to a total of 1356 sites that were used for the analyses (Fig. 2).

The study area, composed of four countries (i.e. Greece, the Republic of North Macedonia, Albania and Bulgaria). Protected areas are represented with different colours, depending on whether they are large enough to meet the minimum area requirements for the different species (1st: Canis lupus, 2nd: Capreolus capreolus, 3rd: Vulpes vulpes, 4th: Ursus arctos)
Species
We selected four main, large mammal species (C. lupus, C. capreolus, V. vulpes, U. arctos) hosted in the region (Hoffmann and Sillero-Zubiri 2016; Lovari et al. 2016; McLellan et al. 2017; Boitani et al. 2018). Each one of these species is characterized by different physiological (e.g. nutritional choices, body weight) and behavioral (e.g. habitat preferences and movement barriers) attributes. Information on species’ body weight was extracted from the literature (Online Resource). Based on body weight and trophic status of each of the four species, we estimated the minimum area required to cover their biological needs (Online Resource Table S1). To calculate MAR for every species, we used equations proposed by Pe’er et al. (2014). Protected areas with total surface smaller than the estimated MAR of a given species were not included in the landscape connectivity models developed (see below) for this specific species (Fig. 2). Dispersal capacity of each species was calculated based on allometric equations (Santini et al. 2013), using as inputs data on body weight and home range. Data on home range were derived from PanTHERIA (Jones et al. 2009).
Circuit theory-based connectivity models
In order to assess the functional connectivity for the studied species, we used Circuitscape software (v4.0, www.circuitscape.org). As a first step, the landscape mosaic, that consisted of several background layers affecting animals movement, was transferred into raster maps of 1 km2 resolution. Next, these raster maps were converted into a resistance surface, with each cell getting a value based on the properties of the background layers (see next section) reflecting its opposition to species movement. The landscape raster cells represented the circuit nodes, where neighboring nodes were connected by resistors. The cells within the boundaries of PAs were used as the source and exit nodes of the circuit (i.e. focal nodes). For each pair of PAs, one PA was assumed to be the starting point (i.e. the source node), while the other was considered as the ending point (i.e. the exit node). The starting point was treated as connected to a current source of 1 Amp, while the ending point was assumed to be grounded (the exit of the circuit). The current was diffused from the starting point through the surface, up to the ending point (McRae et al. 2008).
The output of this modeling procedure was a current map, with cell values indicating the amount of current that passed through it. Higher current values indicated areas through which animals were more likely to move. In addition, the effective resistance between each pair of PAs was calculated, with higher values indicating lower permeability to movement (McRae et al. 2008).
Background layers
A total of eleven layers were used to define the landscape properties in respect to the potential movement permeability of the studied species (Online Resource Table S2). To select these layers we originally used information from the literature, next we gathered available spatial datasets and further received inputs from experts.
The first five layers were built upon five main land use types (i.e. (a) artificial surfaces, (b) agricultural areas, (c) forest and semi-natural areas, (d) wetlands and (e) water bodies). For each one of these land use types, the percentage cover in every cell was calculated, resulting to a distinct background layer. Data on land use types were obtained from the CORINE (Co-ORdinated INformation on the Environment) Land Cover project of the European Environment Agency (EEA 2012), while the spatial resolution of the database was 100 m. For each background layer, cells with no presence of the respective land use type were assigned with the value of zero, while they got the value of one in case of full coverage.
To account for the impact of linear transportation infrastructure, two additional background layers were developed by combining information from roads and railways. The first one represented the distance of each cell from linear transportation, while the second one represented the linear transportation density. The spatial distribution of roads and railways was derived from OpenStreetMap (download.geofabrik.de). The original road data contained information on various road categories expanding across Europe. Here, we maintained only the major roads (motorway, trunk, primary, secondary, tertiary) along with their links (slip roads, ramps), as these road categories are acknowledged for posing barriers for large mammal movement (Benitez-Lopez et al. 2010; van der Ree et al. 2015). The road network was then integrated with the distribution map of railways to provide a comprehensive map of linear transportation. Each cell was assigned a value equal to the shortest distance from the cell’s center to any linear transportation feature. To assign a value on linear transportation density, we calculated the total length of the linear elements enclosed within a given radius from the center of the cell. The radius was assigned to 707 m, so that the circle drawn from the center of each 1 km2 cell (i.e. the resolution of the landscape raster) would enclose the entire cell’s surface as intersects its corners.
Three background layers were created in order to represent topographic features. The first of these layers was regarding elevation data derived from Copernicus, the European Programme for Earth Observation, and consisted of the European Digital Elevation Model (EU-DEM v1.1) with a 25 m resolution and vertical accuracy of ± 7 m root mean square error (RMSE). The elevation data layer was further used to create the layers of slope and Terrain Ruggedness Index (Riley et al. 1999). Slope layer represented the biggest slope faced from the cell to the neighboring cells. Terrain Ruggedness Index described the variation of a cell’s elevation and its neighboring cells.
The final layer represented human population density; data were derived from European Commission (Schiavina et al. 2019).
Resistance surfaces
To model the effort needed by an animal to move through a particular cell of the landscape we produced resistance surfaces by extracting information from the eleven background layers. To overcome the lack of information regarding the potential impact of the different landscape features to animals movement in the study region, we engaged a number of experts. Next, we asked the experts to complete a questionnaire, evaluating the selected layers regarding their opposition to animal movement. As a result, every single cell of each layer received a resistance value ranging from 1 (minimum resistance to movement) to 100 (maximum resistance to movement) (for more details, also see Online Recourse).
Acknowledging that a selected layer might be more critical determinant for a species movement than other layers, we applied a weighting scheme to define the final resistance surfaces. Therefore, for each layer and species, we asked experts to assign a weight, ranging from zero (i.e. the layer has no impact on movement) to ten (i.e. the layer is highly critical for movement decisions).
The final resistance values (Online Resource Table S3) and weights (Online Resource Table S4) were calculated as the average of the values given by each expert. The resistance values were used to transform the background layers to resistance layers, indicating their opposition to movement. The resistance surfaces were then created as a weighted linear combination of the resistance layers and their corresponding weights.
Consensus map
Based on the resistance surfaces, we used Circuitscape to produce one current map for each species. Then, in order to identify potential differences and similarities between the current maps of the different species, we normalized each map, by dividing the current value of every cell with the maximum obtained current value. A consensus current map was further constructed as the average of the four normalized current maps, and was used to delineate key corridors that could be suitable for all four species.
Quantifying connectivity properties
Statistical analysis and novel connectivity metrics
In order to investigate whether the magnitude of the current values assigned across the landscape differ among species, we used Kruskal–Wallis H test. The Dunn’s post hoc test was applied in order to determine significant differences between species. Two species are likely to share the same corridors and demonstrate a similar preference in landscape use if the assigned current values are highly correlated. On the contrary, two species are likely to have opposite preferences and different capacities to cross the landscape if the assigned current values are significantly negatively correlated. To delineate these patterns, we applied a pairwise Spearman’s rank-order correlation analysis for all the studied species.
To identify PAs that support overall connectivity of the network, we developed two indices. The first index called current score was calculated as the mean current value of the cells within each PA. Higher values of the current score indicated PAs that could facilitate the most movement across the network, and thus could potentially serve as critical stepping stones.
A second index, the conductance score was developed, by accounting the inverse of effective resistance values (i.e. conductance values). Since connectivity increases with the decrease of effective resistance, we created this index in order to detect PAs with the strongest connections. For each PA, we computed the conductance score as the mean conductance between the specific PA and the rest of the network. Higher values of conductance score would be indicative of the PAs that are better connected than others and therefore have a critical role for the connectivity of the network. We applied Spearman correlation coefficient, to explore for potential patterns of association between the two indices.
A graph theory assessment of connectivity properties
To investigate connectivity properties of the full network of PAs, we applied graph theory. For each species, a weighted, undirected network was created by using each PA as a node and considering the effective resistance as the edge weight. For any of these networks, we considered that the connection between any two sites was achieved if the effective resistance value was below a certain threshold. We evaluated these networks with a series of effective resistance thresholds, starting from a value of 1 up to the maximum obtained value (i.e. 165) with increasing steps of 1.
Network density (i.e. the number of network edges divided by the number of all possible network edges) was investigated, with networks of higher densities being more robust and thus offering higher connectivity quality between PAs. We examined if a network was connected by exploring whether there were paths between PAs which ensured that any PA was connected with at least another one. In the case of non-connected networks, the number of their components (i.e. the connected subnetworks of a network) and the giant component (i.e. the component with the biggest number of members) were identified. In addition, we calculated the effective mesh size (i.e. the summary of all components’ squared total area divided by total study area) (Jaeger 2000), which takes into account the PAs’ area, along with the number of components.
Even if the landscape poses minimum opposition to animal movement, connections among PAs might not be feasible due to species dispersal limitations. Therefore, we compared networks constructed based on effective resistance with networks based on geographical distance in order to identify potential differences on their efficiency. Networks based on geographical distance maintained connections among PAs if their geographical distance was equal or lower to the species dispersal capacity. The geographical distance based networks were compared with networks developed based on effective resistance with a threshold of 10. As resistance values were evaluated in a scale from 1 to 100, this threshold indicates relatively low resistance to movement.
Raster layers were constructed in ArcMap v10.4 Geographic Information Systems software of the Environmental Systems Research Institute (ESRI). The resistance surfaces, statistical analyses and development of graph models were conducted in R v3.4.0, using ‘raster’, ‘rgdal’, ‘gdistance’, ‘igraph’ and ‘ggplot2’ packages.
Results
The analyses demonstrated that current values of the landscape mosaic (Fig. 3) differed significantly among all species (Kruskall Wallis H Test, p < 0.05, in all cases; Dunn’s Test, p < 0.05, in all cases; Online Resource Table S5). Still, for every pair of species, we found that the obtained current values of landscape mosaic had a statistically significant positive relation (rs values ranged from 0.58 up to 0.91, p < 0.05, in all cases).

The current maps for Canis lupus (a), Capreolus capreolus (b), Vulpes Vulpes (c) and Ursus arctos (d). Only the protected areas that are suitable for each species are highlighted. Current value indicates predicted movement. Areas outside the study area are represented with light grey
The consensus map delineated corridors that facilitate movement for all studied species (Fig. 4), with four main corridors identified in the study area. The largest corridor was recognized in the center of the study area, crossing the national borders of three countries as expanding from the southwestern part of Bulgaria to the central part of Greece, spanning the biggest part of the Republic of North Macedonia. Another large corridor was detected crossing the central Bulgaria. This corridor mostly overlapped with the bulgarian National park of Tsentralen Balkan. Two additional smaller corridors were detected at the eastern part of Albania and at the wider area of Pindos Mountains in southwestern Greece.

The consensus current map. All protected areas that are suitable for at least one species are highlighted. Current value indicates predicted movement. Country borders are represented with a black line. Areas outside the study area are represented with light grey
Current and conductance score indices revealed some distinct patterns for the PAs’ connectivity (Online Resource Table S6–S7). Focusing on the PAs with high value of current score (i.e. top five, Online Resource Table S6 & Fig. S1) we found that species with high dispersal capacity (i.e. C. lupus and U. arctos) had two PΑs in common (Voras Mountain Peaks and Rodopi mountains, Greece), while the ones with low dispersal capacity (i.e. C. capreolus and V. vulpes) had four PAs (Mount Olympous National Park, Pierian Mountains and Vermio Mountain, Greece; Skrino, Bulgaria). Only one PA (i.e. Voras Mountain Peaks, Greece) was common for three species, while no PA was identified as common for all four species. Regarding the PAs with the highest conductance score (Fig. 5), three PAs were common for all species (i.e. Rodopi Mountains, Greece; Kotlenska planina–Provadiysko and Tsentralen Balkan, Bulgaria) (Online Resource Table S7). Comparing the PAs with the highest scores (i.e. top five) in both indices, we recognised one site for C. lupus (Rodopi Mountains, Greece) and two sites for U. arctos (Rodopi Mountains and Evros Delta, Greece) that were highly ranked by both indices, while no common, highly ranked site was identified for C. capreolus and V. vulpes.

Protected areas’ conductance score for Canis lupus (a), Capreolus capreolus (b), Vulpes Vulpes (c) and Ursus arctos (d). The colour of protected areas ranges from red to green representing low to high values of the conductance score respectively
For all species, we found a significant, but rather weak association between current score and conductance score (rs ranged from 0.50 up to 0.69, p < 0.05). A visual inspection of the relationship between current score and conductance score (Online Recourse Fig. S2) revealed that given areas with high current score do not necessarily have high values in both indices.
When studying connectivity properties of networks generated based on effective resistance across the landscape, we found some contradicting patterns for the four species. In all cases, networks developed for C. lupus and U. arctos had higher density (Online Resource Fig. S3), as well as the least number of components (Online Resource Fig. S4). In contrast, networks of PAs that could host C. capreolus and V. vulpes had higher effective mesh size values (Online Resource Fig. S5).
Our analyses on networks that were built based on geographical distance among PAs, demonstrated that only for the two species with high dispersal capacity (C. lupus and U. arctos) the networks supported a robust structure, with all PAs being interconnected. Both networks based on effective resistance (threshold of 10) and geographical distance were dense, with all PAs being included in a giant component. For the two species, effective mesh size had the same high values for both networks (Table 1).
For the other two species (C. capreolus and V. vulpes), networks based either on effective resistance (threshold of 10) or geographical distance were not connected. The networks based on effective resistance were relatively dense, with the giant component supporting more than 90% of PAs. Still, PAs outside the giant component formed numerous single–node components, as they were not connected with the other PAs. For C. capreolus and V. vulpes, the networks that were built based on geographical distance were highly fragmented (network density: 0.01 and 0.02 respectively, 48 and 42 components respectively). Effective mesh size values of the latter networks were relatively lower compared to the ones obtained by the network based on effective resistance (Table 1).
Discussion
Assessing connectivity patterns could be a complex and highly demanding task, as different species perceive the landscape in a different way reflecting their biological and behavioral needs (Brodie et al. 2015). Our analysis showed that the decisions taken during the process of modeling connectivity could greatly affect the relative importance of landscape elements. It is therefore imperative to identify and apply modeling protocols that could reduce the uncertainty that is derived from our limited knowledge on the species-landscape nexus.
In our study, areas detected as favorable for one species movement were often included within broader corridors selected for other species with higher dispersal abilities. One could suggest that this outcome promotes the application of the umbrella species perspective as a key framework for the development of connectivity networks (Roberge and Angelstam 2004; Baguette et al. 2013). Still, the quality of cells recognized for different species could differ; therefore, prioritizing landscape permeability based on larger dispersers might fail to recognize areas that are critical for low distance dispersers (Brodie et al. 2015). Our results are in line with such findings, as, despite similarities, the corridors identified as suitable for movement differ for each of the four studied species. For example, this is the case for the parts of southern Greece that were predicted to be used more for V. vulpes compared to C. capreolus, while they are unrelated to U. arctos’ movement (Fig. 3). As an alternative, several studies have tried to assess landscape quality based on species’ habitat preferences and use this information for delineating potential corridors (Abrahms et al. 2017). It is however likely that patches of low quality could serve as critical elements of the movement networks even for the same species (Almpanidou et al. 2014). Under these concerns, the application of modeling frameworks addressing potential needs of multiple species and applying multiple forcing scenarios seem as a promising way towards accounting for the uncertainty that accompany decisions related to connectivity analysis and assessment (Brodie et al. 2015).
Electric circuit theory is becoming very popular for connectivity analysis, as could offer advantages compared to other traditional tools such as least cost and graph theory methods (Correa Ayram et al. 2016). One of its main improvements is that circuit theory-based connectivity models allow the identification of multiple alternative paths among all patches, without being limited simply to assessments between pairs of nodes (e.g. PAs) or to a single least cost path (McRae et al. 2008). In order to generate robust predictions, circuit theory-based models are often accompanied by the application of different types of sensitivity analysis, by the means of using sets of resistance values (e.g. Stevens et al. 2006; Spear et al. 2010). Here, in an effort to provide additional evidence on landscape permeability, we aggregated information from expert opinion and developed two indices that allowed us not only to highlight the potential corridors, but also detect which PAs might have a critical role in overall connectivity. Our analyses indicated PAs (please see Online Resource Fig. S2) that despite their limited connection to the rest of the PAs (i.e. low conductance score), had high current score and thus high contribution to animal movement across the landscape. This finding indicates that less connected sites might also be critical for facilitating connectivity patterns, since they function as stepping stones. Therefore, PAs with a high value of either current or conductance score should be considered as important conservation targets. From a practical point of view, the information on the properties and contribution of each site to overall network connectivity could further be used for supporting systematic conservation planning as a sophisticated approach to derive the optimal decision based on sets of choices between alternative actions (McIntosh et al. 2017).
To the best of our knowledge, this is the first study that focused on the coherence and connectivity across the Balkan Peninsula, offering some insights on future conservation directions. Interestingly, the network of PAs in the region seemed to be rather well connected. Our results showed a robust structure of PAs for species with high dispersal abilities (C. lupus and U. arctos) with all sites being interconnected, with multiple alternative paths, indicating a network resilient to changes (e.g. climate change, land use change) and habitat alterations (e.g. local catastrophe). Such findings suggest that conservation plans and policies should not be site-based, as such dispersers do not limit their movement only to neighboring sites. Therefore, it becomes apparent that conservation policies that consider the specific biological and behavioral features of the species would be more efficient for such group of dispensers (Mazaris et al. 2013).
Still, we caution that for species with lower dispersal abilities (C. capreolus and V. vulpes) the network of PAs was not connected, with a number of sites remained isolated, which could likely jeopardize conservation capacity of the regional network of PAs. Such results are likely to indicate the inability of these species to disperse to new areas, raising serious concerns on the efficiency of the network structure to ensure population persistence and viability. The inclusion of new sites that could be used also as stepping stones, as well as the expansion of the borders of the current PAs area could contribute to the increase of network connectivity and efficiency (Mazaris et al. 2013).
The results of our analysis further demonstrated that the largest corridor spans beyond national boundaries, suggesting that transnational efforts are urgently required to foster conservation capacity. Indicative of this need is the fact that even though approximately one-fifth of the territory of the Balkan Peninsula is covered by PAs, countries as the Republic of North Macedonia have a small number of designated PAs, rendering the conservation network less effective (Radenković et al. 2017) especially as an extensive part of the largest corridor passes through its national boundaries. Under the same context, our analyses revealed that conservation efforts should not be limited to areas that are traditionally selected for the designation of PAs which are biased to higher elevations and isolated sites (Joppa and Pfaff 2009).
Several factors affect movement patterns with their influence being overwhelmed by either the properties of the landscape or individual variability. To somehow overcome limitations which inherent our knowledge on the perception and responses of species to landscape elements we developed a framework built upon expert opinion. We do not support that the use of expert knowledge could replace the wealth of information enclosed within data on actual movement and behavioral responses, but we acknowledge that their contribution could be valuable, especially at broad spatial scales that are subjected to major data gaps. In our case, even though some layers of information have been highlighted as important based on different sources of literature, we finally eliminate them following experts' knowledge on the specificities of the local populations and the study area. As the availability of field data is limited, expert opinion is and will likely continue to be a widely used method to evaluate the impact of landscape features to animal movement (Zeller et al. 2012; Correa Ayram et al. 2016).
Conclusion
In this paper we developed a method that can provide a valuable tool for assessing landscape connectivity at broad spatial scales, where scarce information on movement patterns exists. Measuring landscape connectivity for a set of species with different attributes allows for an extensive estimation of movement corridors that are meaningful for multiple species, instead of assessing a single species’ connectivity, where results would not necessarily represent other species. Therefore, these results could contribute in decision making for biodiversity management and planning of efficient PAs network.
The use of four focal species, the development of multiple layers of spatial information, the use of expert opinion and the combination of different methods for connectivity quantification provided for the first time a comprehensive assessment of connectivity status of the PAs network of the Balkan Peninsula. The identification of ecological corridors suitable for multiple species with different biological and behavioral features offers the potential of development of spatially specific conservation plans at transnational scale.
References
Abrahms B, Sawyer SC, Jordan NR, McNutt JW, Wilson AM, Brashares JS (2017) Does wildlife resource selection accurately inform corridor conservation? J Appl Ecol 54:412–422
Adriaensen F, Chardon J, De Blust G, Swinnen E, Villalba S, Gulinck H, Matthysen E (2003) The application of ‘least-cost’modelling as a functional landscape model. Landsc Urban Plan 64:233–247
Almpanidou V, Mazaris AD, Mertzanis Y, Avraam I, Antoniou I, Pantis JD, Sgardelis SP (2014) Providing insights on habitat connectivity for male brown bears: a combination of habitat suitability and landscape graph-based models. Ecol Model 286:37–44
Amano T, Sutherland WJ (2013) Four barriers to the global understanding of biodiversity conservation: wealth, language, geographical location and security. Proc R Soc B 280:20122649
Baguette M, Blanchet S, Legrand D, Stevens VM, Turlure C (2013) Individual dispersal, landscape connectivity and ecological networks. Biol Rev 88:310–326
Bănărescu PM (2004) Distribution pattern of the aquatic fauna of the Balkan Peninsula. In: Balkan biodiversity. Springer, pp 203–217
Benítez-López A, Alkemade R, Verweij PA (2010) The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol Conserv 143:1307–1316
Boitani L, Phillips M, Jhala Y (2018) Canis lupus. The IUCN Red List of Threatened Species 2018: e. T3746A119623865
Brodie J, Post E, Laurance WF (2012) Climate change and tropical biodiversity: a new focus. Trends Ecol Evol 27:145–150
Brodie JF, Giordano AJ, Dickson B, Hebblewhite M, Bernand H, Mohd-Azlan J, Anderson J, Ambu L (2015) Evaluating multispecies landscape connectivity in a threatened tropical mammal community. Conserv Biol 29:122–132
Christie MR, Knowles LL (2015) Habitat corridors facilitate genetic resilience irrespective of species dispersal abilities or population sizes. Evol Appl 8:454–463
Cooke SJ, Hinch SG, Wikelski M, Andrews RD, Kuchel LJ, Wolcott TG, Butler PJ (2004) Biotelemetry: a mechanistic approach to ecology. Trends Ecol Evol 19:334–343
Correa Ayram CA, Mendoza ME, Etter A, Salicrup DRP (2016) Habitat connectivity in biodiversity conservation: a review of recent studies and applications. Prog Phys Geogr 40:7–37
Cushman SA, McRae B, Adriaensen F, Beier P, Shirley M, Zeller K (2013) Biological corridors and connectivity. In: Macdonald DW, Willis KJ (eds) Key topics in conservation biology, vol 2. Wiley-Blackwell. Hoboken, NJ, pp 384–404
Dickson BG, Albano CM, Anantharaman R, Beier P, Fargione J, Graves TA, Gray ME, Hall KR, Lawler JJ, Leonard PB, Littlefield CE, McClure ML, Novembre J, Schloss CA, Schumaker NH, Shah VB, Theobald DM (2019) Circuit-theory applications to connectivity science and conservation. Conserv Biol 33:239–249
Dilkina B, Houtman R, Gomes CP, Montgomery CA, McKelvey KS, Kendrall K, Graves TA, Bernstein R, Schwartz MK (2017) Trade-offs and efficiencies in optimal budget-constrained multispecies corridor networks. Conserv Biol 31:192–202
Etherington TR (2016) Least-cost modelling and landscape ecology: concepts, applications, and opportunities. Curr Landsc Ecol Rep 1:40–53
Etherington TR, Holland EP (2013) Least-cost path length versus accumulated-cost as connectivity measures. Landsc Ecol 28:1223–1229
European Environment Agency - EEA (2012) Copernicus Land Monitoring Service, CORINE Land Cover (CLC, Version 20). Available at: https://land.copernicus.eu/pan-european/corine-land-cover/clc-2012. Accessed 20 January 2018
European Environment Agency - EEA (2016) Copernicus Land Monitoring Service, European Digital Elevation Model (EU-DEM, Version 1.1). Available at: http://land.copernicus.eu/pan-european/satellite-derived-products/eu-dem. Accessed 20 January 2018
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst 34:487–515
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin MP, Collins CD, Cook WM, Damschen EI, Ewers RM, Foster BL, Jenkins CN, King AJ, Laurance WF, Levey DJ, Margules CR, Melbourne BA, Nicholls AO, Orrock JL, Song D, Townshend JR (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv 1:e1500052
Hagen M, Kissling WD, Rasmussen C, De Aguiar MAM, Brown LE, Carstensen DW, Alves-Dos-Santos I, Dupont YL, Edwards FK, Genini J, Guimaraes Jn PR, Jenkins GB, Jordano P, Kaiser-Bunbury CN, Ledger ME, Maia KP, Darcie Marquitti FM, Mclaughlin O, Morrellato LC, O’Gorman EJ, Trojelsgaard K, Tylianakis JM, Vidal MM, Woodward G, Olesen JM (2012) Biodiversity, species interactions and ecological networks in a fragmented world. In: Advances in ecological research, vol 46. Elsevier, pp 89–210
Hebblewhite M, Haydon DT (2010) Distinguishing technology from biology: a critical review of the use of GPS telemetry data in ecology. Philos Trans R Soc B 365:2303–2312
Hoffmann M, Sillero-Zubiri C (2016) Vulpes vulpes. The IUCN Red List of Threatened Species. 2016: e. T23062A46190249
Jaeger JA (2000) Landscape division, splitting index, and effective mesh size: new measures of landscape fragmentation. Landsc Ecol 15:115–130
Jones KE, Bielby J, Cardillo M, Fritz SA, O’Dell J, Orme CL, Safi K, Sechrest W, Boakes EH, Carbone C, Connolly C, Cutts MJ, Foster JK, Grenyer R, Habib M, Plaster CA, Price SA, Rigby EA, Rist J, Teacher A, Bininda-Emonds ORP, Gittleman JL, Mace GM, Purvis A (2009) PanTHERIA: a species-level database of life history, ecology, and geography of extant and recently extinct mammals. Ecol Arch Ecol 90:2648–2648
Joppa LN, Pfaff A (2009) High and far: biases in the location of protected areas. PLoS ONE 4:e8273
Lovari S, Herrero J, Masseti M, Ambarli H, Lorenzini R, Giannatos G (2016) Capreolus capreolus. The IUCN Red List of Threatened Species 2016: e.T42395A22161386
Marrotte RR, Bowman J (2017) The relationship between least-cost and resistance distance PloS one 12:e0174212
Mazaris AD, Papanikolaou AD, Barbet-Massin M, Kallimanis AS, Jiguet F, Schmeller DS, Pantis JD (2013) Evaluating the connectivity of a protected areas’ network under the prism of global change: the efficiency of the European Natura 2000 network for four birds of prey. PLoS ONE 8:59640
McIntosh EJ, Pressey RL, Lloyd S, Smith RJ, Grenyer R (2017) The impact of systematic conservation planning. Annu Rev Environ Resour 42:677–697
McLellan B, Proctor M, Huber D, Michel S (2017) Ursus arctos (amended version of 2017 assessment). The IUCN Red List of Threatened Species 2017: e. T41688A121229971
McRae BH (2006) Isolation by resistance. Evolution 60:1551–1561
McRae BH, Dickson BG, Keitt TH, Shah VB (2008) Using circuit theory to model connectivity in ecology, evolution, and conservation. Ecology 89:2712–2724
Mimet A, Clauzel C, Foltête J-C (2016) Locating wildlife crossings for multispecies connectivity across linear infrastructures. Landsc Ecol 31:1955–1973
Nathan R, Getz WM, Revilla E, Holyoak M, Kadmon R, Saltz D, Smouse PE (2008) A movement ecology paradigm for unifying organismal movement research. Proc Natl Acad Sci 105:19052–19059
Pe’er G, Tsianou MA, Franz KW, Matsinos YG, Mazaris AD, Storch D, Kopsova L, Verboom J, Baguette M, Stevens VM, Henle K (2014) Toward better application of minimum area requirements in conservation planning. Biol Conserv 170:92–102
Pelletier D, Clark M, Anderson MG, Rayfield B, Wulder MA, Cardille JA (2014) Applying circuit theory for corridor expansion and management at regional scales: tiling, pinch points, and omnidirectional connectivity. PLoS ONE 9:e84135
Pimm SL, Raven P (2000) Biodiversity: extinction by numbers. Nature 403:843–845
Pinaud D, Claireau F, Leuchtmann M, Kerbiriou C (2018) Modelling landscape connectivity for greater horseshoe bat using an empirical quantification of resistance. J Appl Ecol 55:2600–2611
Radenković S, Schweiger O, Milić D, Harpke A, Vujić A (2017) Living on the edge: forecasting the trends in abundance and distribution of the largest hoverfly genus (Diptera: Syrphidae) on the Balkan Peninsula under future climate change. Biol Conserv 212:216–229
Richard Y, Armstrong DP (2010) Cost distance modelling of landscape connectivity and gap-crossing ability using radio-tracking data. J Appl Ecol 47:603–610
Riley SJ, DeGloria SD, Elliot R (1999) Index that quantifies topographic heterogeneity intermountain. J Sci 5:23–27
Roberge JM, Angelstam P (2004) Usefulness of the umbrella species concept as a conservation tool. Conserv Biol 18:76–85
Rondinini C, Di Marco M, Chiozza F, Santulli G, Baisero D, Visconti P, Hoffmann M, Schipper J, Stuart SN, Tognelli MF, Amori G, Falcucci A, Maiorano L, Boitani L (2011) Global habitat suitability models of terrestrial mammals. Philos Trans R Soc B 366:2633–2641
Sahraoui Y, Foltête J-C, Clauzel C (2017) A multi-species approach for assessing the impact of land-cover changes on landscape connectivity. Landsc Ecol 32:1819–1835
Santini L, Di Marco M, Visconti P, Baisero D, Boitani L, Rondinini C (2013) Ecological correlates of dispersal distance in terrestrial mammals. Hystrix 24:181–186
Santini L, Saura S, Rondinini C (2016) A composite network approach for assessing multi-species connectivity: an application to road defragmentation prioritisation. PLoS ONE 11:e0164794
Saura S, Bertzky B, Bastin L, Battistella L, Mandrici A, Dubois G (2018) Protected area connectivity: shortfalls in global targets and country-level priorities. Biol Conserv 219:53–67
Sawyer SC, Epps CW, Brashares JS (2011) Placing linkages among fragmented habitats: do least-cost models reflect how animals use landscapes? J Appl Ecol 48:668–678
Schiavina M, Freire S, MacManus K (2019): GHS population grid multitemporal (1975, 1990, 2000, 2015) R2019A. European Commission, Joint Research Centre (JRC). https://doi.org/10.2905/42e8be89-54ff-464e-be7b-bf9e64da5218 PID: http://data.europa.eu/89h/0c6b9751-a71f-4062-830b-43c9f432370f. Accessed 20 November 2019
Spear SF, Balkenhol N, Fortin MJ, McRae BH, Scribner K (2010) Use of resistance surfaces for landscape genetic studies: considerations for parameterization and analysis. Mol Ecol 19:3576–3591
Stevens VM, Verkenne C, Vandewoestijne S, Wesselingh RA, Baguette M (2006) Gene flow and functional connectivity in the natterjack toad. Mol Ecol 15:2333–2344
UNEP-WCMC, IUCN (2018) Protected Planet: The World Database on Protected Areas (WDPA). UNEP-WCMC and IUCN, Cambridge, UK, Available at: www.protectedplanet.net. Accessed 20 January 2018
UNEP-WCMC, IUCN, NGS (2018) Protected Planet Report 2018. UNEP-WCMC, IUCN and NGS. Cambridge, UK; Gland, Switzerland and Washington, DC, USA
van der Ree R, Smith DJ, Grilo C (2015) The ecological effects of linear infrastructure and traffic: challenges and opportunities of rapid global growth. In: Handbook of road ecology, pp 1–9
Webster MS, Marra PP, Haig SM, Bensch S, Holmes RT (2002) Links between worlds: unraveling migratory connectivity. Trends Ecol Evol 17:76–83
Weiss S, Ferrand N (2007) Phylogeography of southern European refugia. Springer, Berlin
Whitmee S, Orme CDL (2013) Predicting dispersal distance in mammals: a trait-based approach. J Anim Ecol 82:211–221
Zeller KA, McGarigal K, Cushman SA, Beier P, Vickers TW, Boyce WM (2017) Sensitivity of resource selection and connectivity models to landscape definition. Landsc Ecol 32:835–855
Zeller KA, McGarigal K, Whiteley AR (2012) Estimating landscape resistance to movement: a review. Landsc Ecol 27:777–797
Acknowledgements
The work of ADM was supported by Combine2Protect, a Project co-funded by the European Union and by National Funds of the Participating Countries under the IPA Cross-Border Programme “Greece – the Republic of North Macedonia 2014–2020. The views expressed in this publication do not necessarily reflect the views of the European Union, the participating countries and the Managing Authority. This research work was supported by the Hellenic Foundation for Research and Innovation (HFRI) under the HFRI PhD Fellowship grand (Fellowship Number: 1018). We would like to thank D. Bakaloudis, L. Georgiadis, D. Gioulatos, Μ. Hernando, G. Iliopoulos, Α. Karamanlidis, and G. Mertzanis for their assistance in background layer selection and evaluation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Petsas, P., Tsavdaridou, A.I. & Mazaris, A.D. A multispecies approach for assessing landscape connectivity in data-poor regions. Landscape Ecol 35, 561–576 (2020). https://doi.org/10.1007/s10980-020-00981-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-020-00981-2