
Abstract
Context
Forest landscapes at the boreal–temperate ecotone have been extensively altered. Reducing the gap between current and presettlement forest conditions through ecosystem-based forest management (EBFM) is thought to enhance ecological integrity. However, climate change may interfere with this goal and make these targets unrealistic.
Objectives
We evaluated the impacts of climate change on the ability of EBFM to reduce discrepancies between current and presettlement forest conditions in southeastern Canada.
Methods
We used early-land-survey data as well as projections from a forest landscape model (LANDIS-II) under four climate change scenarios and four management scenarios to evaluate future discrepancies between presettlement forest conditions and future forest landscapes.
Results
By triggering swift declines in most late-succession boreal conifer species biomass, climate change would greatly reduce the ability of forest management to reduce the gap with presettlement forest composition, especially under severe anthropogenic climate forcing. Scenarios assuming extensive clearcutting also favor aggressive competitor species that have already increased with high historical harvest levels (e.g., poplars, maples).
Conclusions
EBFM would still be the “less bad” forest harvesting strategy in order to mitigate composition discrepancies with the presettlement forests, though it is likely to fail under severe climate forcing. In this latter case, one might thus question the relevancy of using presettlement forest composition as a target for restoring degraded forest landscapes. As such, we advocate that managers should relax the centrality of the reference condition and focus on functional restoration rather than aiming at reducing the gaps with presettlement forest composition per se.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Over the past two centuries, forest landscapes in northeastern North America have been extensively modified by humans following European settlement (Cogbill et al. 2002; Nowacki and Abrams 2014). Extensive harvesting practices coupled with urbanization, agriculture and human-caused changes in wildfire activity have strongly disrupted the natural disturbance regime. Accordingly, present-day forest landscapes are quite different in age structure and tree species composition compared with pre-European settlement conditions (e.g., Boucher et al. 2009a, 2014; Dupuis et al. 2011; Danneyrolles et al. 2016a, b). For example, forests at the boreal-temperate ecotone were historically dominated by late-seral, uneven-aged stands (Boucher et al. 2009a). Extensive harvesting has considerably increased forest fragmentation and the proportion of young stands. Furthermore, widespread high-grading and diameter-limit cuts during the 19th century greatly reduced the abundance of large-diameter spruces and pine species (Boucher et al. 2009b). In addition, the extensive use of clearcut harvesting during the 20th century has reduced the abundance of several late-succession, slow-growing species (e.g., white cedar, spruces), while enhancing the recruitment of pioneer and/or fast-growing species (e.g., trembling aspen, sugar and red maples). Because pre-European forest conditions are viewed as a “bench mark” of the natural forest state of northeastern North America (Agee 2003), partially restoring, or at least “reducing the gap” between current and presettlement conditions is considered an important, long-term management objective to enhance ecological integrity (Sabogal et al. 2015).
Ecosystem-based forest management (EBFM) is a management philosophy that emphasizes the preservation of biodiversity and viability of ecosystems while maintaining sustainable timber supply. It may be achieved through reducing dissimilarities between current and presettlement forest conditions by maintaining the disturbance regime or the forest structure within its historical range of variability (Landres et al. 1999). Under this approach, different intensities of partial cutting, may be most appropriate for the boreal-temperate ecotone, since its historical, natural disturbance regime was dominated by small, gap-forming disturbances such as insect outbreaks, small patches of windthrow, and single-tree mortality events (Lorimer 1977; Frelich 2002). Increased diversity in forest composition and age structure at the boreal-temperate ecotone is considered to improve resilience by enhancing the ability of forest ecosystems to recover from or withstand disturbances (Duveneck et al. 2014). EBFM has recently been implemented in various regions of eastern Canada’s forests with the intent to reduce structural and compositional dissimilarities with presettlement forests (Vaillancourt et al. 2009).
Although EBFM may provide a reasonable approach to steer forests back to presettlement conditions, the projected global warming at the boreal-temperate ecotone is likely to drive further changes in forest age structure and composition (Strahan et al. 2016; Duveneck et al. 2017). For instance, increased anthropogenic climate forcing over the coming decades is projected to strongly alter forest composition and age structure at the boreal-temperate forest ecotone (e.g., Duveneck et al. 2014, Duveneck and Scheller 2015a; Boulanger et al. 2016, 2017; Taylor et al. 2017). More specifically, discrepancies in tree species’ physiological responses to climate change are likely to drive shifts in forest composition (e.g., Price et al. 2015; Reich et al. 2015) by enhancing the competitiveness of temperate tree species adapted to warmer conditions at the expense of cold-adapted boreal species. Concurrently, modification of insect outbreak regimes (e.g., Régnière et al. 2012) may cause increased recruitment of pioneer tree species over longer-lived “climax” species, and shift forest age structure to higher proportions of younger age classes. Furthermore, increases in the frequency of climate-driven disturbances (e.g., fire and windthrow) that create canopy openings may facilitate colonization by tree species that are currently at their northern climatic range limit, especially those capable of long-distance seed dispersal thus leading to future ecosystems with no current analogs (Stanturf et al. 2014). In light of these potential climate-driven changes, focusing efforts on restoring presettlement forest conditions may be unrealistic as future forest ecosystems may behave differently from those that characterized the presettlement era (Ravenscroft et al. 2010; Millar 2014). It is unknown to what extent forest management strategies, including EBFM, may be successful in reducing the dissimilarity between current and presettlement forests when considering climate change. If not, current forest management objectives involving restoring presettlement landscapes should be reconsidered.
Indeed, forest management strategies have the potential to influence how future forest landscapes will respond to climate change (e.g., Ravenscroft et al. 2010; Steenberg et al. 2013). For instance, management strategies can help maintain a desirable species even if climate conditions are not optimal, for example, by promoting its competitive ability through selective removal of competitors or by modulating light conditions (Landhäusser et al. 2010; Ravenscroft et al. 2010; Steenberg et al. 2013). Alternatively, harvesting and climate change may interact to yield unanticipated changes in forest landscapes (Steenberg et al. 2013). For example, partial cutting strategies may impede the northward migration of warm-adapted, pioneer species compared with even-aged management such as clearcutting, which creates larger openings that promote early-succession species (Steenberg et al. 2013). Strategies favoring structural and compositional diversity have been suggested to promote forest resilience under increasing climate warming within the temperate–boreal interface (Duveneck et al. 2014; Duveneck and Scheller 2015a). Interactions between harvesting and climate change need to be investigated to help define realistic conservation targets and forest management strategies (Millar et al. 2007).
In this study, we evaluated the impacts of climate change on the ability of EBFM to reduce the dissimilarity between current and presettlement forest composition within the boreal-temperate ecotone (Brandt et al. 2013) of southeastern Canada, a region that is projected to experience major changes in species composition before the end of the 21st century under rapid climate change (Steenberg et al. 2013; Duveneck et al. 2014; Duveneck and Scheller 2015a; Reich et al. 2015; Boulanger et al. 2016; Taylor et al. 2017). Specifically, we conducted spatially explicit forest landscape simulations in which we assessed the long-term impacts of contrasting harvesting strategies on forest composition, using different anthropogenic climate forcing scenarios. We estimated the relative importance of the simulated harvesting scenarios compared with climate change as drivers of discrepancies between presettlement and future forest composition. We expected that increased anthropogenic climate forcing would drive forest composition further away from presettlement conditions. We investigated whether EBFM-based scenarios would help attenuate these gaps by favoring late-seral species at the expense of aggressive pioneer species, which are favored by extensive past logging rates above presettlement levels.
Methods
Study area
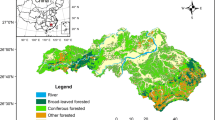
The 70,113 km2 study area (Fig. 1) covers most forested lands located south of the St. Lawrence River in the province of Quebec, Canada. It lies mainly within the northern extent of the Appalachians Mountains, a broadly rolling mosaic of upland plateaus, generally lying between 200 and 800 m above sea level (but reaching 1200 m in the easternmost part), and valleys covered by glacial till and humo-ferric podzols. Climate is typical of the humid continental zone with mild mean annual temperatures ranging from 0 to + 5 °C from north to south and precipitation ranging from 800 to 1000 mm from west to east. The vegetation is part of the “boreal-temperate ecotone” (Evans and Brown 2017), which transitions from northern hardwood forests to mixedwood and mainly coniferous southern boreal forests along a southwest to northeast gradient. This gradient encompasses four different bioclimatic domains according to the ecological land classification of Quebec (Robitaille and Saucier 1998), namely (i) Sugar maple—Basswood; (ii) Sugar maple—Yellow birch; (iii) Balsam fir—Yellow birch; and iv) Balsam fir—White birch. For this study, the first two bioclimatic domains were grouped into “northern hardwood” forest and the latter two into “mixedwood” forest. Unlike more northern boreal-type coniferous forests, these bioclimatic domains are characterized by rather small- to medium-scale natural disturbances. Indeed, recurrent spruce budworm outbreaks are the most important natural disturbance in the mixedwood forest region (Boulanger et al. 2012), while small windthrows and single-tree mortality mostly drive natural forest succession in the northern hardwood forest region. Wildfires can occur, but they are rather uncommon with intervals ranging between 400 and > 3000 years over the last two centuries (Bergeron et al. 2006; Payette et al. 2017).

Location of the study area (red) and of the forest regions and the 25 km2—cells (dark grey) within which both early land-survey and LANDIS outputs were considered for analyses. PEI Prince Edward Island
Materials and methods
Presettlement forest composition
The presettlement forest composition data used in this study were derived from historical land-surveys conducted between 1795 and 1940 for 10 square mile (approximately 16 × 16 km) townships located within the study area (Dupuis et al. 2011; Terrail et al. 2014; Danneyrolles et al. 2016b). Historical surveys were conducted systematically along the boundaries of the townships as well as along parallel “range lines”, at approximately 1.6 km intervals, subdividing the townships. Observations were georeferenced and classified as either linear or point data according to the surveyor’s spatial description. Linear segments were generally delineated by lot limits (about 260 m long) whereas point observations were frequently distributed at each lot corner (about 260 m) or every 10 imperial chains (about 200 m). For each observation point or line segment, surveyors described forest composition by listing taxa either at the species or genus level, according to their relative importance in the stand (Terrail et al. 2014). Only surveys conducted before 1900 were considered (except in the Gaspé peninsula where observations were made between 1900 and 1940, still before large-scale extensive industrial harvesting began) to avoid observations of forests affected by harvesting. Further details regarding the compilation of early land-surveys can be found in the original publications (Dupuis et al. 2011; Terrail et al. 2014).
Future forest composition
Climate data and future climate projections
Forest landscape changes were simulated for the 2000–2200 period using three different radiative forcing scenarios, known as Representative Concentration Pathways (RCP; van Vuuren et al. 2011), namely RCP 2.6, RCP 4.5 and RCP 8.5. For the period 2000–2010, data interpolated from climate station records by McKenney et al. (2013) were used to produce monthly time series of “current climate”. Future climate projections for 2011–2100 for each RCP, as simulated by the Canadian Earth System Model version 2 (CanESM2; Arora and Boer 2010), were downloaded from the World Climate Research Program (WCRP) Climate Model Intercomparison Project Phase 5 (CMIP5) archive. A bias correction was performed on the data from CanESM2 for 1961–2100 by expressing them as differences from (temperature) or ratios of (precipitation) the CanESM2 monthly means for the 1961–1990 period (McKenney et al. 2013). The bias-corrected data were then merged with observed 30-year monthly climate normals for 1961–1990, interpolated from climate station records by McKenney et al. (2013), to produce three RCP scenario climate projections for 2011–2100. The 2100–2200 period was an extension of the 2070–2100 conditions. According to the CanESM2 projections, mean annual temperature would increase by about 3.5 °C (RCP 2.6) to 7.5 °C (RCP 8.5) throughout the study area by 2100 (compared with circa 2000), while average precipitation is projected to vary little from current values, regardless of the forcing scenario (Suppl. Mat. S2).
The LANDIS-II forest landscape model
Future forest composition was simulated using LANDIS-II, a spatially-explicit raster-based model that simulates forest dynamics by integrating landscape-scale processes (e.g., disturbance and seed dispersal) as well as stand-level forest processes (e.g., succession; Scheller et al. 2007). In LANDIS-II, tree species are defined in each grid cell using unique life-history attributes and are represented as 10-year age cohorts. The great majority of common species occurring south of the study area are also observed in this study (Wilson et al. 2012; Wang et al. 2016; Taylor et al. 2017). That being said, species typical of the Carolinean forest such as Carya spp., Tilia americana and some Quercus spp. were not included as very few Canadian FIP including these species were available for calibration. We believe interactions with these potentially northward migrating species should be minimal and potentially restrained to the very southwestern tip of the study area considering their current occurrence in nearby regions (mainly New England, southwestern Quebec and Ontario). We set grid cell size to 250 m (6.25 ha). Forest community structure in each cell was initialized as present-day vegetation using forest attribute data derived from the Canadian National Forest Inventory (NFI) and age cohort data from Quebec’s provincial permanent and temporary forest inventory plots (FIP) as in Boulanger et al. (2016, 2017). It is important to note here that the current (2001) forest composition was used as initial conditions in the simulations, not the presettlement one.
Each grid cell was assigned to a landtype in which local soil (Mansuy et al. 2014) and climate conditions were assumed to be homogeneous across all cells of that landtype. Grid cells with less than 50% forest cover (by area, according to the inventories) were excluded from the simulations, leaving 845,108 “active” cells covering 52,819 km2. Landtype soil information included rooting depth, available water content, pH, and available nitrogen, and was obtained from Mansuy et al. (2014) and the Soil Landscapes of Canada (Centre for Land and Biological Resources Research 1996).
Forest succession and species growth potential
Forest succession was simulated using the LANDIS-II Biomass Succession extension v 3.1 (Scheller and Mladenoff 2004). This extension simulates changes in cohort biomass in each grid cell over time as each cohort regenerates, ages and dies. Biomass growth takes into account tree species’ life-history traits and species-specific responses to environmental conditions that are unique to each landtype. Species’ life-history traits information was collected from various sources (e.g., Burns and Honkala 1990) and several previous studies involving LANDIS-II for North American forest landscapes (Suppl. Mat. S3, Table 1).
We used PICUS (Lexer and Hönninger 2001) to derive three climate-sensitive dynamic growth parameters, i.e., species establishment probability (SEP), maximum aboveground NPP (maxANPP) and maximum aboveground biomass (maxAGB), as inputs to the LANDIS-II Biomass Succession model. PICUS, v. 1.5 (http://picus.boku.ac.at) is an individual tree-based, spatially explicit, forest gap model (Lexer and Hönninger 2001) that simulates the germination, establishment, growth, and mortality of individual trees on interacting 100 m2 dissimilarities or “patches” of forest area. PICUS runs on annual time steps and accounts for the effects of climate and soil properties on tree germination and growth (see Taylor et al. 2017 for a complete description). Mono-specific stands of each tree species on all landtypes were simulated with PICUS starting from bare-ground, using the species parameters given in Suppl. Mat. S3, Table 2. Monthly time series of climate data for each time period (2000–2010, 2011–2040, 2041–2070, 2071–2200) and each forcing scenario (baseline, RCP 2.6, RCP 4.5, RCP 8.5) were used to drive each simulation for 300 years. Dynamic growth parameters, as well as static growth and mortality curve shape parameters used in the Biomass Succession extension, were validated under baseline climate conditions by (i) assessing the realism of emerging successional pathways using pixel-scale simulations; and (ii) comparing species-specific biomasses at time t = 0 with the actual biomass estimated from NFI forest cover maps (Beaudoin et al. 2014; Boulanger et al. 2016). For further details on how these parameters were derived for LANDIS-II, see Suppl. Mat. S4. Mean values for each dynamic growth parameter for the mixedwood and northern hardwood forest region under each climate forcing scenario are presented in Suppl. Mat. S5. Trends in species-specific aboveground biomass as simulated by LANDIS-II under each climate forcing x harvesting scenario for the mixedwood and northern hardwood forest regions are presented in Suppl. Mat. S6.
Natural disturbances
Two natural disturbance agents, namely fire and outbreaks of spruce budworm (SBW, Choristoneura fumiferana [Clem.]), were considered in the LANDIS-II simulations. Fire simulations were conducted using the LANDIS-II Base Fire extension, which simulates stochastic fire events. Fire regime data (annual area burned, fire occurrence, and mean fire size) were summarized into “fire regions” corresponding to the intersection between the study region and the Canadian Homogeneous Fire Regime (HFR) zones of Boulanger et al. (2014). As in Boulanger et al. (2016), baseline and future fire regime parameters within each fire region were calibrated according to models developed by Boulanger et al. (2014) and further updated for different RCP scenarios (Gauthier et al. 2015). Boulanger et al. fire models were built in order to predict monthly annual area burned at the HFR zone level from various temperature and precipitation variables, notably related to the Canadian Forest Fire Weather Index System (Van Wagner 1987).
Outbreaks of SBW were simulated using the Biological Disturbance Agent (BDA) extension v3.0 (Sturtevant et al. 2004), which simulates host tree mortality following insect outbreaks. Host tree species for SBW were, from the most to least vulnerable, balsam fir (Abies balsamea), and white (Picea glauca), red (P. rubens) and black (P. mariana) spruce. Outbreaks were simulated as probabilistic events at the grid-cell level with probabilities being a function of site and neighborhood resource dominance (e.g., host tree abundance within a 1-km radius) as well as regional outbreak status. Parameters used in this study were calibrated and validated using various sources for the mixedwood forest (e.g., Hennigar et al. 2008). Regional outbreaks were calibrated at the highest severity level possible and were set to last, at most, one time step (10 years) and to occur every 40 years in accordance with typical observed regional recurrence cycles (Boulanger et al. 2012).
Harvesting
Forest harvesting was simulated using the Biomass Harvest extension (v3.0; Gustafson et al. 2000). Four harvesting scenarios were simulated according to a gradient of harvesting pressure, from no harvesting, to EBFM, to standard clearcutting [CC], to very high clearcutting [VHCC] (Table 1). Altogether, these simulated harvesting scenarios represent the “harvesting envelope” from which most of the potential outcomes related to harvesting impacts can be expected. EBFM was emulated by implementing different intensities of clearcutting and partial harvesting; harvesting methods were set to vary according to potential vegetations as defined by Quebec’s Hierarchical System for Territorial Ecological Classification (Bergeron et al. 1992). This system classifies forest stands according to their potential natural vegetation type, which is a function of climatological and geomorphological constraints on vegetation growth and succession. As a result, the EBFM scenario should closely mimic the historical disturbance regimes (e.g., spruce budworm outbreaks, single-tree mortality, small gap openings) of both forest regions. Potential vegetation types were grouped into six generalized groups in which harvesting methods were as presented in Table 1. Rotation length time and biomass removal levels were fixed according to current harvesting regulations and expert advice. Although unrealistic for the upcoming decades, clearcutting with rates of 2% per year (VHCC) was also considered as this scenario as it was applied extensively during the post-settlement period (Boucher et al. 2009a). When a harvesting event occurred in a grid cell, all species were assumed to be harvested at the same rate. Harvest was performed by “management areas”, i.e., either by forest management units for public lands or by ecodistricts for private lands. Total biomass harvested following each harvesting scenario under each climate scenario is presented in Suppl. Mat. S7. No salvage logging was simulated. During a given timestep, harvesting was simulated first, then fire, then SBW.
Experimental design and simulations
LANDIS simulations were run according to a full factorial design with climate scenarios and harvesting prescriptions as factors. The four climate scenarios were: baseline, RCP 2.6, RCP 4.5 and RCP 8.5, while there were four levels of harvesting prescriptions (No harvest, EBFM, CC and VHCC). Five replicates were run for each cross-factor level for a total of 80 simulations (4 climate scenarios x 4 harvesting strategies x 5 replicates). All simulations were run for 200 years at 10-year time steps starting in year 2000. Climate-sensitive parameters (fire regime; growth [maxANPP, maxAGB and SEP]) were allowed to change in 2010, 2040 and 2070 according to the forcing scenario and were held fixed thereafter for the 2070-2200 period. Parameters calibrated for the baseline climate were used for the 2000-2010 period for all simulations as well as for the spin-up phase, when initializing tree species biomass.
Data analyses
For each taxon, we derived a relative abundance (RA) index, which expresses the relative abundance of each taxon in both the early land-survey records and the LANDIS outputs. Rank was used as a variable since it represents the smallest common denominator between early land-survey and LANDIS outputs. For each taxon, RA was calculated for a given observation (early land-survey) or cells (LANDIS outputs) based on the rank occupied by the taxon. For early land-survey data, the order in which taxa were enumerated by the surveyors was considered as a proxy of the relative basal area of this taxon, with the first enumerated taxon considered dominant at that location (Terrail et al. 2014). Taxa were then ranked according to their RA values as follows: 1st rank = 4, 2nd rank = 3, 3rd rank = 2, 4th rank = 1. Any other taxa included in the list of observed species were assigned zero RA. For LANDIS data, AGB of some species (spruces [Picea spp.], maples [Acer spp.], pines [Pinus spp.], poplars [Populus spp.]) was summed at the genus level to match the taxa observed by early surveyors. Each taxon’s AGB were translated into BA using random forest (RF) models (See Suppl. Mat. S8 for more details). Taxa were then ranked in each cell according to their predicted BA.
As observations from early land-surveys are not evenly distributed over the study area, we restrained the spatial coverage of the LANDIS data analyses to areas with sufficient historical observations. To do so, we divided the study area into 25 km2 (5 × 5 km) cells (Fig. 1). Only historical forest data and LANDIS output data in 25 km2 cells with at least four historical observations were retained in the final analyses.
Comparisons between presettlement and future forest composition were conducted at the forest region level for (i) forest composition as a whole and (ii) each taxon separately. First, RA indices were computed for each forest region and for each taxon from the historical dataset and from LANDIS simulations at time t. The RA index for a given taxon was assessed as the average RA value over all locations (LANDIS cells or early land-survey observations). Then, the Bray–Curtis dissimilarity metric was used to estimate pairwise dissimilarities in the community matrices between the LANDIS output data at time t and the presettlement dataset. Community matrices were built using the RA indices of each taxon within each forest region for a given set of simulations at time t. As variations among replicates were very small, community matrices from LANDIS outputs were built from the average of the five replicates. Variation through time in dissimilarities between presettlement and projected forests were then examined for each forest region by contrasting harvesting prescriptions for each climate scenario separately. In addition to analyses of forest composition per se, similar analyses of temporal trends were conducted for each taxon separately, to assess the level to which taxa would be affected by harvesting and climate change scenarios. This was achieved by comparing taxon-specific RA between LANDIS outputs and presettlement forests according to the same experimental scheme as the one just described for forest composition.
We quantified the relative importance of harvesting scenario and climate change impacts on the discrepancy between projected and presettlement forest landscapes, considering only EBFM and CC to limit analyses to the most “realistic” management scenarios. Following a two-way factorial PERMANOVA (Anderson 2001), where harvesting scenario (2 levels) and climate change (2 levels: baseline and the given RCP scenario) were each considered as a factor, we calculated the partial R-squared for harvesting and climate change. We used Bray–Curtis dissimilarities to calculate pairwise distances in the PERMANOVA results. Each community matrix was determined based on the RA discrepancy of each species within the forest region for a given simulation at time t. Analyses were performed separately for each RCP scenario and for each forest region. PERMANOVA tests were run with the adonis function of the vegan package v2.4-2 (Oksanen et al. 2017) in R 3.3.0 (R Core Team 2016)
Results
Bray–Curtis dissimilarity in relative abundance (RA) generally increased with increasing harvesting pressure (no harvesting < EBFM < CC < VHCC) in both forest regions, especially under RCP 8.5 (Fig. 2). When the “no harvesting” scenario was excluded, the EBFM harvesting scenario had the strongest tendency to stabilize or impede increases in dissimilarity between projected and presettlement forests. In contrast, Bray–Curtis dissimilarity strongly increased under both clearcut harvesting scenarios. Under baseline, RCP 2.6 and RCP 4.5 climate scenarios, dissimilarity between projected and presettlement forest slightly increased over time regardless of the harvesting scenario. Under RCP 8.5, however, dissimilarity sharply increased after 2100, especially for the CC and VHCC harvesting strategies. Dissimilarity with presettlement forests increased the least under the no harvest scenario, regardless of climate forcing and forest regions (Fig. 2).

Temporal trends in Bray–Curtis dissimilarity in relative abundance between projected and presettlement forests for a the mixedwood and b the northern hardwood forest region
Anthropogenic climate forcing mostly impacted balsam fir, beech, maples and spruce RA (Fig. 3). Increased climate forcing strongly boosted maple and beech RA, especially in the northern hardwood forest region, while RA of balsam fir and spruces declined (especially after 2100) relative to presettlement conditions. In contrast to clearcutting, EBFM harvesting or no harvesting would impede an increase in balsam fir RA discrepancies between projected and presettlement forest landscapes under baseline or mild anthropogenic climate forcing (RCP 2.6 and RCP 4.5), while under RCP 8.5, balsam fir would collapse under all harvest scenarios. Except for white birch in the mixedwood forest region, harvesting scenarios had rather similar impacts among taxa whereas partial harvesting and no harvesting limited the increase in RA differences between presettlement and projected forests (Fig. 3). Both clearcut scenarios decreased RA of white cedar, thereby reducing its abundance deficit relative to presettlement conditions, regardless of the climate scenario. The simulations suggested maples would remain above presettlement levels in both forest regions, regardless of the climate or logging scenario; poplars would be similarly enhanced in the mixedwood forest region. In the northern hardwood forest, only the no-harvest and EBFM logging scenarios permitted the slow recovery of poplar RA, regardless of climate forcing. Yellow birch remained below presettlement levels, except under RCP 8.5 in the mixedwood forest region. Only small temporal variations in RA were projected for pines (Fig. 3).

Difference in relative abundance (assessed as mean difference in taxa rank) between projected and presettlement forest landscapes for each taxon in a the mixedwood and b the northern hardwood forest region. Positive values indicates higher relative abundance in projected landscapes relative to presettlement conditions
Importance of harvesting vs climate scenarios
In both forest regions, harvesting scenarios explained much of the variation in RA discrepancies (ω2 > 0.70) throughout the simulation period under mild (RCP 2.6) and moderate (RCP 4.5) anthropogenic climate forcing. Under RCP 8.5, harvesting scenarios remained important up to 2100, after which climate-induced changes became more important (Fig. 4).

Trends in partial R2 values for climate change (yellow) and harvesting scenario (blue) for forest vegetation under RCP 2.6, RCP 4.5 or RCP 8.5 forcing scenarios
Discussion
Climate change impacts on future forest landscapes
Our results project that increased anthropogenic climate forcing will trigger substantial declines in most boreal conifer species biomass, producing strong deficits compared with presettlement conditions. Such declines within the temperate-boreal transition zone have been reported in other modeling studies (e.g., Scheller and Mladenoff 2008; Ravenscroft et al. 2010; Steenberg et al. 2013; Duveneck and Scheller 2015a; Boulanger et al. 2016; Taylor et al. 2017). Field studies have shown that boreal species growing at the southern edge of their present-day distribution are likely to be vulnerable to warmer and drier climates (Huang et al. 2013; Fisichelli et al. 2014; Reich et al. 2015). Previous studies (e.g., Girardin et al. 2015) have shown that the projected warming, particularly under the RCP 8.5 forcing scenario, would impose strong growth constraints, likely sufficient to eradicate several boreal species (notably balsam fir, white and black spruces) in some regions.
Such climate-induced declines in boreal species will likely benefit co-occurring temperate species (notably beech and maples). Further improved growth potential of temperate species will provide them with a competitive advantage over boreal species (Reich et al. 2015; Boulanger et al. 2016; Taylor et al. 2017). Large increases in maple abundance under a warming climate would worsen an already large gap with presettlement conditions as this taxon is already considered overabundant in both forest regions (Brisson and Bouchard 2003; Dupuis et al. 2011). That being said, it is possible that other more southerly species typical of the Carolinean forest (e.g., Carya spp., Tilia americana, some Quercus spp.) and that were not included in this study, might reach the very southwestern tip of the study area and locally become common along with maple and beech. As such, the future importance of these two taxa might be locally overestimated. Poplars have also extensively increased since presettlement times (Dupuis et al. 2011; Boucher et al. 2017), and seem likely to maintain greater abundance than during presettlement conditions in the mixedwood forest region, presumably because of climate-induced increased in fire activity (Suppl. Mat. S2; Boulanger et al. 2017). Altogether, the climate change-induced decline of coniferous boreal species and the concomitant rise in deciduous taxa (both temperate and boreal) will amplify the already ongoing trend of increasing discrepancy with the largely conifer-dominated presettlement forest landscapes in this area (Dupuis et al. 2011; Danneyrolles et al. 2016a, b). Thus, our simulations suggest that future climate-induced changes in forest composition would add to the large changes that have already occurred since preindustrial times and are likely to threaten multiple ecosystem processes and the provision of goods and services, regardless of the harvesting scenario.
Harvesting impacts in the context of climate change
In our simulations, harvesting scenarios strongly dictated how discrepancies with presettlement forest composition would evolve under increasing anthropogenic climate forcing. Post-disturbance recovery and succession pathways were shown to be strongly affected by climate change (Fisichelli et al. 2014). Notably, high rates of stand-replacing disturbance are known to decrease forest landscape resistance to climate change by creating environmental conditions favoring, and thus accelerating, the colonization of pioneer species, including invaders already adapted to a warmer climate (Landhäusser et al. 2010). Similarly, our results show that clearcutting-based scenarios, especially under severe climate forcing, increased the discrepancies with presettlement landscapes by favoring pioneer species (e.g., poplars) and those producing large seedbanks (e.g., balsam fir, mostly under mild forcing) and by accelerating the proliferation of temperate deciduous species (beech and maples). In fact, high clearcutting rates combined with increased climate forcing would promote species that have already increased with high historical harvesting levels (Dupuis et al. 2011; Boucher et al. 2017). Scenarios assuming extensive clearcutting would also favor species that are outside their optimal climate tolerance (Landhäusser et al. 2010; Steenberg et al. 2013). For instance, despite lower growth potential under RCP 8.5 (Suppl. Mat. S4), our simulations show that future poplar abundance will remain higher than presettlement levels in the northern hardwood forest region following clearcutting. It is likely that widespread climate-induced mortality in boreal conifers, along with harvesting-induced vegetative regrowth and less shading in post-harvest conditions, will benefit poplars under severe climate forcing and high clearcutting rates. This emphasizes that forest management strategies involving disturbance rates and severity that are beyond the present range of variability (e.g., as caused by frequent clearcutting) are likely inappropriate to reduce discrepancy with presettlement conditions in these highly degraded forest regions.
How EBFM fares in the context of climate change?
Forest management strategies that exacerbate the deleterious impacts of historical extensive harvesting, including those that worsen the impacts of climate change, must be identified and abated while strategies enhancing forest resilience should be promoted (Millar et al. 2007; Yousefpour et al. 2014). Our results suggest that EBFM would be the “less bad” forest harvesting strategy in order to mitigate forest composition discrepancies with presettlement forests, though it is likely to fail under severe climate forcing. However, reducing the gaps with presettlement forest composition could still fail for some important taxa, such as yellow birch and white cedar, even under the simulated EBFM scenarios, since more aggressive competitors, such as poplars and maples, would likely increase in abundance, worsening the dissimilarities with presettlement landscapes. It is even possible that past harvesting practices have exceeded the “ecosystem inertia” or resilience of the presettlement forest landscapes, potentially creating a new “equilibrium” (a.k.a. “alternative stable state”) from which a return to the presettlement state may be very difficult to reach (Perring et al. 2016). In such a situation, restoration failures should be expected even under the “no harvest/no anthropogenic climate forcing” scenario. Actually, despite rather strong discrepancies in forest landscape trajectories among the different harvesting scenarios, none would seem to allow a full or even partial restoration of presettlement forest composition over the next 200 years, regardless of climate conditions.
Is sensu stricto restoration of presettlement composition a realistic target with climate change?
We have shown that the maintenance of presettlement forest composition is unlikely under severe climate forcing, regardless of the harvesting scenario. Actually, climate change-induced impacts would overwhelm future harvesting-induced impacts beyond 2100 under RCP 8.5 (e.g., see Figs, 2 and 4). Impacts of climate change have been projected to outweigh harvesting impacts at the boreal–temperate ecotone in eastern North America in the near future (Ravenscroft et al. 2010; Duveneck and Scheller 2015a; Boulanger et al. 2017). Previous studies have suggested that under high forcing scenarios, most harvesting scenarios, even those aimed at increasing resilience/resistance of the forest cover to climate impacts, would not be effective in preserving forest composition (e.g., Duveneck et al. 2014; Duveneck and Scheller 2015a). Hence, one might question the relevancy of using presettlement forest composition as a target for restoring degraded forest landscapes (e.g., Millar 2014; Strahan et al. 2016). Notably, Millar (2014) have suggested using presettlement forest composition as a suitable reference condition would be inappropriate if future climate conditions and disturbance regime are projected to change beyond the historical range of variability. Hence, strategies aimed at reducing the gaps with presettlement forest composition could prove to be unsuitable (Falk 2017).
Assessing dissimilarities with presettlement forests helps understand how forests have been affected by harvesting practices and how continuing or changing these practices might affect future forest resilience. Extensive clearcut practices over the last 200 years have strongly favored opportunistic deciduous species (notably, poplar and maples) over several late-successional conifer species (Boucher et al. 2009a; Dupuis et al. 2011). Our simulations suggest that with increasing radiative forcing, such trends would continue, most notably for maples particularly red maple, which is regarded as a “super generalist” (Abrams 1998). Maples have responded positively to anthropogenic disturbances in North American northern hardwood and mixedwood forests (Cogbill et al. 2002; Duchesne and Ouimet 2008; Nowacki and Abrams 2014); comparisons with presettlement forests and simulations help to understand the potential risk associated with these opportunistic invasive species and management strategies that favor them under increasing climate forcing should therefore be avoided.
Management implications
According to our results, EBFM might appear as a good strategy with potential to mitigate the negative impacts of climate change on the forestry sector in eastern Canada, while minimizing the risks associated with uncertainty in climate change scenarios. Considering their potential inability to recover historical forest conditions, it was proposed that future forest ecosystems should aim to be most compatible with their inherent capacities, keeping in mind a range of options based on social and ecological goals (Millar 2014; Falk 2017; Yousefpour et al. 2017). Managers should relax the centrality of the reference condition (Falk 2017) and focus on functional and diversity restoration at various spatial scales rather than aiming at reducing the gaps with presettlement forest composition per se (Dumroese et al. 2015; Duveneck and Scheller 2015b). Such a strategy might help to enhance forest resilience under climate-induced changes in tree growth and disturbance rates (Duveneck and Scheller 2015b). With severe climate change, our results suggest that partial harvesting scenarios are likely to slow down, but cannot ultimately prevent, abrupt climate-induced transitions of forest composition. Furthermore, best strategies must be aimed at mitigating the potential decline in forest productivity/biomass in order to preserve the ability of forest ecosystems to provide sustainable timber supply while maintaining sufficient cover for the provision of other ecosystem goods and services (Dumroese et al. 2015; Yousefpour et al. 2017). Management strategies could target other presettlement attributes that are deemed important for specific goals, e.g., age structure, structural and compositional diversity at the stand and landscape levels for biodiversity conservation. Supplementary analyses (see Suppl. Mat. S9) revealed that partial harvesting scenarios can mitigate climate-induced decline in total biomass in the long-term, even under severe climate forcing. Moreover, these strategies also appeared to be the most effective in maintaining the recovery of old-growth forests despite considerable uncertainties in future climate scenarios (Suppl. Mat. S9).
Conclusion
We demonstrated that climate change is likely to compromise the ability of forest management to reduce the gaps with presettlement forest composition targets at the boreal–temperate ecotone. However, considering that EBFM may help retain presettlement forest age structure and ease transition toward new forest ecosystems, it may represent an adaptive approach to maintain desirable societal and resource objectives (see Suppl. Mat. 7; Dumroese et al. 2015). Such a strategy should be regarded in a holistic approach in which other complementary actions (e.g., assisted migration; Pedlar et al. 2012; Dumroese et al. 2015; Hof et al. 2017) could be undertaken to ease climate-induced transitions in forest landscapes. Furthermore, as our collective understanding of future impacts of climate change on forest state increases, adaptive management is likely to play an increasing role to ensure that long-term management objectives are met (Yousefpour et al. 2017). In this regard, our study provides insights on potential management options that should help decision-makers to adapt in the face of climate change.
References
Abrams MD (1998) The red maple paradox: what explains the widespread expansion of red maple in eastern forests? Bioscience 48:355–364
Agee JK (2003) Historical range of variability in eastern Cascades forests, Washington, USA. Landscape Ecol 18:725–740
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austr Ecol 26:32–46
Arora VK, Boer GJ (2010) Uncertainties in the 20th century carbon budget associated with land use change. Glob Change Biol 16:3327–3348
Beaudoin A, Bernier PY, Guindon L, Villemaire P, Guo XJ, Stinson G, Bergeron T, Magnussen S, Hall RJ (2014) Mapping attributes of Canada’s forests at moderate resolution through kNN and MODIS imagery. Can J For Res 44:521–532
Bergeron JF, Saucier JP, Robert D, Robitalle A (1992) Québec forest ecological classification program. For Chron 68:53–63
Bergeron Y, Cyr D, Drever CR, Flannigan M, Gauthier S, Kneeshaw D, Lauzon E, Leduc A, Le Goff H, Lesieur D, Logan K (2006) Past, current, and future fire frequencies in Quebec’s commercial forests: implications for the cumulative effects of harvesting and fire on age-class structure and natural disturbance-based management. Can J For Res 36:2737–2744
Boucher Y, Arseneault D, Sirois L (2009a) Logging history (1820–2000) of a heavily exploited southern boreal forest landscape: insights from sunken logs and forestry maps. For Ecol Manag 258:1359–1368
Boucher Y, Arseneault D, Sirois L, Blais L (2009b) Logging pattern and landscape changes over the last century at the boreal and deciduous forest transition in Eastern Canada. Lands Ecol 24:171–184
Boucher Y, Grondin P, Auger I (2014) Land use history (1840–2005) and physiography as determinants of southern boreal forests. Landscape Ecol 29:437–450
Boucher Y, Perrault-Hébert M, Fournier R, Drapeau P, Auger I (2017) Cumulative patterns of logging and fire (1940–2009): consequences on the structure of the eastern Canadian boreal forest. Landscape Ecol 32:361–375
Boulanger Y, Arseneault D, Morin H, Jardon Y, Bertrand P, Dagneau C (2012) Dendrochronological reconstruction of spruce budworm (Choristoneura fumiferana Clem.) outbreaks in southern Québec for the last 400 years. Can J For Res 42:1264–1276
Boulanger Y, Gauthier S, Burton PJ (2014) A refinement of models projecting future Canadian fire regimes using homogeneous fire regime zones. Can J For Res 44:365–376
Boulanger Y, Taylor A, Price DT, Cyr D, McGarrigle E, Rammer W, Sainte-Marie G, Beaudoin A, Guindon L, Mansuy N (2016) Climate change impacts on forest landscapes along the Canadian southern boreal forest transition zone. Landscape Ecol 32:1415–1431
Boulanger Y, Taylor A, Price DT, Cyr D, Sainte-Marie G (2017) Stand-level drivers most important in determining boreal forest response to climate change. J Ecol 106:977–990
Brandt JP, Flannigan MD, Maynard DG, Thompson ID, Volney WJA (2013) An introduction to Canada’s boreal zone: ecosystem processes, health, sustainability, and environmental issues. Environ Rev 21:207–226
Brisson J, Bouchard A (2003) In the past two centuries, human activities have caused major changes in the tree species composition of southern Québec, Canada. Écoscience 10:236–246
Burns RM, Honkala BH (1990) Silvics of North America: 1. Conifers; 2. Hardwoods. Agriculture Handbook 654. U.S. Department of Agriculture, Forest Service, Washington, DC
Centre for Land and Biological Resources Research (1996) Soil Landscapes of Canada, v.2.2. Research Branch, Agriculture and Agri-Food Canada, Ottawa
Cogbill CV, Burk J, Motzkin G (2002) The forests of presettlement New England, USA: spatial and compositional patterns based on town proprietor surveys. J Biogeogr 29:1279–1304
Danneyrolles V, Arseneault D, Bergeron Y (2016a) Pre-industrial landscape composition patterns and post-industrial changes at the temperate–boreal forest interface in western Quebec, Canada. J Veg Sci 27:470–481
Danneyrolles V, Arseneault D, Bergeron Y (2016b) Long-term compositional changes following partial disturbance revealed by the resurvey of logging concession limits in the northern temperate forest of eastern Canada. Can J For Res 46:943–949
Duchesne L, Ouimet R (2008) Popultion dynamics of tree species in southern Quebec, Canada. For Ecol Manag 255:3001–3012
Dumroese RK, Williams MI, Stanturf JA, St Clair JB (2015) Considerations for restoring temperate forests of tomorrow: forest restoration, assisted migration, and bioengineering. New Forest 46:947–964
Dupuis S, Arseneault D, Sirois L (2011) Change from presettlement to present-day forest composition reconstructed from early land-survey records in eastern Quebec, Canada. J Veg Sci 22:564–575
Duveneck MJ, Scheller RM (2015a) Measuring and managing resistance and resilience under climate change in northern Great Lake forests (USA). Landscape Ecol. https://doi.org/10.1007/s10980-015-0273-6
Duveneck MJ, Scheller RM (2015b) Climate-suitable planting as a strategy for maintaining forest productivity and functional diversity. Ecol Appl 25:1653–1668
Duveneck MJ, Scheller RM, White MA, Handler SD, Ravenscroft C (2014) Climate change effects on northern Great Lake (USA) forests: a case for preserving diversity. Ecosphere 5:23
Duveneck MJ, Thompson JR, Gustafson EJ, Liang Y, de Bruijn AMG (2017) Recovery dynamics and climate change effects to future New England forests. Landscape Ecol 32:1385–1397
Evans P, Brown CD (2017) The boreal-temperate forest ecotone response to climate change. Environ Rev 25:423–431
Falk DA (2017) Restoration ecology, resilience, and the axes of change. Ann Miss Bot Garden 102:201–216
Fisichelli NA, Frelich LE, Reich PB (2014) Temperate tree expansion into adjacent boreal forest patches facilitated by warmer temperatures. Ecography 37:152–161
Frelich LE (2002) Forest dynamics and disturbance regimes. Studies from temperate evergreen-deciduous forests. Cambridge University Press, Cambridge
Gauthier S, Bernier PY, Boulanger Y, Guo J, Guindon L, Beaudoin A, Boucher D (2015) Vulnerability of timber supply to projected changes in fire regime in Canada’s managed forests. Can J For Res 45:1439–1447
Girardin MP, Hogg EH, Bernier PY, Kurz WA, Guo X, Cyr G (2015) Negative impacts of high temperatures on growth of black spruce forests intensify with the anticipated climate warming. Glob Change Biol 22:627–643
Gustafson EJ, Shifley SR, Mladenoff DJ, Nimerfro KK, He HS (2000) Spatial simulation of forest succession and timber harvesting using LANDIS. Can J For Res 30:32–43
Hennigar CR, MacLean DA, Quiring DT, Kershaw JA Jr (2008) Differences in spruce budworm defoliation among balsam fir and white, red, and black spruce. For Sci 54:158–166
Hof AR, Dymond CC, Mladenoff DJ (2017) Climate change mitigation through adaptation: the effectiveness of forest diversification by novel tree planting regimes. Ecosphere 8:e01981
Huang JG, Bergeron Y, Berninger F, Zhai L, Tardif JC, Denneler B (2013) Impact of future climate on radial growth of four major boreal tree species in the eastern Canadian boreal forest. PLoS ONE 8:e56758
Landhäusser SM, Wan X, Lieffers VJ, Chow PS (2010) Nitrate stimulates root suckering in trembling aspen (Populus tremuloides). Can J For Res 40:1962–1969
Landres PB, Morgan P, Swanson FJ (1999) Overview of the use of natural variability concepts in managing ecological systems. Ecol Appl 9:1179–1188
Lexer MJ, Hönninger K (2001) A modified 3D-patch model for spatially explicit simulation of vegetation composition in heterogeneous landscapes. For Ecol Manag 144:43–65
Lorimer CG (1977) The presettlement forest and natural disturbance cycle of northeastern Maine. Ecology 58:139–148
Mansuy N, Thiffault E, Paré D, Bernier P, Guindon L, Villemaire P, Poirier V, Beaudoin A (2014) Digital mapping of soil properties in Canadian managed forests at 250 m of resolution using the k-nearest neighbor method. Geoderma 235–236:59–73
McKenney D, Pedlar J, Hutchinson M, Papadopol P, Lawrence K, Campbell K, Milewska E, Hopkinson RF, Price D (2013) Spatial climate models for Canada’s forestry community. For Chron 89:659–663
Millar CI (2014) Historic variability: informing restoration strategies, not prescribing targets. J Sustain For 33:S28–S42
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and forests of the future: managing in the face of uncertainty. Ecol Appl 17:2145–2151
Nowacki GJ, Abrams MD (2014) Is climate an important driver of post-European vegetation change in the Eastern United States? Glob Change Biol 21:314–334
Oksanen J, Blanchette GF, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2017) vegan: Community Ecology package v 2.5-2. https://CRAN.R-project.org/package=vegan. Accessed 18 Dec 2018
Payette S, Pilon V, Couillard PL, Laflamme J (2017) Fire history of Appalachian forests of the Lower St-Lawrence region (Southern Quebec). Forests 8:120
Pedlar JH, McKenney DW, Aubin I, Beardmore T, Beaulieu J, Iversion L, O’Neill GA, Winder RS, Ste-Marie C (2012) Placing forestry in the assisted migration debate. Bioscience 62:835–842
Perring MP, De Frenne P, Baeten L, Maes SL, De Pauw L, Blondeel H, Caron MM, Verheyen K (2016) Global environmental change effects on ecosystems: the importance of land-use legacies. Glob Change Biol 22:1361–1371
Price DT, Cooke BJ, Metsaranta JM, Kurz WA (2015) If forest dynamics in Canada’s west are driven mainly by competition, why did they change? Half-century evidence says: climate change. Proc Natl Acad Sci USA 112:E4340
R Core Team (2016) R: a language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria https://www.R-project.org/. Accessed 29 June 2018
Ravenscroft C, Scheller RM, Mladenoff DJ, White MA (2010) Forest restoration in a mixed-ownership landscape under climate change. Ecol Appl 20:327–346
Régnière J, St-Amant R, Duval P (2012) Predicting insect distributions under climate change from ecophysiological responses: spruce budworm as an example. Biol Inv 14:1571–1586
Reich PB, Sendall KM, Rice K, Rich RL, Stefanski A, Hobbie SE, Montgomery RA (2015) Geographic range predicts photosynthetic and growth response to warming in co-occurring tree species. Nature Clim Change 5:148–152
Robitaille A, Saucier JP (1998) Paysages régionaux du Québec méridional. Publications du Québec, Sainte-Foy, QC
Sabogal C, Besacier C, McGuire D (2015) Forest landscape restoration: concepts, approaches and challenges for implementation. Unasylva 245:3–10
Scheller RM, Mladenoff DJ (2004) A forest growth and biomass module for a landscape simulation model, LANDIS: design, validation, and application. Ecol Model 180:211–229
Scheller RM, Mladenoff DJ (2008) Simulated effects of climate change, fragmentation, and inter-specific competition on tree species migration in northern Wisconsin, USA. Clim Res 36:191–202
Scheller RM, Domingo JB, Sturtevant BR, Williams JS, Rudy A, Gustafson EJ, Mladenoff DJ (2007) Design, development, and application of LANDIS-II, a spatial landscape simulation model with flexible spatial and temporal resolution. Ecol Model 201:409–419
Stanturf JA, Palik BJ, Dumroese RK (2014) Contemporary forest restoration: a review enphasizing function. For Ecol Manag 331:292–323
Steenberg JWN, Duinker PN, Bush PG (2013) Modelling the effects of climate change and timber harvest on the forests of central Nova Scotia, Canada. Ann For Sci 70:61–73
Strahan RT, Sanchez Meador AJ, Huffman DW, Laughlin DC (2016) Shifts in community-level traits and functional diversity in a mixed conifer forest: legacy of land-use change. J Appl Ecol 53:1755–1765
Sturtevant BR, Gustafson EJ, Li W, He HS (2004) Modeling biological disturbances in LANDIS: a module description and demonstration using spruce budworm. Ecol Model 180:153–174
Taylor AR, Boulanger Y, Price DT, Cyr D, McGarrigle E, Rammer W, Kershaw JA (2017) Rapid 21st century climate change projected to shift composition and growth of Canada’s Acadian Forest Region. For Ecol Manag 405:284–294
Terrail R, Arseneault D, Fortin MJ, Dupuis S, Boucher Y (2014) An early forest inventory indicates a high accuracy of forest composition data in early land-survey records. J Veg Sci 25:691–702
Vaillancourt MA, Gauthier S, Kneeshaw D, Bergeron Y (2009) Implementation of ecosystem management in boreal forests: examples from eastern Canada. Sustainable Forest Management Network, Edmonton, AB
van Vuuren DP, Edmonds J, Kainuma M, Riahi K, Thomson A, Hibbard K, Hurtt GC, Kram T, Krey V, Lamarque JF, Masui T, Meinhausen M, Nakicenovic N, Smith SJ, Rose SK (2011) The representative concentration pathways: an overview. Clim Change 109:5–31
Van Wagner CE (1987) Development and structure of the Canadian Forest Fire Weather Index System. Forestry Technical Report 35. Canadian Forestry Service, Ottawa, Canada
Wang WJ, He HS, Thompson FR, Fraser JS, Dijak WD (2016) Changes in forest biomass and tree species distribution under climate change in the northeastern United States. Landscape Ecol 32:1399–1413
Wilson BT, Lister AJ, Riemann RI (2012) A nearest-neighbor imputation approach to mapping tree species over large areas using forest inventory plots and moderate resolution raster data. For Ecol Manag 271:182–198
Yousefpour R, Jacobsen JB, Meilby H, Thorsen BJ (2014) Knowledge update in adaptive management of forest resources under climate change: a Bayesian simulation approach. Ann For Sci 71:301–312
Yousefpour R, Temperli C, Jacobsen JB, Thorsen BJ, Meilby H, Lexer MJ, Lindner M, Bugmann H, Borges JG, Palma JHN, Ray D, Zimmermann NE, Delzon S, Kremer A, Kramer K, Reyer CPO, Lasch-Born P, Garcia-Gonzalo J, Hanewinkel M (2017) A framework for modeling adaptive forest management and decision making under climate change. Ecol Soc 22:40
Acknowledgements
We thank G. Fortin, R. Terrail, A. deRomer, M. Leroyer for constructing the presettlement forest composition database. This research was funded by the Forest Change project of the Canadian Forest Service, Natural Resources Canada.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Boulanger, Y., Arseneault, D., Boucher, Y. et al. Climate change will affect the ability of forest management to reduce gaps between current and presettlement forest composition in southeastern Canada. Landscape Ecol 34, 159–174 (2019). https://doi.org/10.1007/s10980-018-0761-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-018-0761-6