
Abstract
Invasive pests pose a threat to the key environmental and social benefits provided by urban forests, and diverse tree planting is a primary management strategy for reducing pest vulnerability. For example, past urban forest losses to Dutch elm disease (DED) prompted municipal foresters to emphasize diversification, but it is unclear whether residential properties developed after the peak DED outbreak are actually more diverse than older properties. To address this issue, we inventoried all public and private trees on 150 residential properties in the Twin Cities Metropolitan Area, Minnesota, USA, and compared genus diversity on pre- and post-Dutch elm properties. We then quantified vulnerability to two current invasive pest threats, emerald ash borer (EAB) (Agrilus planipennis) and Asian longhorned beetle (ALB) (Anoplophora glabripennis), to evaluate whether higher diversity corresponds with lower pest vulnerability. We assessed vulnerability based on two fundamental urban forest metrics–frequency and size of vulnerable trees. Surprisingly, properties developed after the peak DED outbreak were less diverse than older properties. At the same time, less diverse post-Dutch elm properties exhibited low ALB vulnerability and modest EAB vulnerability, while more diverse older sites were highly susceptible to ALB. The importance of pest host specificity in characterizing urban forest vulnerability was underscored by low EAB vulnerability and high ALB vulnerability on our oldest study sites. This research highlights an apparent disconnect between the theoretical notion that higher diversity should reduce invasive pest vulnerability, and our empirical data indicating that genus diversity does not necessarily correspond with pest vulnerability.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Urban forests are a key component of the urban ecological landscape because they provide key environmental benefits including stormwater interception (Dwyer et al. 1992), air quality improvement (Nowak et al. 2006), and urban heat island reduction (Hardin and Jensen 2007). In addition, urban forests are a valuable structural asset (Nowak et al. 2002), and they require substantial economic investment for tree planting and maintenance (McPherson 2000; McPherson et al. 2005). Furthermore, urban forests provide socioeconomic benefits including recreation opportunities, aesthetics, privacy, and increased property values (Tyrväinen 1997; Payton et al. 2008). Trees on residential lands are particularly important because residential areas may account for half of urban land area and approximately 75 % of urban trees and related basal area (McPherson 1998). Thus, in light of the environmental, economic, and social values of urban forests, it is critically important to protect their long-term structural integrity.
Invasive pests and pathogens pose serious threats to urban forests. For example, Dutch elm disease (DED) devastated American elm (Ulmus americana) populations for decades following its introduction to North America in 1930 (Carter 1975). Originally from Asia and thus considered an exotic species in North America, the DED-causing fungi Ophiostoma ulmi and O. novo-ulmi played a key role in prompting urban foresters to consider the importance of diverse plantings to protect urban forests from future pest outbreaks (Miller 1997). In fact, various diversification goals have been proposed to improve long-term urban forest stability in the face of invasive pests (Barker 1975; Grey and Deneke 1986; Santamour 1990; Miller and Miller 1991). For instance, Santamour (1990) suggested that urban forests should contain no more than 10 % of a single species, no more than 20 % of a single genus, and no more than 30 % of a single family.
Such efforts to diversify urban forests met three key challenges. First, only a small set of species is well suited to stressful urban environments, so increased diversity may come at the expense of tree health and longevity where poorly suited species are planted (Richards 1982/1983). Second, nurseries carry limited ranges of species that do not satisfy the requests of urban foresters (Sydnor et al. 2010). Third, diversity goals have historically focused on municipal street tree and park tree populations (Barker 1975; Santamour 1990; Miller and Miller 1991). Consequently, this focus overlooks tree diversity on private residential properties where most urban trees are located (McPherson 1998). So while coordinated diversification efforts at the municipal scale may have increased public street tree diversity, it is largely unknown whether overall urban forest diversity increased following the DED outbreak and subsequent loss of elm trees, because most tree planting decisions are made by private landowners and land developers. For instance, reduced diversity in trees planted on residential properties, arising from uncoordinated planting decisions among many landowners at the household scale, may have offset or outweighed any diversity increases in street tree populations. Understanding whether overall urban forest biodiversity increased in residential areas following the devastating DED outbreak is crucial for evaluating the effectiveness of diversification efforts and vulnerability to future pest outbreaks.
At present, urban forests in the Midwestern US are threatened by two exotic beetles—the emerald ash borer (EAB) and Asian longhorned beetle (ALB). EAB (Agrilus planipennis) is a phloem-feeding beetle first discovered in North America near Detroit, MI in 2002 (Poland and McCullough 2006). As its name implies, EAB feeds on ash (Fraxinus) trees, which were once a popular planting choice in urban environments (Poland and McCullough 2006). In fact, an estimated 37.9 million ash trees grow on developed lands in the eastern US alone (Kovacs et al. 2010). EAB is spreading rapidly across North America (Prasad et al. 2010), and it generally kills infested ash trees within 5 years without ongoing insecticide treatments (McKenney and Pedlar 2012).
ALB (Anoplophora glabripennis) was discovered in the US before EAB (1996 and 2002, respectively; USDA APHIS 2013), but is currently not as widespread. To date, notable infestations have been reported in the metropolitan regions of New York, NY, Chicago, IL, Cincinnati, OH, and Worcester, MA (Haack et al. 2010; USDA APHIS 2013). Unlike EAB, ALB has been successfully eradicated in two states (USDA APHIS 2013), but ALB remains a major concern because it threatens approximately 30–35 % of urban trees in the US (Nowak et al. 2001; Smith and Wu 2008). According to Wang (2012), ALB’s broad range of preferred host genera includes maple (Acer), buckeye/horse chestnut (Aesculus), birch (Betula), willow (Salix), and elm (Ulmus). Additionally, Wang (2012) indicates occasional or rare ALB infestations have been noted in mimosa (Albizia), Katsura (Cercidiphyllum), ash, plane tree (Platanus), poplar (Populus), and mountain ash (Sorbus), with questionable accounts of US infestations in genera such as hackberry (Celtis), apple (Malus), and oak (Quercus). ALB kills trees via larval tunneling in the wood and cambium, which may impede vascular function and/or cause structural failure (Haack et al. 2010). Considered together, the combined threat posed by EAB and ALB place approximately half of the trees in eastern US cities at risk (Raupp et al. 2006), yet the linkages between tree diversity and invasive beetle vulnerability remain uncertain within inherently heterogeneous metropolitan regions.
In this study, we examined intra-regional variation in pest vulnerability across time and space within Minnesota’s Twin Cities Metropolitan Area (TCMA) by focusing on two central questions. First, is urban forest genus diversity higher on residential properties developed after the peak DED outbreak, as compared to older properties? Second, does higher urban forest diversity correspond with lower pest vulnerability, as measured by the frequency and sizes of vulnerable trees? We hypothesized that urban forest diversity would be higher on post-DED properties, reflecting a lesson learned in the importance of diversity following the DED devastation. We also hypothesized that this increased diversity on post-DED properties would correspond with lower EAB and ALB vulnerability on these sites. Given the limited data on US urban forest structure (Nowak et al. 2001), this type of vulnerability assessment advances the characterization of potential invasive beetle impacts on valuable residential urban forest resources.
Methods
Study area
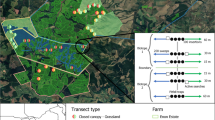
The study area is located within the TCMA, an area defined here as the 7,215 km2 seven-county region surrounding the central cities of Minneapolis and Saint Paul (Fig. 1). The TCMA had a 2010 population of 2.85 million (US Census Bureau 2010), and is expected to grow to 3.74 million by 2040 (Metropolitan Council 2012). From World War II (WWII) to present, the study area has experienced rapid urbanization, largely at the expense of row crop agricultural lands (Adams and VanDrasek 1993; Berland 2012). Minneapolis is characterized by older neighborhoods with compact property parcels situated along regularly gridded streets, while suburban neighborhoods have larger lots, often on winding streets with fewer boulevards bordering the streets (Fig. 2; Adams and VanDrasek 1993). In comparison to Minneapolis, suburban neighborhoods typically have a lower proportion of municipal street trees owing to the general lack of boulevards, less active tree planting programs, and proportionally smaller building footprints on a given parcel of land leaving more space for private trees. DED peaked in the TCMA in 1977; in that year, Minneapolis lost 32,000 trees and Saint Paul lost over 50,000 trees (Adams and VanDrasek 1993; French 1993). EAB was discovered in the TCMA in 2009, at the same time that we were collecting field data for this project. See http://gis.mda.state.mn.us/eab/ for a current map of EAB distribution in Minnesota. ALB has yet to be found in the TCMA.

Study area. a Minnesota is located in the north central United States. b Location of the study transect within the TCMA. c The study transect is situated along an urban–rural gradient from north to south. 150 residential study sites are categorized by housing age group

Characteristic urban form in each housing age group. All panels shown at 1:10,000 scale to highlight differences in property parcel sizes
Data collection
We collected data within a 120 km2 transect (40 km long by 3 km wide) positioned along an urban–rural gradient from the center of downtown Minneapolis (44.98°N 93.26°W) south through suburbs to the peri-urban fringe (Fig. 1). Confining our study to an urban–rural transect prevented us from making direct claims about urban forest diversity and vulnerability patterns within the entire TCMA, but this strategy had several advantages. For example, while similar ground-based studies of urban forest structure have typically focused on a single central city, we were able to characterize the urban forest across a range of housing ages and urban forms by sampling along an urban–rural transect (Fig. 2). This focus beyond the central city of Minneapolis was critical for obtaining adequate representation of suburban residential property parcels developed after the peak DED outbreak in 1977. In addition, this particular transect placement minimized complicating factors by avoiding the downtown Saint Paul secondary urban core and numerous water bodies found elsewhere in the TCMA. Finally, limiting our investigation to two counties reduced problems associated with inconsistent geospatial data and tax database availability across multiple counties.
We focused our study on residential land, which comprised 62 km2 (51.7 %) of the 120 km2 study transect. Candidate study sites were selected from a tax parcel database consisting of all residential properties (e.g., single family houses, duplexes, townhomes, apartment buildings) in the study transect. We originally drew 150 random candidate parcels and set study site quotas for each study area municipality (Fig. 1c) based on this initial drawing. When sampling permission was not obtained for a candidate site, it was replaced with a random parcel in the same municipality, thus ensuring that each municipality was adequately represented in the study. We did not pursue a sampling strategy aimed at inventorying equal numbers of property parcels or trees across housing age groups because parcels had varying numbers of trees. Targeting a set number of parcels in each housing age group (e.g., post-DED sites) would result in varying tree counts among housing age groups, while targeting a set number of trees would incorporate varying numbers of parcels and thus would include varying numbers of homeowners who make tree planting decisions. Hence, given that either target—equal trees or equal parcels—could affect the study results, we implemented a completely random sampling strategy to avoid potential biases introduced by an arbitrary sampling decision. Avoiding these housing age biases was particularly important because our primary goal was to compare diversity and vulnerability among housing age groups.
We visited candidate sites from May to October 2009, and asked for permission to sample the residential property. When permission was denied (n = 9) or nobody answered the door, we moved to the next candidate parcel on the list until we met each municipality’s quota. Qualitative observations indicating diverse participant demographics alleviated concerns about study participant bias toward those who were at home during the day (e.g., the retired). To ensure that our study sites were reasonably representative of their respective municipalities in terms of housing age, we compared mean study site housing ages for each municipality to the overall municipality mean derived from the tax parcel database. At each study site, we sampled all trees on the property, including all private and public land from mid-street to mid-alley. Consequently, the sample included both privately managed trees and municipal street trees, providing a more complete picture of urban forest composition on residential lands. Trees were defined as any woody vegetation >2.54 cm (1 in.) diameter at breast height (1.37 m; DBH). For each tree, we recorded its species and DBH to the nearest 0.1 cm. For multi-stem trees, DBH was summed for up to the six largest stems >2.54 cm DBH. While DBH is only one measure of tree size, it is closely related to other fundamental tree measures including height, crown diameter, and leaf area (Peper et al. 2001).
Data analysis
For data analysis, study sites were grouped into the following three age class bins based on the year of housing construction: pre-WWII (1889–1945), post-WWII (1946–1977), and post-DED peak outbreak (1978–2009). DED peak outbreak was characterized as the year when the most elms in the TCMA were lost to DED, which coincided with a spike in funding to fight the disease (Adams and VanDrasek 1993; French 1993). This sample stratification added a spatiotemporal component to facilitate comparison of older (primarily urban and near suburban) sites to post-DED sites concentrated in the outlying suburbs (Fig. 1c).
Genus diversity
We calculated diversity metrics to determine whether tree diversity was higher on residential properties developed after the peak DED outbreak. Several related considerations led us to calculate diversity at the genus level rather than the species level. Both EAB and ALB impact multiple species within genera (Poland and McCullough 2006; Haack et al. 2010). While vulnerability to both EAB (Rebek et al. 2008) and ALB (Dodds and Orwig 2011) varies by species within a particular genus, limited understanding prevents reliable quantitative estimates of species-level vulnerability. For example, Dodds and Orwig (2011) report contrasting findings between two studies assessing ALB preferences within the Acer genus. Finally, previous studies have set a precedent of assessing pest vulnerability at the genus level because that is the level at which pest management interventions would likely be made (Nowak et al. 2001; Raupp et al. 2006).
We measured diversity at the housing age group level as genus richness and as Simpson’s reciprocal index (1/D). Richness simply measures the total number of genera observed within each housing age group. Simpson’s index (D, Eq. 1) is a common and robust diversity index that accounts for both richness and dominance by the most abundant genera, with particular emphasis on dominance (Magurran 2004).
where n i is the number of individuals (or basal area) in the ith species, and N is the total number of individuals (or total basal area). Increasing values of Simpson’s reciprocal index (1/D) indicate increasing genus diversity within a housing age group. We calculated Simpson’s reciprocal index separately using both individuals and basal area, because diversity in terms of both individual trees and tree sizes offer useful descriptions of urban forest structure. While urban ecologists have typically calculated diversity indices using individuals (i.e., frequencies), calculating the index using basal area can potentially describe structural diversity better than simple tree counts, which do not account for differences in tree sizes among genera.
It is often difficult to assess whether biodiversity measures are significantly different among sampling groups (Solow 1993; Wiens et al. 1996), and our diversity comparisons were further complicated by an unequal sampling effort across housing age groups. In particular, we sampled uneven numbers of parcels across groups, parcels were different sizes, and parcels contained varying numbers of trees. To provide a reliable evaluation of housing age group genus diversity with respect to sampling effort and observed diversity across the entire study area, we implemented a nonparametric randomization technique developed by Solow (1993). The randomization test was first conducted for individual trees by pooling genus frequencies from all three housing age groups. Then we randomly partitioned the observed genera amongst the three age groups such that each age group contained the same number of trees as the observed sample data set, but not necessarily the same genus frequencies (after Wiens et al. 1996). We calculated genus richness and Simpson’s reciprocal index for each of 1,000 randomizations to serve as a null distribution, or the expected diversity in each housing age group, given the genera frequencies from the entire data set and the number of trees in each housing age group. Using a two-tailed test, the null hypothesis was rejected for any observed housing age group diversity values within the top or bottom 2.5 percentile of the null distribution (after Solow 1993).
Calculating a null distribution for Simpson’s reciprocal index using basal area was more complicated because, while each housing age group received the same number of trees as before, the total basal area for each housing group changed for each randomization. When performing this randomization, we maintained genus and basal area relationships for individual trees through the randomization process, and recomputed both n i and N for each randomization based on the basal area of individuals.
Pest vulnerability
We assessed variability in pest vulnerability among housing age groups based on two fundamental urban forest metrics–frequency and sizes of vulnerable trees. For EAB, all ash species were considered vulnerable. For ALB, we confined our analysis to what Wang (2012) lists as preferred genera, which include maple, buckeye, horse chestnut, birch, willow, and elm. This conservative approach of only considering preferred ALB hosts is appropriate given that to date ALB has spread considerably slower than EAB (USDA APHIS 2013). As described above, limited understanding of relative EAB and ALB species preferences within a particular genus prevented us from characterizing susceptibility at the species level. In addition, genus level vulnerability assessment may have more practical relevance given that pest management interventions are made at the genus level (Nowak et al. 2001; Raupp et al. 2006). First, we tested whether a particular housing age group’s observed vulnerable tree counts were different than would be expected based on random partitioning of all sampled trees among the three housing age groups. This assessment relied on the same randomization approach used to analyze genus diversity by individuals, and was conducted separately for EAB and ALB. Next, we used this randomization technique to assess whether the observed proportion of vulnerable basal area in each housing age group was significantly different than what would be expected given the overall proportion of vulnerable basal area in the entire data set. As with diversity calculations for basal area, we maintained genus and basal area relationships for individual trees throughout the randomization process. After comparing vulnerability across housing age groups, we calculated the number of study sites with at least one vulnerable tree for EAB or ALB, because having to remove or chemically treat just one vulnerable tree incurs a substantial cost for the household.
Results
We inventoried a total of 1,723 trees on 150 property parcels, with a mean and median of 11.5 and 6 trees per parcel, respectively. Overall, 41 genera were represented, with maple, spruce (Picea), ash, oak, and elm being the most common (Table 1). These five most common genera represented 55.3 % of all trees and 73.8 % of total basal area. Housing age groups generally followed an urban-to-rural gradient, with older homes located near the urban core and newer homes concentrated toward the peri-urban fringe (Fig. 1). By municipality, our mean study site ages were all within 3 years of the overall municipality mean, indicating that our sample was reasonably representative of the residential areas within the study transect.
Genus diversity
According to all three measures of diversity, post-DED parcels were less diverse than would be expected based on the overall genus abundances throughout the study area (Table 2). Post-WWII parcels had higher than expected diversity for both calculations of Simpson’s reciprocal index, while diversity values on pre-WWII parcels were within the expected ranges (Table 2).
Pest vulnerability
Tree counts and basal area were used to assess EAB and ALB vulnerability across housing age groups. Of the overall sample of 1,723 trees, 8.4 % were vulnerable to EAB, and 33.1 % were preferred ALB hosts (Table 3). Out of 150 residential properties, 66 (44.0 %) had at least one ash tree, and 126 (84.0 %) had at least one preferred ALB host (Table 3). For EAB, the proportion of vulnerable individuals was low on pre-WWII sites and high on post-WWII sites, while the proportion of vulnerable basal area fell within expected levels for each housing age group (Table 4). ALB vulnerability was high on pre- and post-WWII sites, but low on post-DED sites (Table 4).
Discussion
This study offers new perspectives on urban forest diversity and pest vulnerability across a metropolitan landscape. Given the high quantity of residential trees threatened, irrespective of position along the urban–rural transect, an invasion by either beetle has the potential to impose serious damage on existing urban forest structure. The current EAB outbreak in the TCMA is of great concern because EAB spreads rapidly (Prasad et al. 2010). While ALB has not demonstrated the ability to spread as rapidly as EAB, it threatens nearly four times as many trees as EAB, and ALB-prone trees were found on nearly twice as many properties as EAB hosts (Table 3). Furthermore, our ALB vulnerability estimate is conservative because we excluded occasional, rare, and questionable hosts from the analysis (see Wang 2012), and these hosts may ultimately prove to be important in ALB outbreaks. The high vulnerability rates, particularly with respect to ALB, reinforce the importance of identifying the distribution of susceptible genera within an urban forest (Nowak et al. 2001; Haack et al. 2010).
Lower genus diversity on post-DED sites
Our results indicate that lessons learned from previous DED devastation did not translate into tangible urban forest diversification in newly developed residential areas, which contrasts with our original hypothesis of higher genus diversity on post-DED sites. Consequently, several related explanations merit consideration. For example, newer homes may simply have fewer genera because homeowners have not yet finished planting trees. This is unlikely, however, because post-DED sites had more trees per site than the other housing age groups (Table 2). More likely, a narrow range of species was planted by developers across subdivisions or neighborhoods according to the trees that were fashionable at the time of development and valued for a combination of their aesthetic qualities, climatic hardiness, growth rates, or urban suitability (Miller 1997). As post-DED sites were at most 31 years old, most of the original trees were probably still alive on site. In contrast, some of the original plantings on older sites may have died at staggered intervals and been replaced with a broader range of species, leading to relatively higher diversity on older parcels.
Compared to older housing age groups, post-DED sites had fewer boulevards, less active municipal planting programs, and proportionally smaller house footprints which left more private land area for homeowner landscaping choices. Where homeowners made a higher proportion of tree planting choices, it is likely that these citizens were not aware of the rationale for planting diverse assemblages of trees, or that they prioritized factors like tree costs or landscaping aesthetics over biodiversity and other ecological considerations (Summit and McPherson 1998). A lack of coordinated municipal tree planting outreach programs in newer communities could help explain lower tree diversity in newer areas. In newly developed areas, diversity could potentially be enhanced via policies requiring land developers to meet specified planting diversity targets, or by implementing education or cost-share programs encouraging homeowners to consider diversity in tree planting decisions. For example, creating incentives for homeowners to plant trees from multiple families or orders could reduce vulnerability to pests that attack multiple species or genera (Raupp et al. 2006). This would both promote urban forest stability within the municipality as a whole and reduce the likelihood that individual homeowners would be burdened by future pest infestation costs (i.e., insecticide treatments or tree removal) for multiple trees at any given time.
Methodologically, we combined all trees within a housing age group to quantify biodiversity, so diverse planting at the household scale could be obscured if many households planted a diverse yet similar set of trees. This may occur, for example, where neighbors intentionally mimic one another in landscaping choices (Boone et al. 2010), where a common land developer uses a limited set of trees across entire subdivisions, or where neighborhood tree planting choices are influenced by a local retailer carrying a narrow selection of tree species. In fact, our results may align with Sydnor et al. (2010), who report widespread mismatches between what urban foresters request for diverse planting and the stock availability from nurseries in Ohio. Thus, while we were ultimately unable to determine the exact reason for lower diversity among post-DED sites, each of these explanations reflects the importance of spatial clustering of similar-aged houses, which has consistently proven useful for understanding variability in vegetation within urban landscapes (e.g., Hope et al. 2003; Grove et al. 2006; Berland 2012).
Diversity and pest vulnerability
Contrary to our original hypothesis, there was not a clear link between high diversity and low pest vulnerability. As evidence, even though post-DED sites had low genus diversity (Table 2), they also exhibited low ALB vulnerability and modest EAB vulnerability (Table 4). On the other hand, more diverse sites developed prior to the peak DED outbreak were highly vulnerable to ALB (Table 4). Based on these findings, this research challenges the traditional notion that biodiversity necessarily yields ecological stability. Richards (1982/1983) previously contested the importance of biodiversity for urban forest stability on the grounds that a diverse assemblage of trees poorly suited to urban environments is less stable than a smaller set of species well adapted to urban conditions. Our challenge is more direct, as our analysis did not suggest a positive relationship between diversity and resilience to EAB and ALB, but instead indicated the opposite connection for ALB. Diversification has been promoted to decrease pest vulnerability (e.g., Raupp et al. 2006), but in this study higher genus diversity did not correspond with lower vulnerability to the pests we studied. So while urban forest diversification strategies may prove useful for managing against devastating pest outbreaks in many situations, there is no guarantee that such measures will lead to tangible reductions in pest vulnerability because vulnerability is largely dependent on pest host preferences, which vary from pest to pest and may include species that are locally abundant or rare. Additional research is needed to determine whether our observed pattern is anomalous, or if it is emblematic of a larger disconnect between diversity and pest vulnerability.
Characterizing local- to regional-scale pest vulnerability is particularly challenging because different pests threaten different abundances and sizes of trees, and because we do not know which tree species will be affected by invasive pests arriving in the future. Yet, our finding that pre-WWII sites were highly vulnerable to ALB and low in vulnerability to EAB demonstrates the influence of pest host specificity on diversity–vulnerability linkages. The fact that post-WWII sites exhibited significantly high ALB vulnerability in terms of individual trees but expected levels of ALB vulnerability in terms of basal area highlights the importance of which particular trees on a landscape are susceptible to a pest. In this case, a significant number of individuals on post-WWII sites are susceptible to ALB, but those vulnerable trees do not represent a disproportionate amount of the basal area on these sites.
The differences in pest vulnerability according to housing age group highlight the importance of intra-regional spatiotemporal variability. In light of observed urban forest diversity differences and the influence of host specificity on pest vulnerability, certain neighborhoods and municipalities may bear disproportionate burdens associated with an invasive pest outbreak. At the municipal level, intra-regional variability in EAB and ALB vulnerability could conceivably lead to a highly unequal strain on city budgets, even for neighboring communities with similar socioeconomic characteristics. While post-DED sites did not exhibit high vulnerability to EAB or ALB, lower genus diversity on these sites may leave them particularly vulnerable to new invasive pest threats. At the household scale, documenting the number of residential properties with at least one vulnerable tree (Table 3) is important when considering the socioeconomic impacts of pest outbreaks, because pest invasion of a single tree crosses a threshold from zero to substantial costs for insecticide treatment or removal of the infested tree. For example, an EAB outbreak in this study area would only affect up to about one-fifth of pre-WWII households, but it could impact nearly three-fifths of post-WWII sites (Table 3). Variability among housing age groups was not as great for ALB, but a much higher percentage of homes stand to be affected by ALB as compared to EAB. Considering the documented pest-specific heterogeneity in urban forest vulnerability, similar analyses elsewhere could be useful to further elucidate the relationships among housing age, urban forest diversity, and pest vulnerability.
Conclusions
Invasive pests pose a serious economic threat to urban forests. They also threaten key urban ecosystem services, which may take decades to recover after a pest outbreak due to lagged growth of replacement trees. By characterizing urban forest structure and the potential losses associated with EAB and ALB outbreaks, this study provides the type of urban forest structural information that can help municipal foresters understand their risks and plan for future scenarios. Across the study area, EAB and ALB together threatened over 40 % of the trees sampled, and over 90 % of the study parcels had at least one tree susceptible to either beetle. By sampling across an urban–rural transect, we were able to compare pest vulnerability among housing age groups and incorporate vulnerability perspectives from understudied suburban areas. Our findings underscore the importance of pest host specificity when considering the potential impacts of invasive pests on urban forest resources. For example, pre-WWII sites exhibited low EAB vulnerability but high ALB vulnerability relative to the entire sample. Similarly, variability in urban forest structure, even among adjacent communities, may lead to uneven effects of invasive pest outbreaks within a metropolitan region.
We do not know which species will be impacted by the next invasive forest pest, so diversification is usually cited as the best strategy to promote stability in the face of pest outbreaks. Unexpectedly, genus diversity was lower on properties developed after the peak DED outbreak. Any lessons learned from DED seemingly did not translate into diversification of the study area’s residential urban forest, most likely because private land developers and homeowners are responsible for the majority of tree planting decisions on newer sites. Equally surprising, while newer properties were less diverse than older properties, they were also less vulnerable to ALB than older housing age groups. As such, there was an apparent disconnect between the theoretical notion that biodiversity should decrease vulnerability to invasive pests, and the data from this study indicating that biodiversity does not necessarily correspond with reduced vulnerability. Based on our findings, additional research is warranted to investigate the linkages between urban forest diversity and pest vulnerability. In particular, future work could assess whether our observed disconnect between diversity and vulnerability occurs in other places, examine the relative contributions of private trees versus municipal street trees in determining residential urban forest diversity, and investigate best management practices to simultaneously increase diversity and resilience in the residential urban forest.
References
Adams JS, VanDrasek BJ (1993) Minneapolis-St. Paul: people, place, and public life. University of Minnesota Press, Minneapolis
Barker PA (1975) Ordinance control of street trees. J Arboric 1:212–216
Berland A (2012) Long-term urbanization effects on tree canopy cover along an urban–rural gradient. Urban Ecosyst 15:721–738
Boone CG, Cadenasso ML, Grove JM, Schwarz K, Buckley GL (2010) Landscape, vegetation characteristics, and group identity in an urban and suburban watershed: why the 60 s matter. Urban Ecosyst 13:255–271
US Census Bureau (2010) Demographic profile data. http://factfinder2.census.gov. Accessed July 2013
Carter JC (1975) Major tree diseases of the century. J Arboric 1:141–147
Dodds KJ, Orwig DA (2011) An invasive urban forest pest invades natural environments—Asian longhorned beetle in northeastern US hardwood forests. Can J For Res 41:1729–1742
Dwyer JF, McPherson EG, Schroeder HW, Rowntree RA (1992) Assessing the benefits and costs of the urban forest. J Arboric 18:227–234
French DW (1993) History of Dutch elm disease in Minnesota. Minnesota Agricultural Experiment Station. http://purl.umn.edu/151957. Accessed July 2013
Grey GW, Deneke FJ (1986) Urban forestry. Krieger, Malabar
Grove JM, Troy AR, O‘Neil-Dunne JPM, Burch WR, Cadenasso ML, Pickett STA (2006) Characterization of households and its implications for the vegetation of urban ecosystems. Ecosystems 9:578–597
Haack RA, Hérard F, Sun J, Turgeon JJ (2010) Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: a worldwide perspective. Ann Rev Entomol 55:521–546
Hardin PJ, Jensen RR (2007) The effect of urban leaf area on summertime urban surface kinetic temperatures: a Terre Haute case study. Urban For Urban Green 6:63–72
Hope D, Gries C, Zhu W, Fagan WF, Redman CL, Grimm NB, Nelson AL, Martin C, Kinzig A (2003) Socioeconomics drive urban plant diversity. Proc Natl Acad Sci USA 100:8788–8792
Kovacs KF, Haight RG, McCullough DG, Mercader RJ, Siegert NW, Liebhold AM (2010) Cost of potential emerald ash borer damage in U.S. communities, 2009–2019. Ecol Econ 69:569–578
Magurran AE (2004) Measuring biological diversity. Blackwell, Malden
McKenney DW, Pedlar JH (2012) To treat or remove: an economic model to assist in deciding the fate of ash trees threatened by emerald ash borer. Arboric Urban For 38:121–129
McPherson EG (1998) Structure and sustainability of Sacramento’s urban forest. J Arboric 24:174–190
McPherson EG (2000) Expenditures associated with conflicts between street tree root growth and hardscape in California, United States. J Arboric 26:289–297
McPherson EG, Simpson JR, Peper PJ, Maco SE, Xiao Q (2005) Municipal forest benefits and costs in five US cities. J For 103:411–416
Metropolitan Council (2012) What lies ahead: population, household and employment forecasts to 2040. http://stats.metc.state.mn.us/stats/pdf/MetroStats_Forecasts.pdf. Accessed July 2013
Miller RW (1997) Urban forestry: planning and managing urban greenspaces. Waveland, Long Grove
Miller RH, Miller RW (1991) Planting survival of selected street tree taxa. J Arboric 17:185–191
Nowak DJ, Pasek JE, Sequiera RA, Crane DE, Mastro VC (2001) Potential effect of Anoplophora glabripennis (Coleoptera: Cerambycidae) on urban trees in the United States. J Econ Entomol 94:116–122
Nowak DJ, Crane DE, Dwyer JF (2002) Compensatory value of urban trees in the United States. J Arboric 28:194–199
Nowak DJ, Crane DE, Stevens JC (2006) Air pollution removal by urban trees and shrubs in the United States. Urban For Urban Green 4:115–123
Payton S, Lindsey G, Wilson J, Ottensmann JR, Man J (2008) Valuing the benefits of the urban forest: a spatial hedonic approach. J Environ Plan Manag 51:717–736
Peper PJ, McPherson EG, Mori SM (2001) Equations for predicting diameter, height, crown width, and leaf area of San Joaquin Valley street trees. J Arboric 27:306–317
Poland TM, McCullough DG (2006) Emerald ash borer: invasion of the urban forest and the threat to North America’s ash resource. J For 104:118–124
Prasad AM, Iverson LR, Peters MP, Bossenbroek JM, Matthews SN, Sydnor TD, Schwartz MW (2010) Modeling the invasive emerald ash borer risk of spread using a spatially explicit cellular model. Landscape Ecol 25:353–369
Raupp MJ, Cumming AB, Raupp EC (2006) Street tree diversity in eastern North America and its potential for tree loss to exotic borers. Arboric Urban For 32:297–304
Rebek EJ, Herms DA, Smitley DR (2008) Interspecific variation in resistance to emerald ash borer (Coleoptera: Buprestidae) among North American and Asian ash (Fraxinus spp.). Environ Entomol 37:242–246
Richards NA (1982/1983) Diversity and stability in a street tree population. Urban Ecol 7:159–171
Santamour FS (1990) Trees for urban planting: diversity, uniformity, and common sense. In: Proceedings of the 7th conference of the metropolitan tree improvement alliance, pp 57–65
Smith MT, Wu J (2008) Asian longhorned beetle: renewed threat to northeastern USA and implications worldwide. Int Pest Control 50(311):316
Solow AR (1993) A simple test for chance in community structure. J Anim Ecol 62:191–193
Summit J, McPherson EG (1998) Residential tree planting and care: a study of attitudes and behavior in Sacramento, California. J Arboric 24:89–97
Sydnor TD, Subburayalu S, Bumgardner M (2010) Contrasting Ohio nursery stock availability with community planting needs. Arboric Urban For 36:47–54
Tyrväinen L (1997) The amenity value of the urban forest: an application of the hedonic pricing method. Landsc Urban Plan 37:211–222
USDA APHIS (2013) Asian longhorned beetle. USDA Animal and Plant Health Inspection Service. http://asianlonghornedbeetle.com/. Accessed July 2013
Wang B (2012) Asian longhorned beetle: annotated host list. http://www.aphis.usda.gov/plant_health/plant_pest_info/asian_lhb/downloads/hostlist.pdf. Accessed July 2013
Wiens JA, Crist TO, Day RH, Murphy SM, Hayward GD (1996) Effects of the Exxon Valdez oil spill on marine bird communities in Prince William Sound, Alaska. Ecol Appl 6:828–841
Acknowledgments
This material is based upon work supported by the National Science Foundation under Grant #1003138, and by the Graduate School at the University of Minnesota. We thank Tom Crist for helpful discussions of biodiversity metrics, and Brewster Malevich for field assistance. Three anonymous reviewers and the handling editor provided constructive comments that improved the quality of the paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Berland, A., Elliott, G.P. Unexpected connections between residential urban forest diversity and vulnerability to two invasive beetles. Landscape Ecol 29, 141–152 (2014). https://doi.org/10.1007/s10980-013-9953-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-013-9953-2