
Abstract
Cattle move to access patches that vary in forage quantity and quality. Fragmentation can prevent animals from reaching patches. I used an integrative ecosystem model applied to three African landscapes to explore the sensitivity of cattle populations to fragmentation (here, changes in populations as parcel areas decreased) under different precipitation patterns. I hypothesized that low and high precipitation would yield populations relatively insensitive to fragmentation, intermediate precipitation would yield more sensitive populations, and more variable inter-annual precipitation would reduce sensitivity to fragmentation. Precipitation data were altered to yield averages of 100–1,000 mm year−1 and inter-annual coefficients of variation of 0–60%. A 1,000 km2 landscape in each area was divided into progressively smaller parcels and simulations conducted for each parcel. Rainfall at 100 mm year−1 supported low populations that were insensitive to fragmentation. Populations peaked at rainfall levels similar to those observed, and declined under higher precipitation, due in-part to shrub expansion. Fragmenting landscapes caused up to a 62% decline in cattle. High inter-annual variation in precipitation reduced sensitivity to fragmentation when precipitation was above that observed. The pattern was opposite when precipitation was below what was observed. Cattle on the landscape with fine-scale heterogeneity were relatively insensitive to fragmentation, and those on the heterogeneous but coarse-grained landscape were extremely sensitive. Fragmentation in landscapes where populations are sensitive will require more intensive inputs to offset losses, and changes in the frequency of extreme weather associated with climate change will alter the sensitivity of some populations to fragmentation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Cattle either move or are moved by herders to access patches that maximize forage intake, constrained by such things as dietary selection and digestibility, risk of predation or disease, thermal comfort, and access to other resources such as water. Patches may be relatively stable in the forage they provide throughout the year (e.g., swamps), or vary in a predictable, synchronous way (e.g., high elevation meadows). More commonly, ephemeral patches vary asynchronously in the quantity or quality of forage they provide, related to the amount of precipitation they receive (Ash et al. 2004). Arid and semi-arid lands have precipitation that is highly variable in space and time (Conrad 1941; Nicholson 1980; Ellis and Galvin 1994), which can lead to very dynamic and widely-spaced forage patches. In intact semi-arid landscapes, cattle may move hundreds of kilometers annually to access forage and other resources (e.g., Bekure et al. 1991; reviewed in Boone et al. in press).
Fragmenting landscapes may prevent animals from reaching the forage patches they seek. Fragmentation has been well-studied in landscape ecology (e.g., Saunders et al. 1991; Andren 1994; Fahrig 2003; Ewers and Didham 2005; Cushman 2006), and been defined in many ways (reviewed in Fahrig 2003). Fragmentation includes habitat loss or changes in land cover through such things as forest harvest and urbanization, as well as restrictions in access. Here, I focus on effects of the later, fragmentation of landscapes into more numerous but smaller parcels without loss of habitat, land cover change, or increased isolation between patches. Semi-arid and arid landscapes are fragmented into small parcels through fencing (Boone and Hobbs 2004). Although fencing of rangelands is not as visually dramatic as clear cutting of forests, for instance, effects of fragmentation in rangelands can be profound. Theory proposes and research has shown that secondary production is often inversely related to fragmentation of the landscapes inhabited, as the ability of animals to access diverse forage patches is reduced (Ash and Stafford Smith 1996; Ritchie and Olff 1999; Doncaster 2001; Ash et al. 2004). As the mobility of cattle declines through fencing, sedentarization of pastoralists, or other means, animals are no longer able to access the forage patches they wish, and instead must consume forage that is poorer quality, poorer quantity, or more risky to acquire. Boone and Hobbs (2004) simulated a 19% reduction in the number of animals that could be supported on a 300 km2 landscape in South Africa when fragmented to 10 km2 fragments, hereafter called parcels, versus when it was intact. Here I term such a population change as sensitivity to fragmentation. Boone et al. (2005) found ranches subdivided into 1 km2 parcels in Kenya supported up to 25% fewer animals compared to when the ranches were intact, which had profound impacts for modeled Maasai households (Thornton et al. 2006). In both modeling efforts, we used the observed weather patterns in simulations, or patterns derived using random draws from the observed data. Rangelands vary greatly in the spatial distribution and inter-annual variability of rainfall (Ellis and Galvin 1994). How populations in different landscapes vary in their sensitivity to fragmentation under different precipitation patterns has not been explored.
In the current study, I used a process-based ecosystem model applied to three African landscapes to explore the effects of fragmentation and different precipitation patterns on cattle populations. The landscapes span a range of annual precipitation amounts, which in these semi-arid areas is closely related to productivity, and include a site of low fine- and coarse-grain heterogeneity (North-West Province, precipitation averaging 400 mm year−1), one that is heterogeneous at a fine-scale (Kajiado, 550 mm year−1), and one heterogeneous at a coarse-scale (Ngorongoro, 800 mm year−1). For each area, a landscape was fragmented into smaller and smaller parcels, and for each parcel, simulations were conducted with precipitation spanning from xeric to almost mesic, and with precipitation between years spanning from entirely predictable to extremely variable. I expected to see that increased variability in rainfall and primary productivity would cause cattle populations to decline (e.g., Lewontin and Cohen 1969; Davis et al. 2002). Also, a quadratic or hump-shaped relationship between primary productivity and the sensitivity of cattle to fragmentation has been hypothesized (Boone et al. 2005; Boone et al., in press). Areas with low productivity (in this study, equivalent to low annual precipitation) may have animals so sparsely populated that travel costs to reach patches are maximized, and fragmentation into parcels larger than the areas over which animals range would not cause declines. In highly productive areas (i.e., high precipitation), small parcels can provide adequate forage, making movements less important and minimizing the effects of fragmentation. Areas of intermediate productivity are hypothesized to have populations most sensitive to fragmentation. I hypothesized that low inter-annual variability in rainfall would promote higher cattle populations that would increase sensitivity to fragmentation, and that high inter-annual variability in rainfall would lead to primary and secondary productivity being weakly linked (Ellis and Swift 1988) and populations less sensitive to fragmentation. Lastly, populations in landscapes that are homogeneous are hypothesized to be less sensitive to fragmentation than those in landscapes intermediate in heterogeneity (Boone et al. 2005). In homogeneous landscapes where parcels all provide similar resources, fragmentation is less likely to lead to declines in cattle. In turn, landscapes that are very heterogeneous at a fine scale will have populations that are less sensitive to fragmentation than those that are intermediate in heterogeneity. The fine-grained (Forman 1995) nature of resources within such landscapes means animals will be able to fulfill their needs in small parcels.
Study areas
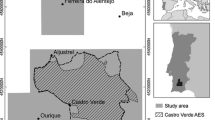
Ecosystem model applications to three African sites were used in simulations. The sites (Fig. 1) were:

Three study areas were used in simulations, (a) Vryburg 1 District of the North-West Province, South Africa, (b) southern Kajiado District, Kenya, with group ranch boundaries included, and (c) Ngorongoro Conservation Area, Tanzania. For each area, a 1,000 km2 area was modeled, with its outline shown in black. Topography is shaded, and areas are shown in their countries as inserts. For scale, each area modeled was 32 km across
North-West Province, South Africa (Boone et al. 2004; Thornton et al. 2004)—a gradient in precipitation spanned from an average of 500 mm year−1 on the eastern side of the area to 300 mm year−1 on the northwestern edge bordering the Orange River and the Kalahari Desert. Precipitation in the area averaged 400 mm year−1, was very patchy, and inter-annual variability was high, with a coefficient of variation (CV) from 1960 to 1995 of 37.4%. The area was dominated by thornveld and bushveld. The application included five domestic livestock species and seven plant functional groups: high, moderate, and low palatability perennial grasses, annual grasses, acacia shrubs, camphorbush shrubs (Tarchonanthus camporatus), and acacia trees (Acacia sp.).
Southern Kajiado District, Kenya (Boone 2005; Boone et al. 2005; Thornton et al. 2006)—precipitation ranged from 400 mm to 800 mm year−1 in the study area, with more rainfall to the northwest and on mountain slopes. From 1969 to 1998, rainfall averaged 550 mm year−1, with an inter-annual CV in precipitation of 27.6%. Grasslands, wooded grasslands, and brushlands dominate the district. The model application included nine wild and domestic animal functional groups and seven plant functional groups: palatable grasses, palatable forbs, unpalatable grasses and forbs, swamps represented by papyrus (Cyprus papyrus), palatable and unpalatable shrubs, and deciduous woodlands, with acacias forming the dominant overstory species.
Ngorongoro Conservation Area, Tanzania (Boone et al. 2002; Thornton et al. 2003; Galvin et al. 2004, 2006; Boone et al. 2006)—climate was diverse in Ngorongoro, with rainfall more than 1,000 mm year−1 within the high-altitude forest reserve, and less than 450 mm year−1 in the southern Serengeti Plains, in the shadow of the Ngorongoro mountains. From 1963 to 1992, Ngorongoro Conservation Area received an average rainfall of 800 mm year−1, with an inter-annual CV of 20%. In the ecosystem application, I included 7 plant and 11 animal functional groups. The plants modeled in the application were: palatable grasses, palatable forbs, unpalatable grasses and forbs, palatable and unpalatable shrubs, evergreen trees of the highland forests, and deciduous woodlands, which were mostly acacias.
Methods
Ecosystem model
I use the process-based, spatially explicit ecosystem model Savanna, which is well suited to represent variable cattle herbivory on landscape fragments. Development of the model began more than 20 years ago by M. Coughenour of Colorado State University, applied to the Turkana District of Kenya (Coughenour 1985), with the model updated and used widely since (e.g., Coughenour 1992; Eastman et al. 2001; Ludwig et al. 2001; Boone et al. 2002, 2005; Thornton et al. 2004). The model divides landscapes into a grid of square cells, and for each cell, ecological processes are modeled. Savanna reads spatial data that characterizes the cells to elevation, slope, aspect, soils, and vegetation type. Precipitation from a series of weather stations is read by the program, plus more detailed data for a station with ample data that includes minimum and maximum temperature, wind speed, radiation, humidity, and CO2 concentration. During each weekly time step of simulations, plant functional groups compete for soil moisture, light, space, and nutrients. In each time step, the plants may produce seed, germinate, out-compete other functional groups and gain ground cover, be browsed, or may die. Cattle feed on the forage produced and gain energy from it. The cattle expend energy through basal metabolism, travel, and lactation. Net energy gained goes to reproduction and weight gain, which is reflected in condition indices. Condition indices are linked to cattle birth rates and death rates, so that there is potential for density dependent feedback in cattle populations (see Ellis and Coughenour 1998; Boone 2000 for more detail).
Model modifications
The sites cover a range of African landscapes, average annual rainfall from 400 to 800 mm, and CVs of annual rainfall from 20 to 37.4%. I wanted to retain the different representations of African grazing systems in these analyses, but standardize the applications to make interpretation of the results easier. For each study area, a 1,000 km2 area was modeled at 1 km2 resolution (i.e., a 32 × 32 km2 landscape, with 8 cells in the southern-most row modeled). The locations of the 1,000 km2 parcels are shown in Fig. 1, and were: North-West Province—24.140 decimal degrees latitude, −26.357 longitude to 24.464, −26.071; Kajiado—37.309, −2.470 to 37.597, −2.759; Ngorongoro—35.339, −2.785 to 35.627, −3.074.
In each case, the plant functional groups in the original applications were retained, but I reduced the number of animal groups modeled to a single group, cattle, following Boone and Hobbs (2004) and Boone et al. (2005). Interpreting simulation results when up to 11 animal functional groups change in compensatory ways can be confusing. Representing a single cattle population makes inferences about the capacity of the grazing lands to support cattle more straightforward, and the relationship between fragmentation and the mobility of cattle has important implications for pastoral livelihoods (Thornton et al. 2006). The initial densities of cattle in simulations were set as in the original applications (i.e., North-West Province: 14 animals/km2; Kajiado: 19 animals/km2; Ngorongoro 35 animals/km2). Habitat selection by animals in Savanna is usually influenced by the distance to water. In these analyses animals used parcels exclusively, and so as land was fragmented few parcels would contain water, and animals elsewhere would not survive without their moving or water being transported. The distribution of water sources was not a focus of this research, so I disabled that relationship. Prior to the start of analyses, in simulations on the intact 1,000 km2 parcels, I made small changes (≤0.02 MJ/kg/day) to the energy requirements of the cattle until their populations remained relatively constant over 40 year simulations, although the populations changed in response to wet and dry years.
Weather data
For each of the applications, weather data was available for 30 or more years (North-West Province: 1960–1995, 166 stations; Kajiado: 1969–1998, 47 stations; Ngorongoro: 1963–1992, 55 stations). I altered the rainfall amount and inter-annual CV in rainfall for each study area, using the observed rainfall pattern as a template. Altering the CV of rainfall required care to avoid changing its seasonality or spatial pattern. For each data set, I used a coefficient applied to the deviation from the mean precipitation and an offset constant applied to the mean to essentially reduce or elevate rainfall (mean) and stretch or compress variability (CV). The approach yielded appropriate rainfall amounts and variability in rainfall, without altering seasonality or the spatial pattern of rainfall. I used a custom program to alter average rainfall across years so that it spanned from 100 to 1,000 mm (i.e., 100, 200, 300 ... 900, 1,000 mm) and the inter-annual CV to span from 0 to 60% (0%, 5, 10 ... 55, 60%) (Fig. 2). It was not possible using coefficients applied to observed rainfall to simulate CVs equal to 0%. In practice, the minimum CV I could calculate was <0.3%.

The precipitation record from 1960 to 1995 from the North-West Province, South Africa, adjusted to yield annual rainfall from (a) 200 mm, (b) 600 mm, and (c) 1,000 mm, and inter-annual coefficients of variation (CV) from 0 to 60%. In practice, rainfall for each area was incremented by 100 mm from 100 to 1,000 mm, and CV by 5% from 0 to 60%. [Reprinted from Boone and Wang (2007) with permission from Elsevier]
Fragmentation
I sought to quantify the effects of fragmentation of landscapes on cattle capacity under variable precipitation patterns. The 1,000 km2 parcel, modeled with cells 1 km × 1 km, was divided into progressively smaller parcels, until the area was ultimately represented as 100 parcels each 10 km2 (i.e., 1 parcel at 1,000 km2, 2 parcels at 500 km2, 3 parcels at 333 km2 ... 50 parcels at 20 km2, 75 parcels at 13 km2, and 100 parcels at 10 km2). In every simulation the underlying model had a resolution of 1 km × 1 km, but those cells were grouped into different parcel sizes to which cattle had access. For instance, at the smallest parcel size of 10 km2, animals within each parcel had ten cells from which to choose. Figure 3 shows the spatial arrangement of the parcels modeled. Figure 4 summarizes the standard deviation in mean elevation across parcels as each study site was fragmented. The change in average standard deviation in the elevation of parcels reflects the inter-parcel variability within each site. The heterogeneity of the Ngorongoro site relative to the North-West Province and southern Kajiado is apparent. Animals in Ngorongoro may move shorter distances to access vegetation that is presumably of a different type, or at a different phenological stage. Animals may occur at higher densities in more heterogeneous than in more homogenous sites. That was true for cattle in these three sites, but differences in precipitation and productivity confound a simple comparison. For each area (n = 3), a simulation was conducted with animals confined to each parcel (1 + 2 + 3 + … + 50 + 75 + 100 = 394 parcels), for each rainfall amount (100 to 1,000 mm; 10 levels) and CV (0 to 60%; 13 levels) combination, yielding 153,660 simulations (3*394*10*13). I created a local area network with a host computer storing simulation results and several client computers running simulations to conduct analyses in a timely way.

The 1,000 km2 areas were divided into progressively smaller parcels, and simulations were conducted for every combination of precipitation amount and variation for each parcel. Parcels spanned from a single 1,000 km2 area to 100 parcels each 10 km2 in area

Variability in mean elevation between parcels as study sites were fragmented. North-West Province—solid circles, southern Kajiado District—open triangles, Ngorongoro—open circles
Analysis
My focus is not that small parcels can support fewer animals than large parcels, but that landscapes divided into small parcels can support fewer animals; I report the number of animals on the landscape, summing populations across parcels. In Savanna simulation results, as in reality, animal populations exhibit annual cyclic changes as births occur in single season and deaths occur throughout the year. For each set of results, I calculated the average cattle population at the annual peak population size simulated (March in Ngorongoro and Kajiado, November in the North-West Province). I also calculated metrics reflecting ecosystem status, including leaf, root, standing dead, and total biomass for herbaceous plants, shrubs, and trees, and cover for shrubs and trees. I plotted the log10 of parcel area against the sum of the number of animals that could be supported across the entire 1,000 km2 area. I used linear regression with Systat 7 (SPSS, Inc., now Systat Software, Inc., Richmond, CA, USA) to identify least-squares smoothed lines. Slopes that differed from 0 indicated landscape-climate combinations that were sensitive to fragmentation, in so far as cattle populations changed when the landscape was divided into small parcels. Multiple linear regression was used as an exploratory rather than predictive tool, to summarize the amount of variation in sensitivity to fragmentation that could be explained by precipitation patterns in each site.
Results
Results for the North-West Province (Fig. 5) include several responses typical of all three sites. First, I will focus on the effects of climate on animal populations independent of effects of fragmentation (i.e., the intercepts in graphs in Fig. 5, which plot populations on a single intact landscape of 1,000 km2). Rainfall at 100 mm year−1 allowed few animals to be supported. More animals were supported with higher average rainfall. In the North-West Province, the population reached a maximum at 300–400 mm year−1 rainfall (Supplement A shows populations in the intact landscapes under the different precipitation patterns). At greater rainfall amounts, low inter-annual CVs in precipitation allowed shrub cover to expand to 42% (Supplement B), which reduced the number of grazing cattle that could be supported over the long-term. Also, in some cases increased precipitation led to populations that grew to exceed the capacity of the parcel, and then declined. In contrast, at high inter-annual CVs, frequent drought did not allow cattle populations to increase to the same degree, and the more modest increase in shrub cover (to ca. 30%) did not cause a decline in populations. At peak cattle populations (i.e., where precipitation was 300–400 mm year−1), increased variability in precipitation led to declines in cattle populations.

Cattle populations in the North-West Province, South Africa, under decreasing parcel sizes for all combinations of rainfall amount and variability
The slope of each line (Fig. 5) indicates the sensitivity of the cattle population to fragmentation under the given precipitation pattern. North-West Province cattle were relatively insensitive to fragmentation when rainfall was 400 mm year−1 or below and inter-annual CV in rainfall was 25% or less (Fig. 5). The cattle were most sensitive to fragmentation when rainfall was 500 mm year−1 and the CV was low (≤15%). The predictable climatic patterns and low shrub cover allowed the cattle populations to build to high levels in the intact landscape. Then, as the landscape was fragmented into smaller isolated parcels, animals were unable to access remote green forage, condition indices declined, and populations were reduced (Fig. 5). At higher annual rainfall, the lines are not as steeply sloped because of lower populations on the intact landscape due to shrub expansion. In contrast, at higher rainfall amounts and higher CVs, less shrub cover led to fragmented parcels with more homogenous herbaceous forage patches and a cattle population less sensitive to fragmentation. These relationships taken together suggest an interaction, where intermediate conditions would yield greater sensitivity to fragmentation (steeper slope in the figures); that region appears to be at 600–700 mm and a CV of 30% in the North-West Province (Fig. 5). That said, most precipitation amounts ≥200 mm and CV ≥15% yielded cattle populations sensitive to fragmentation (slopes ≠ 0, P ≤ 0.05). In the North-West Province, 67% of the variation in slopes could be explained by annual rainfall, inter-annual CV, and their interaction (P < 0.001).
Results for southern Kajiado District, Kenya show few animals supported at 100 mm rainfall, the population peaking at 500–600 mm annual rainfall, and peaking at higher rainfall levels when inter-annual CVs were ≥50% (Fig. 6; Supplement A). However, sensitivity to fragmentation was mixed. Shrubs increased to 86% cover under the highest precipitation amounts. Herbaceous biomass (Supplements C, D) declined sharply with increasing annual rainfall and declining CV, and ultimately, increasing shrubs. In general, cattle populations were sensitive or neutral to fragmentation with precipitation ≤500 mm. Small declines in simulated cattle populations occurred as the land was fragmented. At higher annual rainfall amounts, populations were neutral or increased in response to fragmentation (Fig. 6). We have simulated such increases in southern Kajiado District in past research (Boone et al. 2005). Animals from throughout an intact 1,000 km2 landscape move to the most productive areas, increasing competition with resident animals. However, if the area is fenced into parcels, cattle cannot immigrate into the optimum parcels, and the populations within those parcels can increase markedly. If the parcels that remain can support populations similar to those when the landscape was intact, the overall population can increase under fragmentation. The shallow slopes and mixed patterns led to 14% of the variation in sensitivity to fragmentation being explained by annual rainfall, inter-annual CV, and their interaction (P < 0.001).

Cattle populations in southern Kajiado District, Kenya, under decreasing parcel sizes for all combinations of rainfall amount and variability
Ngorongoro is the most topographically diverse of the three sites modeled (Fig. 4), and even at 100 mm year−1 rainfall minor sensitivity to fragmentation is evident (Fig. 7). Populations were lowest at 100 mm annual rainfall, and increased to a peak at about 800 to 900 mm rainfall when CV was <45%. I believe a peak in population would occur at precipitation greater than 1,000 mm and CVs >45%, but that is beyond the range of annual precipitation modeled. Cattle populations were more sensitive to fragmentation at high inter-annual CVs in rainfall than at low CVs, sometimes dramatically. For example, at 60% CV and rainfall of 1,000 mm year−1, the total number of cattle supported on the intact landscape was 51,000, but that declined to 19,160 when the land was divided into one hundred 10 km2 parcels (Fig. 7). At low CVs, consistent rainfall across years allowed the majority of parcels to provide adequate forage year-round, but frequent droughts under high CVs caused the animals confined to the least productive parcels to decline. Overall, 53% of the variation in slopes, representing sensitivity to fragmentation, could be explained by annual rainfall, CV, and their interaction (P < 0.001). In follow-up exploratory analyses, Ngorongoro was the only site where shrub cover increased explanatory power, and then only slightly (to 55% variation explained). Shrub cover is closely related to annual precipitation and the inter-annual CV of precipitation (i.e., ≥90% of variation in shrub cover explained), so shrub cover acts as a surrogate for the precipitation metrics.

Cattle populations in Ngorongoro Conservation Area, Tanzania, under decreasing parcel sizes for all combinations of rainfall amount and variability
Discussion
I have used process-based, integrative ecosystem modeling to quantify potential effects of landscape fragmentation on cattle populations under variable precipitation in three African landscapes that span gradients in their observed annual precipitation and in topographic heterogeneity. The hypothesis that productivity, which is closely related to precipitation in these semi-arid areas, would affect sensitivity to fragmentation in a hump-shaped relationship was confirmed. In the North-West Province (Fig. 5), for example, the slope of lines showing changes in the cattle population were steepest at 500–600 mm precipitation; sensitivity to fragmentation was lower when precipitation was below or above that range. I had hypothesized that low inter-annual variability in precipitation would make populations more sensitive to fragmentation as cattle increased under predictable precipitation. That was generally true where rainfall was greater than that observed in the sites (e.g., North-West Province, Fig. 5, 500 mm column; Kajiado, Fig. 6, 700 mm column; beyond the precipitation range tested in Ngorongoro, Fig. 7). For analyses with precipitation below the observed average the pattern was opposite, with landscapes becoming more sensitive to fragmentation as inter-annual CV of rainfall increased. The North-West Province provides the clearest example. With rainfall at 500 mm year−1, which is above the mean of the area, populations in the fully fragmented landscape were the same or smaller as inter-annual variability in CV increased. However, there was a decline in the number of animals that could be supported in the intact landscapes as the CV increased (Fig. 5, Supplement A), resulting in decreasing slopes. Stable precipitation across years allowed populations in the intact landscapes to build, whereas when CVs were high and precipitation unstable across years, droughts caused the populations to decline (Ellis and Swift 1988; Boone and Wang 2007). When rainfall was below that observed for the site (e.g., Fig. 5, 300 mm column), populations also declined as inter-annual CV in rainfall increased, but they declined farther in fragmented landscapes, yielding sloping lines as CV increased. Droughts had a greater effect in the fragmented landscapes under conditions of lower precipitation. This general result is relevant to conditions under climate change where precipitation declines and becomes more variable, as anticipated for many African regions and confirmed in South Africa (Mason et al. 1999; Easterling et al. 2000).
We have hypothesized that landscapes of very low heterogeneity and very high heterogeneity and fine grain are less sensitive to fragmentation than landscapes that are intermediate in heterogeneity (Boone et al. 2005). Here I used three landscapes, so inferences related to landscape heterogeneity are limited (Fahrig 2003). Kajiado has fine-grained heterogeneity, and was relatively insensitive to fragmentation, but increases in shrub cover complicated interpretation. Ngorongoro was extremely sensitive to fragmentation, but the grain of heterogeneity within that landscape is coarse. The landscape modeled in Ngorongoro includes portions of the Serengeti Plains, the Gol Mountains, Olbulbul Depression, and the slopes of the craters of Ngorongoro. As the landscape was fragmented, parcels were distinct in nature, whereas in a heterogeneous but fine-grained landscape like Kajiado, parcels were similar (Fig. 4).
At low inter-annual CVs in rainfall, populations peaked at about the observed annual rainfall in each of the three landscapes modeled (Figs. 5, 6, 7; Supplement A). An increase in secondary productivity is expected as primary productivity increases (e.g., Phillipson 1975; Coe et al. 1976; East 1984). With primary productivity higher still, cattle populations can build to a high density, then decline in a density dependent response. Also, shrub encroachment at higher precipitation amounts would lead to a decline in grazers like the cattle in these simulations. A peak in animal populations is therefore expected. That the peak in each case is at the observed annual rainfall amount is likely an artifact of adapting the simulation rather than an emergent result. The applications were set-up to yield relatively stable cattle populations, shrub cover, tree cover, etc. over the long-term using the observed rainfall pattern.
Results from Kajiado District, Kenya, mirror in some ways those reported in Boone et al. (2005). In that work, we used observed but randomized weather patterns to quantify changes in livestock numbers from fragmentation. In group ranches that were productive, landscapes were not sensitive to fragmentation, whereas those that were less productive were more sensitive. In this work, relatively small changes in productivity led to different sensitivities to fragmentation. Changes in cattle numbers are less pronounced in these analyses. I have shown this is partially due to shrub encroachment, but it is probably also related to the use of a single species, whereas in Boone et al. (2005) the full complement of 11 species or groups of species were modeled, and to the minimum parcel area analyzed, which was 1 km2 in Boone et al. (2005) and 10 km2 here.
The population summations shown mask a great deal of variability in responses for individual parcels. Landscapes may contain key resources, such as wetland depressions, and more expansive lower productivity grazing areas. As the landscape is fragmented, parcels that contain key resource areas can support many animals. Populations in these parcels may build to high levels, protected from immigrants that they would otherwise have to compete with during forage bottleneck periods (Fahrig 2003; Boone et al. 2005). Populations in favorable parcels may build to an unsustainable level and decline. Other areas may only support a fraction of what they did when the land was intact. Parcels with forage availability changing synchronously may support few animals, because of bottlenecks during the low ebb of forage availability, whereas areas where forage availability changes asynchronously (and at a sufficiently fine grain) may support more animals (Ash et al. 2004). The importance of variability between parcels in determining overall population responses is an important area for future research.
My focus is on testing general hypotheses related to sensitivity to fragmentation, but these results are most directly applicable to the three African regions modeled:
-
Landscapes in the commercial districts of the North-West Province are highly fragmented, but they are managed intensively. Management, such as rotation grazing, supplemental feeding, or other inputs from outside the system can disconnect the link between primary and secondary productivity, and reduce or eliminate sensitivity to fragmentation. In contrast, communal livestock owners graze their animals close to the few water sources within their districts, and more distant areas go ungrazed—the results I report here apply to other pathways to reduced mobility beyond fragmentation, such as sedentarization of pastoralists around water points (Lange 1985; James et al. 1999; Niamir-Fuller and Turner 1999; FAO 2001). Climate has become drier and more variable in South Africa (Mason et al. 1999). Should that pattern continue (i.e., in Fig. 5, <400 mm and >35% CV, which was observed), this work suggests that the sensitivity of the North-West Province cattle to fragmentation is not likely to change markedly.
-
Fragmentation is ongoing in Kajiado District, Kenya, as group ranches are subdivided into individually owned parcels. The sensitivities of livestock populations to fragmentation there are mixed, but areas of moderate or low productivity will support up to 25% fewer animals when the sites are fragmented (Boone et al. 2005; this work). If climate change causes precipitation to decline and become more variable (Easterling et al. 2000) (i.e., in Fig. 6, <500 mm and >30% CV, about what was observed), modest changes in the sensitivity of cattle populations to fragmentation in the area modeled may be expected. That said, small declines in the number of cattle that could be supported on lands would have profound effects upon food security of Maasai (Thornton et al. 2006).
-
Ngorongoro is a conservation area and therefore is not considered to be at risk from fragmentation into parcels. There are some concerns about reduced mobility through cultivation in the area affecting wildlife, but they appear misplaced (Boone et al. 2006). Maasai are unable to graze their livestock in the Serengeti Plains during the late wet season due to the risk of disease spread by wildebeest (Connochaetes taurinus) (McCabe 1995; Mduma et al. 1999), and tick-borne diseases are prevalent in the highlands. Reduced mobility due to disease risks is straining the ability of the area to support livestock. If climate change causes precipitation to decline and become more variable (Easterling et al. 2000) (i.e., in Fig. 7, <800 mm and >20% CV that was observed), Ngorongoro cattle may be far more sensitive to reduced mobility.
The model applications I used are reasonable representations of the ecosystems they reflect, and have supported published research (e.g., Boone et al. 2002, 2004, 2005; Galvin et al. 2004; Thornton et al. 2003, 2006). There remain caveats that must be considered when interpreting these results. The simulations do not provide predictions as to how fragmentation will alter cattle stocking 40 years hence. Innumerable changes to ecosystems and policies will occur over that time that cannot be accounted for. However, results do show the magnitude and direction of change due to fragmentation, all else being equal. Analyses that used annual rainfall spanning from 100 to 1,000 mm and 0 to 60% inter-annual CVs by their nature exceeded the natural range of rainfall and its variability in the study areas. The implications of this, such as changes in cattle populations and shrub cover, are cited above. Predators were not included in these models, which can alter cattle population dynamics (Davis et al. 2002), nor were browsing animals, which would have used the shrub resource that expanded under heavy rainfall. In general, the caveats cited in the original works remain applicable to this work.
I have shown that a linear decline in parcel area can lead to an accelerated (non-linear) decline in cattle populations, and that the decline is dependent on the amount and variability of precipitation and heterogeneity of landscapes. Regarding hypotheses, I showed that low precipitation made cattle populations relatively insensitive to fragmentation. High precipitation reduced sensitivity to fragmentation as well, but caused shrubs to expand. Increased inter-annual variability in precipitation decreased sensitivity to fragmentation when annual precipitation was above the observed average, but the opposite was true when precipitation was below the long-term average. The messages for policy makers, land managers, and others from this work are that: different landscapes have different sensitivities to fragmentation; fragmentation in sensitive landscapes will require more intensive inputs (e.g., rotation grazing, supplemental feeding) to offset declines in forage access; and changes in the severity of extreme weather events associated with climate change (Mason et al. 1999; Easterling et al. 2000) can alter the sensitivity of landscapes to fragmentation. If landscapes are fragmented due to policies or land tenure, efforts should be made to allow for the greatest mobility possible by animals (Ellis et al. 1993).
References
Andren H (1994) Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71:355–366
Ash AJ, Stafford Smith DM (1996) Evaluating stocking rate impacts in rangelands: animals don’t practice what we preach. Range J 18:216–243
Ash A, Gross J, Stafford-Smith M (2004) Scale, heterogeneity and secondary production in tropical rangelands. Afr J Range Forage Sci 21:137–145
Bekure S, de Leeuw PN, Grandin BE, Neate PJH (eds) (1991) Maasai herding: analysis of the livestock production system of Maasai Pastoralists in eastern Kajiado District, Kenya. Addis Ababa, Ethiopia: ILCA (International Livestock Centre for Africa), ILCA Systems Study 4
Boone RB (2000) Integrated management and assessment system training manual. Manual for IMAS, Global Livestock Collaborative Research Support Program, US Agency for International Development, University of California, Davis
Boone RB (2005) Quantifying changes in vegetation in shrinking grazing areas. Conserv Soc 3:150–173
Boone RB, Hobbs NT (2004) Lines around fragments: effects of fragmentation on large herbivores. Afr J Range Forage Sci 21:79–90
Boone RB, Wang G (2007) Cattle dynamics in African grazing systems under variable climate. J Arid Environ 70:495–513
Boone RB, Coughenour MB, Galvin KA, Ellis JE (2002) Addressing management questions for Ngorongoro Conservation Area using the Savanna Modeling System. Afr J Ecol 40:138–150
Boone RB, Galvin KA, Coughenour MB, Hudson JW, Weisberg PJ, Vogel CH, Ellis JE (2004) Ecosystem modeling adds value to a South African climate forecast. Clim Change 64:317–340
Boone RB, BurnSilver SB, Thornton PK, Worden JS, Galvin KA (2005) Quantifying declines in livestock due to land subdivision in Kajiado District, Kenya. Range Ecol Manage 58:523–532
Boone RB, Galvin KA, Thornton PK, Swift DM, Coughenour MB (2006) Cultivation and conservation in Ngorongoro Conservation Area, Tanzania. Hum Ecol 34:809–828
Boone RB, BurnSilver SB, Worden JS, Galvin KA, Hobbs NT (in press) Large-scale movements of large herbivores: livestock following changes in seasonal forage supply. In: van Langeveld F, Prins HHT (eds) Resource ecology. Kluwer Publishing, Dordrecht
Coe MJ, Cumming DH, Phillipson J (1976) Biomass and production of large African herbivores in relation to rainfall and primary production. Oecol 22:341–354
Conrad V (1941) The variability of precipitation. Month Weather Rev 69:5–11
Coughenour MB (1985) Graminoid responses to grazing by large herbivores: adaptations, exaptations, and interacting processes. Ann MO Bot Gard 72:852–863
Coughenour MB (1992) Spatial modelling and landscape characterization of an African pastoral ecosystem: a prototype model and its potential use for monitoring drought. In: McKenzie DH, Hyatt DE, McDonald VJ (eds) Ecological indicators, vol. 1. Elsevier Applied Science, New York, pp 787–810
Cushman SA (2006) Effects of habitat loss and fragmentation on amphibians: a review and prospectus. Biol Conserv 128:231–240
Davis SA, Pech RP, Catchpole EA (2002) Populations in variable environments: the effect of variability in a species’ primary resource. Philos Trans R Soc Lond B 357:1249–1257
Doncaster CP (2001) Healthy wrinkles for population dynamics: unevenly spread resources can support more users. J Anim Ecol 70:91–100
East R (1984) Rainfall, soil nutrient status and biomass of large African savanna mammals. Afr J Ecol 22:245–270
Easterling DR, Evans JL, Groisman PY, Karl YR, Kunkel KE, Ambenje P (2000) Observed variability and trends in extreme climate events: a brief review. Bull Am Met Soc 81:417–425
Eastman JL, Coughenour MB, Pielke RA Sr (2001) The regional effects of CO2 and landscape change using a coupled plant and meteorological model. Global Change Biol 7:797–815
Ellis J, Coughenour MB (1998) The Savanna integrated modelling system. In: Squires V, Sidahmed A (eds) Drylands: sustainable use of rangelands in the twenty-first century. IFAD technical reports, Rome, pp 97–106
Ellis J, Galvin KA (1994) Climate patterns and land-use practices in the dry zones of Africa. Bioscience 44:340–349
Ellis JE, Swift DM (1988) Stability of African pastoral ecosystems: alternate paradigms and implications for development. J Range Manage 41:450–459
Ellis JE, Coughenour MB, Swift DM (1993) Climate variability, ecosystem stability, and the implications for range and livestock development. Rethinking range ecology: implications for rangeland management in Africa. In: Behnke RH, Scoones I, Kerven C (eds) Range ecology at disequilibrium: new models of natural variability and pastoral adaptation in African savannas. Overseas Development Institute, London
Ewers RM, Didham RK (2005) Confounding factors in the detection of species responses to habitat fragmentation. Biol Rev 81:147–142
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Ann Rev Ecol Evol Syst 34:487–515
FAO (Food, Agriculture Organization of the United Nations) (2001) Pastoralism in the new millennium. FAO, Rome
Forman RTT (1995) Land mosaics. Cambridge University Press, Cambridge
Galvin KA, Thornton PK, Boone RB, Sunderland J (2004) Climate variability and impacts on East African livestock herders: the Maasai of Ngorongoro Conservation Area, Tanzania. Afr J Range Forage Sci 21:183–189
Galvin KA, Thornton PK, de Pinho JR, Sunderland J, Boone RB (2006) Integrated modeling and its potential for resolving conflicts between conservation and people in the rangelands of East Africa. Hum Ecol 34:155–183
James CD, Landsberg J, Morton SR (1999) Provision of watering points in the Australian arid zone: a review of effects on biota. J Arid Env 41:87–121
Lange RT (1985) Spatial distributions of stocking intensity produced by sheep flocks grazing Australian chenopod shrublands. Trans R Soc South Australia 109:167–174
Lewontin RC, Cohen D (1969) On population growth in a randomly varying environment. Proc Natl Acad Sci USA 62:1056–1060
Ludwig JA, Coughenour MB, Liedloff AC, Dyer R (2001) Modelling the resilience of Australian savanna systems to grazing impacts. Environ Int 27:167–172
Mason SJ, Waylen PR, Mimmack GM, Rajaratnam B, Harrison JM (1999) Changes in extreme rainfall events in South Africa. Clim Change 41:249–257
McCabe JT (1995) Wildebeest/Maasai interactions in the Ngorongoro Conservation Area of Tanzania. Final report (Grant No. 4953-93) submitted to the National Geographic Society, Washington
Mduma SAR, Sinclair ARE, Hilborn R (1999). Food regulates the Serengeti wildebeest: a 40-year record. J Anim Ecol 68:1101–1122
Niamir-Fuller M, Turner MD (1999) A review of recent literature on pastoralism and transhumance in Africa. In: Niamir-Fuller N (ed) Managing mobility in African rangelands: the legitimization of transhumance. Intermediate Technology Publications Ltd, London
Nicholson SE (1980) The nature of rainfall fluctuations in sub-tropical West Africa. Month Weather Rev 108:473–487
Phillipson J (1975) Rainfall, primary production and ‘carrying capacity’ of Tsavo National Park (East), Kenya. East Afr Wildl J 13:171–201
Ritchie M, Olff H (1999) Spatial scaling laws yield a synthetic theory of biodiversity. Nature 400:557–560
Saunders DA., Hobbs RJ, Margules CR (1991) Biological consequences of ecosystem fragmentation: a review. Cons Biol 5:18–32
Thornton PK, Galvin KA, Boone RB (2003) An agro-pastoral household model for the rangelands of East Africa. Agric Syst 76:601–622
Thornton PK, Fawcett RH, Galvin KA, Boone RB, Hudson JW, Vogel CH (2004) Evaluating management options that use climate forecasts: modeling livestock production systems in the semi-arid zone of South Africa. Clim Res 26:33–42
Thornton PK, BurnSilver SB, Boone RB, Galvin KA (2006) Modelling the impacts of group ranch subdivision on agro-pastoral households in Kajiado, Kenya. Agric Syst 87:331–356
Acknowledgments
This research was supported by the US National Science Foundation Biocomplexity program under grant number 0119618 to N.T. Hobbs et al., known as the SCALE project. Discussions with SCALE project personnel improved the manuscript, and comments from two anonymous reviewers and the Editor were helpful.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Boone, R.B. Effects of fragmentation on cattle in African savannas under variable precipitation. Landscape Ecol 22, 1355–1369 (2007). https://doi.org/10.1007/s10980-007-9124-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-007-9124-4