
Abstract
It has been proposed that high rates of risk-taking in adolescence are partly attributable to patterns of neurobiological development that promote an increase in sensation-seeking tendencies at a time when impulse control is still developing. It is not known, however, whether this pattern is the same for males and females. The present study investigates sex differences in the developmental trajectories of self-reported impulse control and sensation-seeking between the ages of 10 and 25 using longitudinal data from the National Longitudinal Study of Youth 1979 Child and Young Adult Survey (N = 8,270; 49 % female; 33 % Black, 22 % Hispanic, 45 % Non-Black, Non-Hispanic). Prior work has found that, consistent with the dual-systems model of adolescent neurobiological development, sensation-seeking rises and falls across this age span, whereas impulse control increases into the 20s. In the present study, we find that this same general pattern holds for both males and females, but with some key differences. As expected, males exhibit higher levels of sensation-seeking and lower levels of impulse control than females. Differences also emerged in the shapes of the developmental trajectories. Females reach peak levels of sensation-seeking earlier than males (consistent with the idea that sensation-seeking is linked to pubertal development) and decline in sensation-seeking more rapidly thereafter. Also, males increase in impulse control more gradually than females. Consequently, sex differences in both impulse control and sensation-seeking increase with age. The findings suggest that the window of heightened vulnerability to risk-taking during adolescence may be greater in magnitude and more protracted for males than for females.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Adolescence has long been regarded as a period of poor decision-making and rash behavior. This perception of adolescence is reinforced by empirical data surveying a range of behaviors, such as illicit drug use (Monitoring the Future 2009), reckless driving (Chen et al. 2000; National Highway Traffic Safety Administration 2007), unsafe sex (Finer 2010), and criminal activity (Piquero 2008). The dual-systems model of adolescent brain development (Casey et al. 2011; Steinberg 2008) offers an account of the observed age-related pattern in risky behavior. According to this model, patterns of development in two neurobiological systems render adolescence a time of relatively high sensation-seeking (the inclination to pursue exciting, novel or emotionally intense experiences) and low impulse control (the capacity to resist an urge to act). Though this model has generated a good deal of research, there has been little discussion or investigation of the possibility of sex differences in the development of the relevant brain systems and the behaviors that they subserve. Yet, there are empirical and theoretical reasons to suspect sex differences in developmental trajectories of sensation-seeking and impulse control. The present study addresses this lacuna, using data from a large-scale, longitudinal study of American adolescents and young adults.
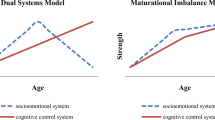
The dual-systems model of adolescent neurobiological development views risk-taking as the product of an imbalance between two brain systems: an incentive processing system and a cognitive control system (Casey et al. 2008; Steinberg 2008). The incentive processing system responds to rewarding stimuli and spurs sensation-seeking behaviors. This system undergoes rapid change in early adolescence, which orients the individual to “adult” social motivations (like attaining social status and sexual partners), and its influence rises and then falls across the adolescent period. The cognitive control system, which undergirds self-regulatory behavior, such as impulse control, follows a different pattern of development, increasing in strength gradually from childhood into young adulthood. The observed adolescent peak in risky behavior is thought to emerge due to the remodeling and heightened activation of the incentive processing system prior to the full maturation of the cognitive control system (Steinberg 2010). This theoretical model has received support from multiple fields of developmental research, including behavioral studies, neuroimaging studies, neurophysiological studies, and animal models [see Casey et al. (2011), Spear (2013), and Steinberg (2008) for reviews of the empirical data supporting the dual-systems theory; see Pfeifer and Allen (2012) for a critique of the theory; and see Strang, Chein and Steinberg (2013) for a rejoinder to that critique].
The psychological traits of sensation-seeking and impulse control are thought to reflect the functional status of the incentive processing and cognitive control systems, respectively. Both traits (high sensation-seeking and low impulse control) appear to contribute to risk-taking behavior (e.g. Dahlen et al. 2005; Donohew et al. 2000; Harden et al. 2012; Quinn and Harden 2013; Zuckerman and Kuhlman 2000). Consistent with the dual-systems model, cross-sectional behavioral (e.g., Romer et al. 2010; Steinberg et al. 2009) and neuroimaging studies (see Casey et al. 2011 for a review) find evidence that sensation-seeking rises during adolescence and then falls in early adulthood, whereas impulse control (and similar constructs) increases monotonically across this same span. Similar patterns emerge in longitudinal analyses. For example, Harden and Tucker-Drob (2011) used data from the same large-scale study employed in the present article to investigate age-related patterns across adolescence and early adulthood in impulse control and sensation-seeking. Their analysis, which included observations spanning ages 12–25, tested several alternative models to characterize the patterns of growth in these constructs. They found that sensation-seeking rose in early adolescence and then declined, whereas impulsivity (i.e., the opposite of impulse control) fell steadily across the age range examined.
Absent from the extant literature is rigorous investigation of whether and how the development of sensation-seeking and impulse control differs for males and females. The fact that males outnumber females in various forms of risky behavior [e.g., fatal accidents (Heron 2012), gambling (Zuckerman and Kuhlman 2000), crime (Federal Bureau of Investigation 2010; Moffitt et al. 2001; Shulman et al. 2013)] is suggestive of sex differences in the levels of traits such as sensation-seeking and/or impulse control. And indeed, there is ample evidence of sex differences in the levels of these traits (e.g., Cross et al. 2013; Moffitt et al. 2013; Schmitt et al. 2008, although there are exceptions, e.g., Patton and Stanford 1995). For example, in two recent meta-analyses, Cross and colleagues found that, across studies, males scored higher than females on measures of sensation-seeking and impulsivity (Cross et al. 2011, 2013).
There are several theoretical reasons to expect sex differences in impulse control and sensation-seeking and in the development of neurobiological systems that underlie their expression. One is the different evolutionary pressures on males and females. Theories of sexual selection argue that women value high social status in male sexual partners because status signals the ability to provide resources and offer protection (Ellis 1992). To attain high social status, men often need to take risks (Wilson and Daly 1985). Thus, high levels of sensation-seeking and low levels of impulse control may have evolved in males as a result of the reproductive success of men who were psychologically predisposed to engage in the sorts of risk-taking that confer high status. Furthermore, because risk-taking poses a threat to the organism, it also seems plausible that the psychological predisposition toward risk-taking in males is experience-sensitive, such that it declines once one’s social position and/or sexual access to females is well-established (Daly and Wilson 2001).
Evolutionary theory also provides a reason to anticipate that sensation-seeking will increase in adolescence for females. Even if females do not face evolutionary pressures identical to males’ with respect to risk-taking, the remodeling of the incentive processing system in early adolescence may, for females, facilitate the motivation to move out of the family circle (which aids in avoiding incest) and pursue sexual interests, both of which involve some degree of risk (Spear 2000). The timing of this rise may not correspond with the age at which today’s young women actually move out of their parents’ homes (although it may have done so during earlier periods of human history). Rather, the rise in sensation-seeking in females may correspond with the transition to having more of one’s important social interactions occur outside of the home and placing greater emphasis on relationships with peers and romantic partners. Thus, to the extent that a rise in sensation-seeking is indicative of developmental changes within the incentive processing system, we would expect some increase in this trait in adolescent females.
Furthermore, if the timing of the remodeling of the incentive processing system is tied to puberty—and there is growing evidence that this is the case (Dahl 2004; Forbes and Dahl 2010; Smith et al. 2013; Steinberg et al. 2008)—we would expect the rise in sensation-seeking to occur earlier for females than for males, because girls on average go through puberty about 18 months earlier than boys. Indeed, a recent study found that pubertal development, over and above chronological age, predicts greater reward-seeking and sex-specific volumetric changes in the nucleus accumbens, a brain region that is sensitive to reward (Urošević et al. 2014). Other studies have found that, among adolescents, higher levels of testosterone (a pubertal hormone) predicts greater activation of brain regions sensitive to reward during reward trials (relative to loss trials) in a gambling task (Op de Macks et al. 2011). These studies reinforce the view that pubertal development spurs changes in the incentive processing system. To the extent that longitudinal trajectories of sensation-seeking reflect puberty-linked changes in the incentive processing system, the pattern for girls should be shifted earlier than the pattern for boys because of the sex difference in pubertal timing. Moreover, if the effects of pubertal development on incentive processing are at least partially mediated by increases in testosterone, as suggested by Op de Macks et al. (2011), this suggests possible sex differences in the magnitude of incentive processing changes, as puberty is associated with much larger rises in testosterone among males than among females (Shirtcliff et al. 2009).
Unlike sensation-seeking, impulse control is thought to develop relatively independently of puberty (see Smith et al. 2013). Thus, the existence of sex differences in the age of pubertal onset does not provide a rationale for expecting sex differences in the development of impulse control. As for the evolutionary perspective, the proposed pattern of prolonged readiness for risk-taking in males (relative to females) could be explained by low levels of impulse control, but it could also be accounted for by high levels of sensation-seeking. Therefore, neither evolutionary theory nor the evidence of the developmental effects of puberty provides a compelling rationale for predicting slower growth or lower levels of impulse control in males than females. Indeed, the few studies that have examined age differences in impulse control across a wide age span and compared males and females do not find evidence that the rate of increase varies as a function of sex (e.g., Steinberg et al. 2008; Galvan et al. 2007). Also, in the meta-analysis by Cross et al. (2011), the evidence for lower impulse control in males was weaker than the evidence for greater sensation-seeking. Again, however, longitudinal research is lacking.
Finally, it is also possible that socialization processes contribute to sex differences in the development of sensation-seeking and impulse control and in consequent risk-taking. For instance, if bravery is more highly prized among males than among females within a given culture, or if exhibitions of poor impulse control are less tolerated among females than among males, then gender-specific reinforcement and punishment of sensation-seeking and impulse control could produce disparities between males and females in these traits. However, while cultural factors may constrain the expression of psychological traits that promote or inhibit risk-taking, it is not known whether these cultural factors actually alter the development of these traits. A recent study of adolescents in five different countries did find differences in levels of “low self-control,” a measure that combined elements of sensation-seeking and (low) impulse control (Vazsonyi and Belliston 2007), consistent with the notion that cultural factors can affect the development of these traits. Still, though there may be cultural variation in levels of sensation-seeking and impulse control, the fact that disproportionate risk-taking by males is seen across cultures (Wilson and Daly 1993) suggests that there is some biological basis for sex differences in risk-taking, which may be exacerbated to different degrees within different cultural contexts. Though we do not, in the present study, investigate the role of culture in producing sex differences in sensation-seeking or impulse control, we are cognizant that cultural factors, as well as biological factors, may have contributed to any observable sex differences.
Current Study
The goal of the present study was to characterize sex differences in the development of sensation-seeking and impulse control during adolescence and early adulthood. Based on observed sex differences in risk-taking, and prior findings of sex differences in impulse control and sensation-seeking in cross-sectional studies, we expected to find that males exhibit higher levels of sensation-seeking and lower levels of impulse control than females across the full age range examined. In addition to testing this specific hypothesis, we conduct analyses to assess whether the disparity between females and males in these constructs changes with age. Of course, many environmental and biological factors (e.g., peer behavior, poverty, non-sex-linked genes) may have greater influences on impulse control and sensation-seeking than sex. We therefore expected only modest sex differences in sensation-seeking and impulse control—particularly with respect to differences in the shapes of the developmental trajectories (as opposed to the levels). Even small effects, however, can help to inform theories of behavior and may contribute to sex disparities in morbidity and mortality when operating across an entire population. We also predicted, based on evidence of a link between puberty and sensation-seeking, that males and females would differ with respect to age-related patterns of sensation-seeking (but not impulse control). Specifically, while we anticipated a rise in sensation-seeking during early adolescence for both sexes, we predicted that the onset or peak of this rise would occur earlier in development for females because puberty begins earlier among females than males. Finally, based on the dual-systems model, we expected to find that sensation-seeking is highest relative to impulse control during early to mid-adolescence for both sexes.
Our analyses build on and extend those reported by Harden and Tucker-Drob (2011), a longitudinal analysis that drew on the same data set as the present study. There are five key differences between our analysis and that of Harden and Tucker-Drob: (1) we analyzed the latent growth curves separately for males and females, in order to test hypotheses about sex differences in the development of these traits, (2) we considered more complicated growth patterns (e.g., models specifying cubic patterns of growth), (3) we included newer releases of the data, (4) we expanded the age range to include the 10–11 year old age group, to better observe the onset of adolescent patterns of growth in sensation-seeking and impulse control, and (5) we retained the original metrics of the scale on which sensation-seeking and impulse control were measured rather than reporting standardized residuals, in order to directly compare levels of sensation-seeking and impulse control.
Methods
Sample
Data from the National Longitudinal Study of Youth 1979 Child and Young Adult Survey (CNLSY) were analyzed. CNLSY respondents are the biological children of women surveyed in the NLSY79, a longitudinal study that began in 1979, when the NLSY79 sample was 14–22 years old (see www.bls.gov/nls/nlsy79.htm for details). The CNLSY includes a large sample followed longitudinally across adolescence and early adulthood, which makes it an optimal data set in which to address our research questions. Our sample consisted of 8,270 individuals in the CNLSY (49 % female, 33 % Black, 22 % Hispanic, 45 % Non-Black, Non-Hispanic) who provided data on impulse control and sensation-seeking at least once during the biennial surveys that took place between 1994 and 2010. The response rate was 95 % at the initial survey and retention was high (about 90 %) through 2006 [see Harden and Tucker-Drob (2011) and Quinn and Harden (2013) for further detail on missing data]. For our analysis, we focused on responses recorded at ages 10 through 25. Because respondents were surveyed biennially, data were organized by age in 2-year groupings (see Table 1). Of the eight potential measurement occasions within this age span, most participants provided data for four (30.4 %) or five (30.0 %) (M = 3.42, SD = 1.28). Smaller percentages provided data at one (2.6 %), two (10.3 %), or six (8.3 %) measurement occasions, and none provided data on more than six occasions. Our statistical analyses included data from all the selected respondents, regardless of the number of waves in which they participated. This is appropriate because our statistical models are estimated using full information maximum likelihood estimation (FIML), which takes advantage of all available data to yield more precise estimates and reduce bias due to missing data (Raykov 2005). To aid in this estimation, our models incorporated control variables that are correlated with data missingness (as described below).
Measures
Sociodemographic Variables
Sex was used as an independent variable in our models except in sex-specific analyses. We also controlled for respondent’s race [categorized as Black, Hispanic, or neither (the reference category)] as well as the age of the respondent’s mother when the respondent was born (maternal birth age). Several other characteristics of respondents’ mothers were used to gauge socioeconomic status. These were maternal income at age 30, which included wages and government support (log-transformed); maternal educational attainment at age 30; and maternal intelligence, assessed in 1980 using the Armed Services Vocational Aptitude Battery (specifically, a composite score for word knowledge, paragraph comprehension, math knowledge, and arithmetic reasoning). (For purposes of analysis, the percentile scores from this measure were centered on their mean and divided by 10.) Continuous variables were centered on their means for analysis. See Table 1 for descriptive statistics.
To the extent that these covariates were correlated with data “missingness,” they helped to reduce bias due to missing data in the full information maximum likelihood estimation of our statistical models. Maternal birth age was an especially important correlate of data missingness because respondents born to younger mothers were able to contribute more data to the study. To illustrate, imagine two respondents whose mothers were born in 1960. One respondent, born in 1998 (when her mother was 38 years old), would have had the opportunity to provide responses only at age 10 (during the 2008 data collection) and age 12 (during the 2010 data collection), whereas a second respondent, born in 1985 (when her mother was 25 years old), would have had the opportunity to provide data eight times—every other year from age 11 (during the 1996 data collection) to 25 (during the 2010 data collection). The correlation between maternal birth age (i.e., the age of the respondent’s mother when the respondent was born) and number of waves of data provided was r = −.21, p < .001. Number of waves of participation was also associated (albeit much more weakly) with maternal income (r = −.04, p < .001) and maternal educational attainment (r = −.03, p < .01), but not with maternal intelligence or race/ethnicity.
Impulse Control and Sensation-Seeking
We operationalized the key constructs using the same variables as Harden and Tucker-Drob (2011). Impulsivity was assessed with three items: (1) “I often get in a jam because I do things without thinking,” (2) “I think that planning takes the fun out of things,” and (3) “I have to use a lot of self-control to keep out of trouble.” Sensation-seeking was also assessed with three items: (1) “I enjoy taking risks,” (2) “I enjoy new and exciting experiences, even if they are a little frightening or unusual,” and (3) “Life with no danger in it would be too dull for me.” These six items for impulsivity and sensation-seeking, respectively were included in the self-administered assessment and were rated on a 4-point scale ranging from strongly agree to strongly disagree. The three items comprising each scale were averaged. We reverse-coded scores for impulsivity so that higher scores indicated greater impulse control.
Prior analysis of data from this study indicates that these six items represent two distinct constructs, and that the two scales they form are correlated with measures of risky behavior in the expected directions: positively for sensation-seeking and negatively for impulse control. Specifically, a slower decline in sensation-seeking (after age 15) is associated with increasing use of alcohol, and slower growth in impulse control is associated with increasing use of alcohol, marijuana and cigarettes (Quinn and Harden 2013). Also, upward change in sensation-seeking is correlated with upward change in delinquency (Harden et al. 2012). These findings provide evidence of the predictive validity of the measures. Furthermore, as reported by Harden and Tucker-Drob (2011), the measure of impulse control is significantly correlated with concurrent measures of the “Big Five” personality traits of Conscientiousness (r = .28) and Emotional Stability (i.e., low Neuroticism, r = .32), assessed with the Ten-Item Personality Inventory (Gosling et al. 2003), which suggests that it exhibits convergent validity with related constructs.
The impulse control and sensation-seeking scales exhibited an acceptable degree of model fit and measurement invariance across age group and sex. [Responses from the most recent complete wave of data collection (N = 6,988) were used to assess model fit and measurement invariance.] Confirmatory factor analyses (conducted separately for each construct), in which the impulse control or sensation-seeking items served as indicators of a latent factor, and in which item variances were constrained to be equal (because, otherwise, the models are exactly identified and do not yield fit statistics), found that model fit was acceptable for both measures according to the Confirmatory Fit Index (CFI) and Root Mean Square Error of Approximation (RMSEA), commonly used measures of model fit that are explained further in the Analytic Plan: Δχ 2 (2) = 36.37, p < .001, CFI = .97, RMSEA = .05 for impulse control; and Δχ 2 (2) = 10.26, p < .01, CFI = 1.00, RMSEA = .02 for sensation-seeking. Measurement invariance was established by estimating confirmatory factor analyses for impulse control and sensation-seeking (separately) while treating age group and then sex (serially) as different groups to be compared. In the unrestricted models, the factor loadings were allowed to vary across age group or sex; in the restricted models, the loadings were constrained to be equal across age group or sex. The restricted models did not fit worse than the unrestricted models, a result that indicates measurement invariance [for age group invariance, Δχ 2 (20) = 23.24, p = ns for impulse control and Δχ 2 (20) = 53.39, p = ns for sensation-seeking; for sex invariance, Δχ 2 (2) = 4.60, p = ns for impulse control and Δχ 2 (2) = 2.52, p = ns for sensation-seeking].
Analytic Plan
To address our research questions, we analyze the data in five steps:
-
1.
We provide basic descriptive information on impulse control and sensation-seeking by age and sex.
-
2.
We test a series of latent growth curve models in order to examine whether the functional form of change in impulse control and sensation-seeking differs for males and females.
-
3.
We assess the degree to which the trajectories of impulse control and sensation-seeking vary by sex.
-
4.
We further characterize how the gap between males and females in each construct change with age.
-
5.
We estimate the degree to which the gap between the level of sensation-seeking and the level of impulse control varies by age, within each sex.
All our statistical models were estimated in Mplus version 5.21 (Muthén and Muthén 2009), using full information maximum likelihood. Because some of the covariates (especially maternal birth age) were known to be correlated with data missingness, we included the covariates in all the models to help assure accurate estimation. Also, because some participants were siblings, who might provide more similar responses than would non-siblings, our models adjust for the non-independence of observations “clustered” within families (Asparouhov and Muthen 2006). Our models were estimated using the MLR estimator in Mplus, which is appropriate for clustered data. As a result, when we compared nested models to test our hypotheses, we used the Satorra-Bentler scaled Chi square difference test, which is the recommended approach for models estimated with MLR (Muthén and Muthén 2009).
Results
Sex Differences in the Marginal Means of Impulse Control and Sensation-Seeking
Descriptive statistics for the study variables are reported in Table 1. Marginal means for impulse control and sensation-seeking (adjusted for the control variables) are reported in Fig. 1a. To obtain the estimates of the marginal means, we regressed each observed measure of impulse control and sensation-seeking (simultaneously) on the control variables with no other structure imposed. The effect of sex (i.e., the effect of being female relative to male) was significant for every 2-year age grouping. Marginal means by sex are reported in Table 2 and Fig. 1b, c. At every age, females were significantly higher in impulse control and lower in sensation-seeking (see Fig. 1b, c). The magnitude of the difference was relatively modest, ranging from 0.08 to 0.17 standard deviations for impulse control and from −0.09 to −0.22 standard deviations for sensation-seeking.

a Estimated marginal means (EMMs) for impulse control and sensation-seeking (adjusted for control variables and sex). Error bars represent standard errors. b EMMs by sex for impulse control. c EMMs by sex for sensation-seeking. As noted in Table 2, the differences between males’ and females’ EMMs (in b, c) are significant at every age
To test whether these sex differences varied by age, we compared the freely estimated marginal means models to models in which the effect of sex on the repeated measures of impulse control or sensation-seeking was constrained to be equal across all age groups. For both impulse control and sensation-seeking, this omnibus test indicated that the magnitude of the sex difference varied across age [scaled Δχ 2 (7) = 33.37, p < .001 for impulse control; scaled Δχ 2 (7) = 38.17, p < .001 for sensation-seeking]. Visual inspection of the marginal means suggested that the sex differences increased with age, at least after early adolescence. To test whether this was the case, we compared the fully constrained models (sex differences equal for every age) to models that allowed the sex difference to be different for the 18–25 year olds than for the 10–17 year olds. In both cases, this model fit better than the fully constrained model, indicating that, for both impulse control and sensation-seeking, the estimated effect of sex was greater for the 18–25 age range than for the 10–17 age range [B younger = 0.10 (SE = 0.01) versus B older = 0.16 (SE = 0.01), scaled Δχ 2 (1) = 14.97, p < .001, for impulse control; B younger = −0.14 (SE = 0.01) versus B older = −0.21 (SE = 0.01), scaled Δχ 2 (1) = 25.12, p < .001, for sensation-seeking], suggesting that sex disparities do increase with age.
Sex-Specific Growth Curve Models
Our next aim was to estimate whether the functional form of change in impulse control and sensation-seeking differed by sex. To find out, we estimated series of latent growth curve models (McArdle and Nesselroade 2002), each specifying a different possible form of growth. The series of models specified included no growth (intercept only), linear growth (intercept and slope), quadratic growth (which allows for a single change in the rate of growth), cubic growth (which allows for two changes in the rate of growth), and latent basis (which does not impose a shape, but rather allows for data-driven estimation of the shape of the growth curve). The results are reported in Table 3 and the formulae for these models are presented in the supplementary materials.
Critically, the series of models was estimated separately not only for each construct but also for each sex. To select the best fitting model in each series, we examined several widely-used fit statistics (reported in Table 3) and, where we could compare nested models to determine whether we could use a more parsimonious model (one with fewer freely estimated parameters and more degrees of freedom) without significantly hurting model fit, we did so. The fit statistics examined include the Confirmatory Fit Index (CFI), for which values closer to 1 and farther from 0 indicate better fit; the sample size adjusted Bayesian Information Criterion (SABIC), for which smaller values indicate a better balance of fit and parsimony; and the root mean square error of approximation (RMSEA), for which values closer to zero indicate a closer fit between the estimated and observed values. We also report, for each model, the Chi square statistic, the degrees of freedom, and the scaling correction factor for MLR, which are necessary to conduct Satorra-Bentler Chi square difference tests. When we encountered errors with model convergence suggestive of overfitting (i.e., too few model constraints), we proceeded by estimating models in which the residual variance term for the highest order growth parameter was fixed to zero, which imposes the assumption that the value of the that parameter is the same for all individuals. In all these models, the latent growth parameters (e.g., the level, slope, etc.) were regressed on the control variables.
For females, the (unrestricted) cubic models for impulse control and sensation-seeking did not converge properly, so estimates are not reported; the cubic models in which the residual variance term for the cubic parameter was fixed at zero (c@0) converged without error. For nested models, we conducted Chi square difference tests to assess which one fit best. For males, scaled Chi square difference tests indicated that the (more parsimonious) cubic c@0 model did not fit worse than the cubic model [Δχ 2 (4) = 5.39, p = ns, for impulse control; Δχ 2 (4) = 8.04, p = ns, for sensation-seeking], so the c@0 models were selected. For all four series, Chi square difference tests indicated that the (even more parsimonious) quadratic models fit significantly worse than the cubic c@0 models [Δχ 2 (7) = 25.57, p < .001, scaled Δχ 2 (7) = 87.83, p < .001, Δχ 2 (7) = 34.94, p < .001, Δχ 2 (7) = 121.58, p < .001, for males’ impulse control, females’ impulse control, males’ sensation-seeking, and females’ sensation-seeking, respectively]. Consistent with the Chi square difference tests, with only one exception, the fit statistics for all the series pointed to the cubic models as the best-fitting. None of the fit statistics favored the latent basis models. Together, these analyses led us to conclude that the cubic models (with the residual variance in the cubic parameter fixed to zero) best described the form of growth for impulse control and sensation-seeking for both sexes. The specification of these models is depicted visually in Fig. 2.Footnote 1

Simplified representation of the cubic c@0 model. For clarity of presentation, only the first three and the final age group (time point) are depicted. Rectangles represent observed variables and ovals represent latent variables. Single-headed arrows represent regression parameters. The labels on the regression lines represent the weights assigned to them in this model. The circles labeled “eL,” “eS,” and “eQ” represent the residual variance terms for the latent level, slope and quadratic terms, respectively. Because the cubic term’s residual variance was fixed at zero, it is not represented in this figure. The observed measure of the dependent variable (DV) at each time point also has a corresponding error variance term, though these are not depicted. Double-headed arrows represent covariance terms
The average trajectories described by the cubic models for impulse control and sensation-seeking for males and females are graphed in Fig. 3. Cubic models can accommodate two changes of direction or velocity. In the case of impulse control, for both sexes the average trajectory was one in which impulse control fell slightly from age 10–11 to a nadir in early adolescence and then rose through age 25, with the rate of growth tapering off near the high end of the age range. For sensation-seeking a different pattern was observed. For both males and females, sensation-seeking rose steeply from age 10–11 to late adolescence and then fell off in early adulthood. The decline in sensation-seeking in early adulthood appeared to be more pronounced for females than for males.

Estimated trajectories for impulse control (IC; solid lines) and sensation-seeking (SS; dashed lines) for females (grey lines) and males (black lines). The estimates for these four trajectories are derived from four separate cubic models (c@0)
Sex Differences in Trajectories of Impulse Control and Sensation-Seeking
Having established that a cubic pattern of growth best characterized the functional form of growth for sensation-seeking and impulse control for both males and females, our next step was to estimate the degree to which males and females differed in their average trajectories of impulse control and sensation-seeking. To do so, we estimated a bivariate cubic growth model for the combined sample (males and females) and regressed the latent growth parameters on sex (as well as on the control variables), enabling us to quantify the sex differences in the growth parameters, and hence the trajectories, of these constructs. This model included both sets of dependent variables (the repeated measures of impulse control and sensation-seeking). We allowed all covariances between the latent growth parameters (except the cubic ones, which had no variance) to be freely estimated and allowed the concurrent measures of impulse control and sensation-seeking to co-vary as well. The key estimates from this model, which provided a close fit to the data [Δχ 2 (155) = 227.60, p < .001, CFI = 0.99, RMSEA = 0.008], are reported in Table 4. A full reporting of the model estimates is available in the supplementary materials. To formally test whether the effects of sex on the growth parameters were significant, we examined the change in Chi square when the effects of sex were constrained to be zero (i.e., specifying no difference between males and females) versus when these effects were freely estimated. A significant increase in the Chi square statistic when the effect of sex is constrained to be zero indicates that the effect is significant.
Sex Differences in the Trajectory of Impulse Control
On average, females’ impulse control, centered at age 18–19, was substantially greater (by about 0.16 SD) than that of males. Growth in impulse control and acceleration in the rate of growth was significantly faster for females than males, as indicated by significant effects of sex on the linear and quadratic latent growth parameters. The effects of sex were modest, however, accounting for only 3, 4, and 1 % of the variation in the intercept, slope, and quadratic terms, respectively (over and above the control variables). When the effects of sex on the slope, quadratic, and cubic parameters for impulse control were set to zero, model fit was significantly worse [Δχ 2 (3) = 29.99, p < .001], confirming that the shape of the trajectory (not just its level) varied significantly by sex.
Sex Differences in the Trajectory of Sensation-Seeking
Males were higher overall than females in sensation-seeking (by 0.21 SD at age 18–19). The shapes of the female and male trajectories also differed, as evidenced by significant sex differences in the slope and cubic parameters. The estimates suggest that females (on average) experience a more dramatic decline in sensation-seeking in the transition from adolescence to early adulthood. Comparison to a model that posited no effect of sex revealed that these sex differences accounted for 3, 4, and 0 % of the variation in the intercept, slope, and quadratic terms respectively (over and above the control variables). When the effects of sex on the slope, quadratic, and cubic parameters for sensation-seeking were set to zero, model fit was significantly worse [Δχ 2 (3) = 37.62, p < .001], again confirming that the shape of the trajectory (not just its level) varied significantly by sex.
Sex Differences in Impulse Control and Sensation-Seeking by Age
Next, we wanted to examine how the gap between males and females in each construct changed with age. To do so, we first iteratively re-centered the growth models at each 2-year age grouping and assessed the estimated effect of sex (the difference between the female and male estimate) on the level of the construct at that age. Then, we formally evaluated whether the estimated effects of sex on the levels of the dependent variables differed at each age. This was accomplished by fixing the estimated effect of sex on the level at one age (e.g., age 14–15) to the value estimated for another age (e.g., age 16–17) and testing whether doing so significantly reduced the model’s fit. In this series of analyses, a significant decrement in model fit indicated that the effect of sex on the level differed for the two ages being compared.
The results (reported on the right side of Table 2) suggested that the disparity between males and females in the levels of sensation-seeking and impulse control was smallest during mid-adolescence (around age 14–15 or 16–17) and grew thereafter, with males becoming progressively higher in sensation-seeking and lower in impulse control than females. Rather than conduct pairwise comparisons of the sex difference in each construct at every age (which would require 56 comparisons), we tested (a) whether the sex difference in the level of each construct at age 14–15 was different from that at age 16–17 (because the sex difference was smallest at these two ages), and (b) whether the sex difference at age 14–15 was different from that at the extreme ends of the age range (age 10–11 and age 24–25). This second analysis revealed whether the apparent changes in the magnitude of the sex difference across the age span sampled—i.e., the shrinking and growing of the sex differences—were significant.
First, to assess whether the sex difference in the level of impulse control was the same at age 14–15 and age 16–17, we fixed the estimated effect of sex on the level at age 16–17 equal to 0.086, the estimated value of that parameter in the model in which age was centered at 14–15. Because fixing this parameter did not result in a decrement in model fit [Δχ 2 (1) = 0.27, p = ns], we could conclude that the effect of sex on the level of impulse control was not significantly different at these two ages. A parallel test for sensation-seeking yielded a similar result: the sex difference in the level at age 16–17 was not significantly different from that at age 14–15 [Δχ 2 (1) = 2.73, p = ns].
Having established that the estimated effects of sex on the levels of impulse control and sensation-seeking at age 14–15 were not different from those at age 16–17, we next tested whether they were different from those at age 10–11. For both impulse control [Δχ 2 (1) = 35.26, p < .001] and sensation-seeking [Δχ 2 (1) = 6.00, p < .05], the magnitude of the difference between males and females at age 10–11 was significantly greater than at age 14–15. We used a similar approach to test whether the magnitude of the sex differences in impulse control and sensation-seeking at age 24–25 was significantly greater than at age 14–15. The Chi square difference tests suggested that this was the case for both impulse control [Δχ 2 (1) = 66.78, p < .001] and sensation-seeking [Δχ 2 (1) = 77.22, p < .001]. Thus, while females are higher than males in impulse control across the full age range tested, and males are higher than females in sensation-seeking across the full age range tested, the magnitude of these gender gaps shrinks from preadolescence to mid-adolescence and then grows from mid-adolescence to early adulthood.
Next, we further explored the nature of the sex differences in the estimated average trajectories of impulse control and sensation-seeking by running linear latent growth curve models on restricted age ranges. These models tested whether apparent sex differences in the rates of change in sensation-seeking and impulse control across certain age spans were statistically significant.
Sex Differences in the Rise and Fall of Sensation-Seeking
Visual inspection of the average estimated trajectories for sensation-seeking for males and females suggested that the rise in sensation-seeking in adolescence (across ages 10–17) was similar for the two sexes, but that, during early adulthood, the decline in levels of sensation-seeking was more pronounced for females than for males. To test whether this was the case, we estimated two linear latent growth curve models (level and slope only), one that included the observations of sensation-seeking and impulse control between ages 10–11 and 16–17, and another that included the observations of these variables between ages 18–19 and 24–25. By examining the effects of sex on the slopes in these models, we were able to formally assess whether there were sex differences in the rate of increase in sensation-seeking in adolescence or in the rate of decline in sensation-seeking in early adulthood. Between ages 10 and 17, the effect of sex on the slope was not significant [B sex = 0.003, SE = 0.008, p = ns], meaning that the rate of growth in sensation-seeking was equivalent for males and females during this age span. However, between 18 and 25, there was a significant effect of sex on the slope [B sex = −0.023, SE = 0.009, p < .01], indicating that the rate of decline in sensation-seeking was more rapid for females than for males during this age span.
Sex Differences in the Fall and Rise of Impulse Control
Finally, visual inspection of the average estimated trajectories of impulse control for males and females suggested that levels of this construct fell slightly from age 10–11 to age 14–15 and then rose monotonically thereafter through age 24–25. To test whether there were sex differences in these patterns, we estimated two more linear latent growth curve models: one on the observations between 10 and 15 years and one on the observations between 16 and 25. Between 10 and 15, the rate of decline in impulse control was not significantly less than zero for males [B slope = −0.023, SE = 0.013, p > .05], but was more negative for females than for males [B sex = −0.029, SE = 0.012, p < .05]. Consequently, we reversed the coding of sex in order to be able to test whether the rate of decline in impulse control between 10 and 15 was significantly negative for females; the resulting model revealed that it was [B slope = −0.052, SE = 0.013, p < .001]. The model positing linear growth between 16 and 25 revealed that the rate of increase in impulse control during this age span was positive for males [B slope = 0.045, SE = 0.006, p < .001], but more so for females [B sex = 0.028, SE = 0.006, p < .001]. So, while females experience a modest decline in impulse control between ages 10 and 15—a drop that is not significant for males—they also outpace males in the development of impulse control between ages 16 and 25.
Age Differences in the Gap Between Sensation-Seeking and Impulse Control, by Sex
Because we were also interested in delineating the window of vulnerability for risk-taking, we next estimated the degree to which the gap between the level of sensation-seeking and the level of impulse control varied by age, within each sex. To explore this, we iteratively re-centered the latent growth curve models (using the full age range) at each 2-year age grouping, but this time we ran the models separately by sex. Then, we identified the point(s) at which sensation-seeking was maximal relative to impulse control by constraining the levels of each construct (at a given age) to be equal to one another in each model and assessing the decrement in model fit compared to the unconstrained model. To the extent that being high sensation-seeking relative to impulse control confers risk for reckless behavior, this final set of analyses helps to delineate the window of greatest vulnerability for risk-taking for each sex.
Examination of the marginal means and estimated growth trajectories for females suggested that sensation-seeking was higher than impulse control at ages 14–15 and 16–17, but just barely. Our formal test revealed that, for females, the level of sensation-seeking was not significantly higher than that of impulse control at either age 14–15 [Δχ 2 (1) = 1.00, p = ns] or 16–17 [Δχ 2 (1) = 0.19, p = ns]. In contrast, at the ages adjacent to 14–17, levels of impulse control were significantly higher than levels of sensation-seeking for females [Δχ 2 (1) = 14.46, p < .001 at age 12–13 and Δχ 2 (1) = 15.32, p < .001 at age 18–19]. At all other ages, the levels of impulse control were higher still than the levels of sensation-seeking. In sum, for females, there was no age at which sensation-seeking was significantly higher than impulse control, but for the period spanning 14–17, the levels of the two constructs were equivalent. This age period (between 14 and 17) therefore represents the time of peak vulnerability to risk-taking among females.
For males, at every age but 10–11, levels of sensation-seeking were higher than levels of impulse control. We pinpointed the period of males’ peak vulnerability to risk-taking (defined as maximal difference between levels of sensation-seeking and impulse control [where sensation-seeking > impulse control]) by constraining the estimated values for the levels of the two constructs to be equal at each age (except 10–11) and identifying the point at which doing so produced the greatest decrement in model fit. This series of tests revealed that the window of heightened vulnerability to risk-taking opened at age 12–13 (when sensation-seeking levels were first found to be significantly higher than levels of impulse control), peaked at age 16–17, and closed at age 24–25, the only age (other than 10–11) at which the Chi square difference test indicated that the level of sensation-seeking was not significantly higher than the level of impulse control [Δχ 2 (1) = 3.45, p = ns] for males. Between ages 12–13 and 22–23, the Chi square differences on one degree of freedom were: 24.69 (age 12–13), 99.64 (age 14–15), 121.38 (age 16–17), 65.97 (age 18–19), 23.46 (age 20–21), and 9.73 (age 22–23), all ps < .001, except age 22–23 for which p < .01. In other words, the window of heightened vulnerability to risk-taking appears to be open wider and for a longer period for males than for females.
Discussion
It is widely observed that adolescence is a time of increased risk-taking. In recent years, scientists have made significant strides toward understanding this phenomenon. The dual-systems model, which attributes adolescent risk-taking to a temporary imbalance during this period between the brain’s responsiveness to reward and its ability to restrain impulses, has proved to be a useful and generative heuristic for explaining the rise and fall of risk-taking propensity across the transition from adolescence to adulthood. However, insufficient attention has been paid to sex differences in the mechanisms that contribute to adolescent risky behavior, which is surprising given the theoretical and empirical reasons to expect to find sex differences in these mechanisms (e.g., different pubertal timing, different evolutionary pressures, and sex differences in many forms of real-world risk-taking).
The study’s findings support both a dual-systems model of adolescent development and evolutionary theories about sex differences in the propensity to take risks. Patterns of development in sensation-seeking and impulse control are distinct: both males and females evince a rise and fall in sensation-seeking across development compared to a relatively steady increase in levels of impulse control after early adolescence. This finding is consistent with the proposition of the dual-systems model that the incentive processing and cognitive control systems undergo disparate courses of development (Harden and Tucker-Drob 2011). We also find that levels of sensation-seeking are higher, and levels of impulse control lower, among males than females across the full age span examined. Therefore, to the extent that relatively high sensation-seeking and relatively low impulse control contribute to willingness to engage in risky behavior, our results are also aligned with evolutionary theories that posit that male adolescents and young adults are more predisposed to risk-taking than their female peers.
Two key sex differences were observed in the shapes of the sensation-seeking trajectories. First, females, on average, reached peak levels of sensation-seeking earlier than males. The peak in sensation-seeking fell close to age 16–17 for females and age 18–19 for males. The earlier peak in sensation-seeking for females relative to males is consistent with the notion that development of the incentive processing system, which is thought to heighten sensation-seeking, is tied to pubertal maturation (Steinberg 2008). Girls begin puberty earlier than boys by approximately 18 months (e.g., Sun et al. 2002), a lag that is similar in magnitude to that between the female and male peaks in sensation-seeking observed in the present study. If males and females experience similar responses to puberty in terms of increasing levels of sensation-seeking, but girls start puberty earlier than boys, then it makes sense that girls would reach peak levels of sensation-seeking earlier than boys. Unfortunately, because measures of puberty are not available for the boys in the CNLSY sample, it was not possible for us to directly test whether sex differences in pubertal timing explain the earlier peak in sensation-seeking among females. Also, we were unable to test whether the initiation of the rise in sensation-seeking occurs earlier for girls than for boys because this construct was not assessed among young enough individuals in the CNLSY. Between age 10–11, the youngest age at which it was assessed, and age 12–13, the rise in sensation-seeking was already well under way for both sexes.
A second observed sex difference in the trajectories of sensation-seeking was a pronounced difference in the rate of decline in sensation-seeking in early adulthood. After reaching its peak in mid-adolescence, females’ sensation-seeking declined relatively rapidly, whereas males’ sensation-seeking remained fairly high throughout adolescence. Although sensation-seeking fell for both sexes between 18 and 25, the rate of decline was twice as fast for females as for males. Though we did not hypothesize a priori that this pattern would emerge, it is consistent with evolutionary models that posit different functional purposes of sensation-seeking for females versus males. For males, the rise in sensation-seeking in adolescence is thought to serve the evolutionary purpose of increasing the likelihood of attaining high social status and securing access to sexual partners. For many men in the population sampled, social position and steady sexual partners will not be secured until well into adulthood. On the other hand, evolutionary theorists have speculated that, for women, sensation-seeking serves the purpose of facilitating the transition out of the natal environment and into the adult social sphere. This is a task that is likely accomplished at an earlier age than the age at which men establish their social status and gain access to sexual partners. In addition, cultural factors likely reinforce sensation-seeking more for men than for women, which could also contribute to the faster rate of decline in this trait among young adult females.
The timing of growth in impulse control is similar for males and females, consistent with speculation that the development of this capacity is not closely linked to puberty. Females’ rate of growth in impulse control is a bit faster than males’, however, for reasons that are not immediately clear. The slight sex difference in the rate of growth in impulse control could reflect greater societal or evolutionary pressure on females to learn to constrain their impulses. Another unexpected finding was that levels of impulse control appeared to decline slightly between ages 10–11 and 14–15, although the decline was significant only for females. Given that most studies using performance tasks to measure constructs related to impulse control (like inhibitory control) find monotonic increases in impulse control across childhood and adolescence, this result is unexpected [see Luna, Padmanabhan, and O’Hearn (2010) for a review]. We suspect that the apparently greater impulse control evinced by preadolescents is artificial, and reflects an inclination of preadolescents (or very young adolescents) to give socially desirable responses.
For both sensation-seeking and impulse control, the gender gap widened between mid-adolescence (age 14–15) and early adulthood (age 24–25). Because females increased in impulse control at a slightly faster rate than males, females pulled progressively farther away from their male peers in this capacity, at least until the mid-twenties. Similarly, because females reached their peak level of sensation-seeking earlier than males and declined more rapidly thereafter, their separation from males grew incrementally across this age span. Our finding of a widening gender gap with increasing age contrasts with the results of a prior meta-analysis (Cross et al. 2011), which found that sex differences in sensation-seeking and impulsivity were more pronounced in adolescence than adulthood. It should be noted, however, that the conclusions reached in that meta-analysis were based on cross-sectional findings, which may be impacted by cohort effects, whereas our findings are based on longitudinal data.
Importantly, the observed sex differences in the trajectories of sensation-seeking and impulse control, taken together, suggest a more pronounced and more protracted window of vulnerability for males relative to females, which likely serves as a psychological mechanism contributing to males’ greater risk-taking in adolescence and early adulthood. We operationally define the “window of vulnerability” for risk-taking as the period during which sensation-seeking is high relative to impulse control, consistent with the literature linking risk-taking both to high sensation-seeking (e.g., Arnett 1992) and to low impulse control (e.g., Stanford et al. 1996). Defined this way, it is clear that the window is much shorter for females (for whom sensation-seeking is on par with impulse control at ages 12–13 and 14–15, but is never significantly higher than impulse control) than for males, for whom it opens—and opens much wider—across most of the age span examined (between ages 12–13 and 24–25). The pattern observed is consistent with the proposition, generated by models of sexual selection, that males should sustain a readiness to engage in risk-taking until their sexual access to females is established (Daly and Wilson 2001). It is also consistent with epidemiological data showing greater risk-taking among males than females across a wide variety of behaviors.
One question that the present study cannot address is whether the extent and duration of males’ window of vulnerability to risk-taking is influenced by the cultural context. It would be consistent with evolutionary theory if this window were correlated with the timing of transition into adult roles, and more so for males than for females. This hypothesis, which should be tested in future cross-cultural research, is supported by studies finding that both steady work (which confers status) and marriage/cohabitation (which confers access to sex) predict desistance from crime (Jaffee et al. 2013; Laub et al. 1998; Sampson and Laub 1993; Savolainen 2009)—a form of risk-taking. It is also consistent with data suggesting that divorced and widowed men engage in more aggressive risk-taking (male-on-male homicide) than their married peers (Daly and Wilson 2001). In the United States, adult milestones such as full financial independence and marriage occur relatively late in development, often not until the late 20s (Sironi 2013; United Nations 2000). We hypothesize that the window of vulnerability for risk-taking will close earlier for males in societies in which adult roles, especially work and marriage, are adopted earlier.
The present study has several key strengths, including the large, diverse sample and a longitudinal design. Almost all research in this area is cross-sectional, which cannot guarantee that individuals of different ages are drawn from the same population. Still, the present report does have limitations. Two main concerns involve the measures of the key constructs. The sensation-seeking and impulse control scales are indirect measures of the functioning of individuals’ incentive-processing and cognitive control systems. Caution is therefore warranted in interpreting our findings as providing insight into the developmental courses of brain systems. Also, sensation-seeking and impulse control were assessed with three-item scales that may not have captured all the facets of these traits. Fortunately, large samples, such as that employed here, facilitate the detection of signal relative to noise even when using imperfect measures. Same-reporter bias, resulting from relying on self-report of the two key measures, poses a threat to validity as well. This concern is partially mitigated by our use of a latent growth curve model, which relegates this sort of bias to the covariances of the residual terms at a given age, allowing for reliable tests of the other parameters. Still, research using collateral report and/or other methods of assessing the functional state of the cognitive control and incentive processing systems within a longitudinal design would be valuable. However, self-report is a well-established method of assessing personality traits, and the use of the same rater at every time point is a strength.
As expected, the magnitude of sex differences observed in the shapes of the developmental trajectories were modest. However, we do not consider this to be a weakness or limitation of the study. On the contrary, relatively small effects of sex at the trait level may translate into substantial disparities in risk-taking behavior (as well as associated morbidity and mortality) at the population level. Furthermore, our analysis likely underestimated the true differences between males and females due to measurement error, and because females and males may use different bases of comparison when evaluating their own behavioral tendencies.
Conclusion
Our findings suggest that males’ and females’ trajectories of sensation-seeking and impulse control are similar in some respects and consistent with a dual-systems model of development. However, we also find notable sex differences in the relative levels of these traits across development. For example, males’ levels of sensation-seeking are higher and their levels of impulse control lower than females’ across the full age span examined. Also, females exhibit a more pronounced decline in sensation-seeking after mid-adolescence than do males. Among males, there is a greater disparity between sensation-seeking and impulse control than among females, which may help to explain the uniquely high propensity of adolescent and young adult males to engage in risky behavior (Wilson and Daly 1985). Further research is needed to parse the contribution of biological mechanisms, such as changes in pubertal hormones, versus cultural mechanisms, such as the sex-specific reinforcement of sensation-seeking or impulse control to the sex differences in these constructs at various points in development.
Notes
When these series of analyses were run as unconditional models (omitting the control variables) the results still favored the cubic models (results available upon request).
References
Arnett, J. J. (1992). Reckless behavior in adolescence: A developmental perspective. Developmental Review, 12, 339–373.
Asparouhov, T., & Muthen, B. (2006). Comparison of estimation methods for complex survey data analysis. Retrieved from http://www.statmodel.com/download/SurveyComp21.pdf.
Casey, B. J., Getz, S., & Galvan, A. (2008). The adolescent brain. Developmental Review, 28, 62–77. doi:10.1016/j.dr.2007.08.003.
Casey, B. J., Jones, R. M., & Somerville, L. H. (2011). Braking and accelerating of the adolescent brain. Journal of Research on Adolescence, 21, 21–33. doi:10.1111/j.1532-7795.2010.00712.x.
Chen, L. H., Baker, S. P., Braver, E. R., & Li, G. (2000). Carrying passengers as a risk factor for crashes fatal to 16- and 17-year-old drivers. Journal of the American Medical Association, 283(12), 1578–1582.
Cross, C. P., Copping, L. T., & Campbell, A. (2011). Sex differences in impulsivity: A meta-analysis. Psychological Bulletin, 137(1), 97–130. doi:10.1037/a0021591.
Cross, C. P., Cyrenne, D. L. M., & Brown, G. R. (2013). Sex differences in sensation-seeking: A meta-analysis. Scientific Reports, 3, 1–5. doi:10.1038/srep02486.
Dahl, R. (2004). Adolescent brain development: A period of vulnerabilities and opportunities. Annals of the New York Academy of Sciences, 1021, 1–22.
Dahlen, E. R., Martin, R. C., Ragan, K., & Kuhlman, M. M. (2005). Driving anger, sensation seeking, impulsiveness, and boredom proneness in the prediction of unsafe driving. Accident Analysis and Prevention, 37, 341–348. doi:10.1016/j.aap.2004.10.006.
Daly, M., & Wilson, M. (2001). Risk-taking, intrasexual competition, and homicide. Nebraska Symposium on Motivation, 47, 1–36.
Donohew, L., Zimmerman, R., Cupp, P. S., Novak, S., Colon, S., & Abell, R. (2000). Sensation seeking, impulsive decision-making, and risky sex: Implications for risk-taking and design of interventions. Personality and Individual Differences, 28, 1079–1091. doi:10.1016/S0191-8869(99)00158-0.
Ellis, B. (1992). The evolution of sexual attraction: Evaluative mechanisms in women. In J. H. Barkow, L. Cosmides, & J. Tooby (Eds.), The adapted mind (pp. 267–288). New York: Oxford University Press.
Federal Bureau of Investigation. (2010). Uniform crime reports, table 33. http://www.fbi.gov/about-us/cjis/ucr/crime-in-the-u.s/2010/crime-in-the-u.s.-2010/tables/10tbl33.xls.
Finer, L. B. (2010). Unintended pregnancy among U.S. adolescents: Accounting for sexual activity. Journal of Adolescent Health, 47, 312–314.
Forbes, E. E., & Dahl, R. E. (2010). Pubertal development and behavior: Hormonal activation of social and motivational tendencies. Brain and Cognition, 72, 66–72. doi:10.1016/j.bandc.2009.10.007.
Galvan, A., Hare, T., Voss, H., Glover, G., & Casey, B. J. (2007). Risk-taking and the adolescent brain: Who is at risk? Developmental Science, 10, F8–F14.
Gosling, S. D., Rentfrow, P. J., & Swann, W. B., Jr. (2003). A very brief measure of the Big-Five personality domains. Journal of Research in Personality, 37(6), 504–528.
Harden, K. P., Quinn, P. D., & Tucker-Drob, E. M. (2012). Genetically influenced change in sensation-seeking drives the rise of delinquent behavior during adolescence. Developmental Science, 15, 150–163. doi:10.1111/j.1467-7687.2011.01115.x.
Harden, K. P., & Tucker-Drob, E. M. (2011). Individual differences in the development of sensation-seeking and impulsivity during adolescence: Further evidence for a dual systems model. Developmental Psychology, 47, 739–746.
Heron, M. (2012). Deaths: Leading causes for 2009. National Vital Statistics Report, 61, 1–95.
Jaffee, S. R., Lombardi, C. M., & Coley, R. L. (2013). Using complementary methods to test whether marriage limits men’s antisocial behavior. Development and Psychopathology, 25(01), 65–77. doi:10.1017/S0954579412000909.
Laub, J. H., Nagin, D. S., & Sampson, R. J. (1998). Trajectories of change in criminal offending: Good marriages and the desistance process. American Sociological Review, 63, 225–238.
Luna, B., Padmanabhan, A., & O’Hearn, K. (2010). What has fMRI told us about the development of cognitive control through adolescence? Brain and Cognition, 72(1), 101–113. doi:10.1016/j.bandc.2009.08.005.
McArdle, J. J., & Nesselroade, J. R. (2002). Growth curve analysis in contemporary psychological research. In J. Schinka & W. Velicer (Eds.), Comprehensive handbook of psychology (Vol. 2, pp. 447–480)., Research methods in psychology New York, NY: Wiley.
Moffitt, T. E., Caspi, A., Rutter, M., & Silva, P. A. (2001). Sex differences in antisocial behaviour: Conduct disorder, delinquency, and violence in the Dunedin Longitudinal Study. Cambridge, England: Cambridge University Press.
Moffitt, T., Poulton, R., & Caspi, A. (2013). Lifelong Impact of early self-control childhood: Self-discipline predicts adult quality of life. American Scientist, 101, 352–359. doi:10.1511/2013.104.352.
Montoring the Future. (2009). Trends in lifetime prevalence of use of various drugs. University of Michigan. Retrieved from http://monitoringthefuture.org/data/09data/pr09t1.pdf.
Muthén, L. K., & Muthén, B. O. (2009). Mplus user’s guide (v5.2). Los Angeles: Muthén & Muthén.
National Highway Traffic Safety Administration. (2007). Comparison analysis of fatality trend by age group—1996 to 2005. Retrieved from http://www-nrd.nhtsa.dot.gov/Pubs/810759.pdf.
Op de Macks, Z. A., Moor, B. G., Overgaauw, S., Güroğlu, B., Dahl, R. E., & Crone, E. A. (2011). Testosterone levels correspond with increased ventral striatum activation in response to monetary rewards in adolescents. Developmental Cognitive Neuroscience, 1(4), 506–516. doi:10.1016/j.dcn.2011.06.003.
Patton, J. H., & Stanford, M. S. (1995). Factor structure of the Barratt impulsiveness scale. Journal of Clinical Psychology, 51, 768–774.
Pfeifer, J. H., & Allen, N. B. (2012). Arrested development? Reconsidering dual-systems models of brain function in adolescence and disorders. Trends in Cognitive Science, 16, 322–329. doi:10.1016/j.tics.2012.04.011.
Piquero, A. R. (2008). Taking stock of developmental trajectories of criminal activity over the life course. The long view of crime: A synthesis of longitudinal research, 23–78. doi:10.1007/978-0-387-71165-2_2.
Quinn, P. D., & Harden, K. P. (2013). Differential changes in impulsivity and sensation seeking and the escalation of substance use from adolescence to early adulthood. Development and Psychopathology, 25, 223–239.
Raykov, T. (2005). Analysis of longitudinal studies with missing data using covariance structure modeling with full-information maximum likelihood. Structural Equation Modeling, 12(3), 493–505. doi:10.1207/s15328007sem1203_8.
Romer, D., Duckworth, A. L., Sznitman, S., & Park, S. (2010). Can adolescents learn self-control? Delay of gratification in the development of control over risk taking. Prevention Science, 11, 319–330.
Sampson, R. J., & Laub, J. H. (1993). Crime in the making: Pathways and turning points through life. Cambridge, MA: Harvard University Press.
Savolainen, J. (2009). Work, family, and criminal desistance: Adult social bonds in a Nordic welfare state. British Journal of Criminology, 49(3), 285–304.
Schmitt, D. P., Realo, A., Voracek, M., & Allik, J. (2008). Why can’t a man be more like a woman? Sex differences in Big Five personality traits across 55 cultures. Journal of Personality and Social Psychology, 94, 168–182. doi:10.1037/0022-3514.94.1.168.
Shirtcliff, E. A., Dahl, R. E., & Pollak, S. D. (2009). Pubertal development: Correspondence between hormonal and physical development. Child Development, 80(2), 327–337. doi:10.1111/j.1467-8624.2009.01263.x.
Shulman, E. P., Steinberg, L. D., & Piquero, A. R. (2013). The age-crime curve in adolescence and early adulthood is not due to age differences in economic status. Journal of Youth and Adolescence, 42(6), 848–860. doi:10.1007/s10964-013-9950-4.
Sironi, M. (2013). The transition to adulthood in the developed Western world: A focus on the achievement of economic independence and on the role of family background. Dissertation retrieved from: http://repository.upenn.edu/dissertations/AAI3565237.
Smith, A. R., Chein, J., & Steinberg, L. (2013). Impact of socio-emotional context, brain development, and pubertal maturation on adolescent risk-taking. Hormones and Behavior, 64(2), 323–332.
Spear, L. P. (2000). The adolescent brain and age-related behavioral manifestations. Neuroscience and Biobehavioral Reviews, 24, 417–463.
Spear, L. P. (2013). Adolescent neurodevelopment. Journal of Adolescent Health, 52(2), S7–S13.
Stanford, M. S., Greve, K. W., Boudreaux, J. K., Mathias, C. W., & Brumbelow, J. L. (1996). Impulsiveness and risk-taking behavior: Comparison of high-school and college students using the Barratt Impulsiveness Scale. Personality and Individual Differences, 21(6), 1073–1075.
Steinberg, L. (2008). A social neuroscience perspective on adolescent risk-taking. Developmental Review, 28(1), 78–106.
Steinberg, L. (2010). A dual-systems model of adolescent risk-taking. Developmental Psychobiology, 52, 216–224. doi:10.1002/dev.20445.
Steinberg, L., Albert, D., Cauffman, E., Banich, M., Graham, S., & Woolard, J. (2008). Age differences in sensation-seeking and impulsivity as indexed by behavior and self-report: Evidence for a dual systems model. Developmental Psychology, 44, 1764–1778.
Steinberg, L., Cauffman, E., Woolard, J., Graham, S., & Banich, M. (2009). Are adolescents less mature than adults? Minors’ access to abortion, the juvenile death penalty, and the alleged APA “flip-flop”. American Psychologist, 64(7), 583–594.
Strang, N. M., Chein, J. M., & Steinberg, L. (2013). The value of the dual systems model of adolescent risk-taking. Frontiers in Human Neuroscience, 7(233), 1–4. doi:10.3389/fnhum.2013.00223.
Sun, S. S., Schubert, C. M., Chumlea, W. C., Roche, A. F., Kulin, H. E., Lee, P. A., et al. (2002). National estimates of the timing of sexual maturation and racial differences among US children. Pediatrics, 110, 911–919.
United Nations Department of Economic and Social Affairs of the United Nations Secretariat. (2000). World marriage patterns. http://www.un.org/en/development/desa/population/publications/pdf/family/marriage-patterns.pdf.
Urošević, S., Collins, P., Muetzel, R., Lim, K. O., & Luciana, M. (2014). Pubertal status associations with reward and threat sensitivities and subcortical brain volumes during adolescence. Brain and Cognition,. doi:10.1016/j.bandc.2014.01.007.
Vazsonyi, A. T., & Belliston, L. M. (2007). The family → low self-control → deviance: A cross-cultural and cross-national test of self-control theory. Criminal Justice and Behavior, 34(4), 505–530. doi:10.1177/0093854806292299.
Wilson, M., & Daly, M. (1985). Competitiveness, risk taking, and violence: The young male syndrome. Ethology and Sociobiology, 6, 59–73.
Wilson, M., & Daly, M. (1993). Lethal confrontational violence among young men. In N. Bell & R. Bell (Eds.), Adolescent risk-taking (pp. 84–106). Newberry Park, CA: Sage.
Zuckerman, M., & Kuhlman, D. M. (2000). Personality and risk-taking: Common biosocial factors. Journal of Personality, 68, 999–1029. doi:10.1111/1467-6494.00124.
Author Contributions
EPS conducted the analyses and took the lead in authoring the article. All four authors helped to generate the research idea and to revise and edit the article. All authors read and approved of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Shulman, E.P., Harden, K.P., Chein, J.M. et al. Sex Differences in the Developmental Trajectories of Impulse Control and Sensation-Seeking from Early Adolescence to Early Adulthood. J Youth Adolescence 44, 1–17 (2015). https://doi.org/10.1007/s10964-014-0116-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10964-014-0116-9