
Abstract
The last interglacial was the most recent time when temperatures were 1–2 °C above modern, but little is known of this period in the lowland Neotropics. Equally, data for the full glacial period are very limited. A detailed analysis of the period between ~ 137 and 100 ka was completed to provide a paleoecological history of the last interglacial in central Panama. Two additional fossil diatom records from the same depositional basin provided records of the glacial period. Diatom assemblages were compared across all cores and the dominant species of both glacial and interglacial periods was almost always Aulacoseira granulata. Other species, e.g. Aulacoseira agassizii, Achnanthidium minutissimum, Nitzschia amphibia, and Navicula radiosa, had distinctive patterns of abundance within the record, indicating a shallowing of the lake in the last millennia of MIS5e. The period between 119 and 108 ka witnessed the most change within the high-resolution portion of the record suggesting an increased lake level, with expanding lake margins. Other sediment cores collected within the caldera revealed the continued presence of a lake at El Valle through much of the glacial period, with shallowing evident at the time of the LGM. The changes in diatom assemblages at El Valle provide one of the oldest precipitation records from Central America tracking mean ITCZ position between the last interglacial and glacial periods. Furthermore, the evidence for both a wet interglacial and glacial period support palynological findings that the humid environments of Central Panama were not interrupted by glacial aridity.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Exceptional tropical biodiversity has previously been attributed to climatic stability in the museum hypothesis (Stebbins 1974; Wiens and Donoghue 2004), and climatic instability in the refugial hypothesis (Haffer 1969; Pennington et al. 2004). At the heart of such studies lie assumptions about interglacial-glacial climate oscillations in the tropics. Refugialists have suggested that glacial periods were times of strong climate change with long-term, continental-scale aridity determining plant communities (Haffer and Prance 2001; Van der Hammen and Absy 1994). However, others have argued that while precipitation varied throughout the Pleistocene, profound aridity sufficient to cause the forest to be replaced by savanna over large areas is a myth (Bush et al. 2011; Colinvaux et al. 2001; Wang et al. 2017). Additionally, empirical data and general circulation model simulations, such as CCSM4, point to the last interglacial as being drier than the last glacial maximum (Bush et al. 2010; D’Agostino et al. 2002; Fritz et al. 2007; Hanselman et al. 2011). As assumptions about past climates give way to empirical data, evidence points to fine-grained temporal and spatial patterns.
Consequently, glacial and interglacial cycles are no longer viewed as climatically monolithic events. Precessional changes in Earth’s orbit around the Sun have been identified as important drivers of tropical precipitation change (Hooghiemstra 2002; Cruz et al. 2007) resulting in approximately 21,000-year wet-dry cycles with the peak of each phase lasting about 3000–6000 years (Baker and Fritz 2015; Bush et al. 2002; Sublette Mosblech et al. 2012). In Central America, wet and dry periods are associated with the position of the Inter-tropical Convergence Zone (ITCZ) and, in turn are linked to Atlantic sea-surface temperatures (SST; Peterson and Haug 2006). Superimposed on precessional rhythms are faster-paced phenomena, e.g. Dansgaard–Oeschger cycles and Heinrich events. In this discussion, glacials have received far more attention than interglacials, largely because of the quality of available records and the greater apparent climatic instability of glacial periods over interglacials. Temperature change, initially downplayed in the tropics, was clearly part of the fabric of that ice-age climate change. Glacial cooling in the tropics of ~ 7–9 °C in the mountains and 4–7 °C in the lowlands has been documented from a wide variety of sources (Colinvaux and De Oliveira 2000; Correa-Metrio et al. 2012; Hooghiemstra and van der Hammen 1998; Ledru et al. 2005; Van der Hammen 1974). Little is still known of interglacial temperatures, though those of MIS5e are thought to have been 1–2 °C warmer than present (Hooghiemstra 1984).
Records reflecting both precession and millennial-scale oceanic changes include the titanium concentrations in the sediments from the Cariaco Basin, and the multiproxy records from Lake Petén Itzá in the Yucatán Peninsula. The river discharge into the Cariaco Basin reflected the position of the ITCZ with a southern (northern) position corresponding to cool (warm) SSTs and low (high) rainfall over Venezuela (Peterson and Haug 2006). The same migration of the ITCZ was inferred to bring similar rainfall patterns to the Yucatán (Haug et al. 2003). The consequence of this pattern was that the early Holocene produced the driest conditions of the last interglacial-glacial transition in Venezuela (Peterson and Haug 2006). However, in the Yucatán, Hodell et al. (2008) suggested that on millennial scales it is the early initiation of dry season conditions forced by precession, and a lagging migration of the ITCZ, that resulted in the driest conditions occurring not in the Holocene but in the deglacial period between approximately 17 and 13 ka BP.
Testing the generality of these observations requires a setting between Venezuela and Guatemala. Very few ancient lakes exist in the lowlands of Central America, but Lake La Yeguada and Paleolake El Valle in Panama offer some insights. La Yeguada filled approximately 14.3 ka, suggesting an onset of wetter conditions at that time (Bush et al. 1992). Between 12 and 8 ka La Yeguada had a highstand above modern levels, and from 8 to 6 ka, the warmest time of the Holocene, the lake became shallow. The El Valle crater supported a lake until c. 8 ka, and thereafter most of the basin was not accumulating organic sediment (Bush and Colinvaux 1990).
The warmest portion of the last interglacial period (MIS5e; approximately 137–120 ka; Tzedakis 2003) is recognized as a period perhaps about 2 °C warmer than present (CAPE 2006; Hanselman et al. 2005). In the southern tropical Andes, MIS5e was dry as Lake Titicaca shrank in size by 85% compared with modern (Seltzer et al. 1998). Conversely, in the Colombian Andes, records from the High Plains of Bogota indicated that the interglacial was warm and wet (Groot et al. 2011; Hooghiemstra 1984). Northern South America as a whole followed this trend (Nikolova et al. 2013) as sediments from the Cariaco Basin also indicated wet interglacial conditions attributed to a more northern position of the ITCZ (Riboulleau et al. 2014; Peterson and Haug 2006).
Only one lowland rain forest site in lower Central America is known to contain sediments that date to MIS5e, the crater of El Valle de Antόn in Panama. The first sediment core raised from that site in 1988 was analyzed for fossil pollen by Bush and Colinvaux (1990). They obtained a 57-m-long core that was beyond the range of 14C bulk dating (> 35,000 years) at 10 m in depth. Thus, the majority of the core was of Pleistocene age. The presence of pollen of montane forest taxa such as Quercus and Morella (ex. Myrica), suggested that the glacial-aged deposits represented climates as much as 5 °C cooler than modern. Bush and Colinvaux (1990) observed that near the base of that core was a section of apparently warmer climatic conditions that they suggested may have represented MIS5e.
Here we provide the first analysis of lake level changes based on fossil diatoms from Central Panama that captures the last interglacial. We seek to answer the basic question: was the last interglacial dry or wet over Central Panama, and how did that compare with the glacial period?
Study site
The El Valle caldera (Fig. 1) is located in the lowlands of central Panama (8°36′08″N/80°7′33″W) on the easternmost edge of the Central American Volcanic Complex (Coates and Obando 1996). The floor of the caldera lies 500 m above sea-level (a.s.l.) with the rim of its horseshoe-shaped bowl stretching to 1185 m a.s.l. and a total basin area of approximately 22 km2. The flat floor of El Valle supported a Pleistocene-aged paleolake that has shrunk to a small marsh (Bush and Colinvaux 1990; Hidalgo et al. 2011). Richard Wharry, an experienced water well driller at this site, told us that beneath the floor of the caldera are several aquifers, including at least one that is warm (R. Wharry pers. commun.). Springs feed the Rio Antón, which rises within the basin and flows through an outlet in the southwestern wall of the crater.

Study site of El Valle, Panama and comparison sites. a Coring locations of EV88, EV90, and EV08 within the El Valle Caldera. b Inset map of Panama showing location of El Valle (1) and La Yeguada (2). c Locations mentioned in the text as follows: Cariaco Basin, Venezuela (3), Petén Itzá, Guatemala (4), Bogotá, Colombia (Fúquene) (5), and Lake Titicaca, Peru/Bolivia (6). The dashed line depicts the mean modern position of the ITCZ
The climate of the El Valle region is humid tropical with temperatures ranging from 18 to 37 °C with an average annual temperature of 27 °C. Mean annual precipitation at the Santiago meteorological station, 108 km away, is 3000 mm with seasonal rain falling mostly between July and January (ETESA 2009). The natural vegetation of the caldera would have been closed canopy lowland tropical forest with grassy marshes around the lake margin (Bush and Colinvaux 1990; Cárdenes-Sandí 2015).
Methods
Three sediment cores from the floor of El Valle de Antón, central Panama were analyzed to reconstruct precipitation patterns during the last interglacial and glacial periods in Central America. The focus of this analysis is a 54-m-long sediment core that was raised in 2008 (EV08) from the center of the basin using a truck-mounted water-well drilling rig. The core was collected in 3-m-long sections of schedule 40 PVC which was cut into 1-m sections and sealed in the field. The other cores, collected in 1988 (EV88) and 1990 (EV90), were collected in a similar manner to EV08. EV90 was drilled approximately 50 m further out from the basin edge than EV88 (Fig. 1).
The lowermost 9 m of EV08 were sampled for 234U/230Th dating at 8 locations. A total of 145 diatom subsamples, collected at 5-cm intervals, were prepared by heating samples in HCl and H2O2, then mounted in Naphrax® (Battarbee 1986). Fossil diatom counts to 300 valves were performed at 1000× magnification on a Zeiss Axioskop photomicroscope. Identifications were based on standard texts (Patrick and Reimer 1966, 1975; Round et al. 1990) and the online resource: Diatoms of the United States (Spaulding et al. 2010). Identifications were checked for the most updated taxonomy using Algaebase (Guiry and Guiry 2015), and ANSP/NAWQA/EPA taxonomy list and algae image database (ANSP 2012). Loss-on-ignition was performed at the same depths sampled for diatoms in EV08 following the methods of Heiri et al. (2001).
Cores EV88 and EV90 were sampled at a lower resolution than EV08 with 11 samples taken in EV88 above core depth 14 m, and 25 samples taken in EV90 between 14 and 53 m core depth. Each sample taken from EV88 and EV90 was counted to at least 500 valves.
Detrended correspondence analysis (DCA; Hill 1979) identified fluctuations in diatom assemblages in the EV08 core. Diatom zones were isolated with cluster analysis CONISS (Grimm 1987) in RStudio v. 0.98.1103 using the “rioja” package. Analysis of similarities (ANOSIM; Clarke 1993) was used to determine if the zone of highest DCA score fluctuations identified in the CONISS tree was statistically different from the rest of the dataset.
Results
Core stratigraphy
Four main types of sediment made up the cores from El Valle: dark gyttja, sandy clay, laminated clay, and organic-rich clay. All cores contained laminated clay sections between 30 and 35 m that overlay organic-rich clay between 43 and 47 m and sandy clays at the base of all cores. Thus the portion of EV08 reported here was comprised of sandy clays. Loss-on-ignition (LOI) values remained low and variable through the entire EV08 core with no clear trends corresponding to peaks in the commonest diatom species. Gaps in each core are sections where no core was recovered in the coring process, rather than gaps in sedimentation. Diatoms were well preserved and no signs of biogenic silica dissolution were evident.
Statistical analysis
Cluster analysis within the El Valle cores revealed three diatom zones from EV08 and two diatom zones from EV88. No zones were isolated from the EV90 core. The axes scores from the DCA of EV08 fossil diatom data indicated broad similarities between sample scores as all lay within a range of 2 standard deviations of species turnover. The ANOSIM results, however, did reveal that the sediments in EV08 between 49.5 and 51 m depth were significantly different (p = 0.001, R = 0.76) from samples above and below. Low abundances of benthic species, the dominance of centric planktonics, and an increased presence of epiphytic species characterized this significantly different period.
Diatom zones: EV08 core
Zone EV08-1: 54–51 m
Zone EV08-1 is composed of the bottom 3 m of the EV08 core. Aulacoseira granulata (Ehrenberg) Simonsen (ESM1) was the dominant species in assemblages throughout the zone (Fig. 2), while Aulacoseira agassizii (Ostenfeld) Simonsen (< 20%), Ulnaria acus (Kützing) Aboal (< 10%), Nitzschia columbiana Sovereign (< 10%) and Nitzschia amphibia Grunow (10%) had low peaks within this zone (Fig. 2). This period ended with the highest abundance of benthic diatoms, e.g. Navicula radiosa Kützing (25%), of the record. Percentages of tychoplanktonic araphid species, both epiphytic and episammic, remained low in comparison with planktonic and benthic species. LOI in this zone shared no peaks with species in the diatom assemblage, but remained consistently higher toward the end of the zone. DCA scores remained low in this period.

Diatoms present from 54 to 45 m depth in core EV08 at El Valle, Panama. Grey, dashed lines indicate zone separations and white areas indicate areas of sediment lost during the coring process. Species shown are those > 1% abundance. Black lines in plots indicate ×5 exaggeration line
Zone EV08-2: 51–49.5 m
Zone EV08-2 was the region between 51 and 49.5 m core depth. This was the only section in the EV08 record where A. agassizii increased above 20% abundance. Navicula radiosa (< 10%) was present in low abundance during the first half of this period until a transition from benthic to mainly planktonic species occurred, including A. agassizii, and epiphytic tychoplanktonic species, e.g. Ulnaria acus. Nitzschia columbiana was replaced by N. amphibia, and Achnanthidium minutissimum (Kützing) Czarnecki was completely absent in this zone. LOI remained variable, with a peak before benthic species decreased below 5%. The 1.5 m of sediment represented by this zone experienced the largest variation in DCA scores as well as the lowest abundances of benthic diatoms found in the EV08 record.
Zone EV08-3: 49.5–45.5 m
The top 4 m of EV08 comprised Zone EV08-3. Aulacoseira granulata was the most dominant species of the assemblage, occurring at > 90% in most samples. Benthic diatoms including N. radiosa and A. minutissimum, reappeared in low abundances in this section. Epiphytic Ulnaria species decreased in abundance, but overall, all tychoplanktonic species remained constant in lower abundances than EV08-2. LOI was variable throughout this zone, fluctuating at levels below 11%. DCA scores returned to levels of low fluctuation during this zone.
Diatom assemblages: EV90 core
CONISS clustering resulted in only one diatom zone for the EV90 core. The sediments within the lowest portion of the core were mainly sandy clay, contained low abundances of planktonic diatoms, and had the highest abundances of benthic species in the core (Fig. 3). Unusually for this site, a combination of epiphytic species, e.g. Ulnaria delicatissima (W. Smith) Aboal & Silva and Ulnaria ulna (Nitzsch) Compére, and benthic species e.g. Nitzschia palea (Kützing) Smith and Rhopalodia musculus (Kützing) Müller were more abundant than A. granulata in the basal meters.

Core EV88 from 32 to 14.5 ka (Top Panel). Age is expressed in thousands of years before present (ka). Dashed grey line indicates zone separation. Solid grey line serves to differentiate between EV88 and EV90 records. Core EV90 from 53 to 14 m depth (Bottom Panel). No ages could be provided from this record, but comparisons among core stratigraphies indicate a mostly glacial age as labelled. Note: The two records should not be interpreted as continuous and are only provided together here for a comparison of glacial conditions. Some overlap may occur in age between the bottom of EV88 and the top of EV90. White bars indicate material lost in coring process. Species shown in each panel are those > 5% abundance
Around 50 m depth, assemblages of EV90 were increasingly dominated by A. granulata, generally accounting for 50–95% of the diatoms present, with either gyttja or laminated clay sediments. Ulnaria delicatissima (10–20%) and Ulnaria ulna (20–40%) were also present, increasing in abundances from 50 to 40 m in depth. The remaining assemblages from EV90 contained fairly steady abundances of planktonic diatoms including A. granulata (40–90%), Cyclotella meneghiniana Kützing (< 40%), and Discostella stelligera (Cleve & Grunow) Houk & Klee (30–40%), which fluctuated in abundance with one another. Additional benthic species found elsewhere in the record, such as Achnanthes Bory and Epithemia Kützing species, were almost completely absent throughout the rest of the record and occurred only in low abundances for short time periods.
Diatom zones: EV88 core
Zone EV88-1: 10–6.8 m
Zone EV88-1 spanned the last 3.2 m of the EV88 core. Aulacoseira granulata fluctuated in abundance while C. meneghiniana remained steady at about 75%. Ulnaria delicatissima increased abruptly to about 30% at the end of the zone when C. meneghiniana began to decrease. The sediment in this zone was composed of mostly gyttja with a transition to organic-rich clay in the final 20 cm.
Zone EV88-2: 6.8–4.7 m
The remaining 2.1 m of EV88 were characterized by the lowest abundances of planktonic diatoms throughout the core. Aulacoseira granulata and C. meneghiniana both declined to levels of < 20%. Benthic species, e.g. N. amphibia (10–20%), N. radiosa (10–30%), Gomphonema spp. Ehrenberg (10–20%), and Achnanthes species (< 10%) dominated the top of the core. The sediment of this zone was composed completely of organic-rich clay.
Establishing a chronology
Prior analyses of the fossil pollen record from EV88, similar in length and gross morphology to EV08, led to the suggestion that the depth between 45 and 54 m represented MIS5e (Bush and Colinvaux 1990). Subsequent 234U/230Th dating of the EV08 record (ESM2) supported this estimate and provided evidence that the onset of deposition at El Valle took place shortly before the onset of MIS5.
Core EV08
The eight samples submitted for 234U/230Th dating from EV08 proved to have too much detrital Th for reliable age estimates. Only one sample from 45.98 m provided an age of 134.6 ± 8.2 ka (ESM2) that was credible.
To refine the chronology, the diatom and pollen data from EV08 were aligned to the composite δ18O speleothem data from Sanbao Cave, China (Cheng et al. 2016). This precipitation record from Sanbao Cave is comprehensive, covering the past 640 ka, and completely dated by 234U/230Th methods. Cheng et al. (2016) attribute precession as the driving force of changes in the Asian monsoon over the course of this record. Similarly, precipitation changes in Central and northern South America have been attributed to changes in precessionally forced Atlantic SST (Hodell et al. 2008; Peterson and Haug 2006; Yarincik et al. 2000). Therefore, we aligned wet-dry events, expected to be antiphased due the geographical variation between the Western and Eastern hemispheres (Maher and Hu 2006), between the Sanbao Cave and El Valle diatom records.
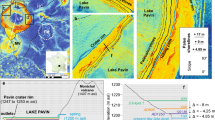
Wet-dry events in the El Valle record were determined using A. granulata abundances. Although a planktonic diatom, A. granulata was found to thrive in relatively well-mixed, shallow, wide, water bodies (Kilham et al. 1986). As planktonic diatoms predominated throughout the record, the observed oscillations in A. granulata reflected shallowing, but not enough to eliminate planktonic diatoms. Increases in A. granulata were interpreted to indicate a shallower and drier system. Wet and dry periods in EV08 samples were aligned (antiphased) to oscillations in the Sanbao Cave record as close to the 134.6 ± 8.2 ka date as possible. Five main tie points were established between the Sanbao and EV08 records, with remaining depths assigned ages through linear interpolation between tie points (Fig. 4). The resulting age range spanned from 100 to 137 ka for sediments between 45.5 and 54 m core depth (Fig. 5).

Core EV08 dating with the A. granulata curve against a longer climate sequence. a The Aulacoseira granulata abundance curve from EV08. No complete drying of the lake occurred within the record and increased percentages indicate drier conditions. The grey bar indicates the period of Quercus absence in the EV08 record by depth. b The dated δ18O composite speleothem record from Sanbao Cave, China (Cheng et al. 2016) used for age alignment. More depleted values (up) indicate wetter conditions in China, and drier conditions in Central America. The 5 major tie points used in dating between El Valle and Sanbao Cave are numbered in panels a and b. c Deviation from modern temperature throughout the longer Sanbao sequence (Kawamura et al. 2007). The cooling necessary for Quercus presence is indicated by the dashed line. The grey bar indicates Quercus absence in the EV08 record after dating

The chronology for El Valle, Panama. a The left panel shows the Markov Chain Monte Carlo iterations for the Bayesian Bacon model used for EV88. The center panel depicts the prior (dark grey line) and posterior (curve) for the accumulation rate. The right panel displays the memory for the age model. b Age-depth plot for core EV88 from El Valle Caldera, Panama. Ages are expressed as calibrated 14C years before present (BP). Calibrations were performed using IntCal13 within the R Bacon package v.2.2 (Blaauw and Christen 2011). Grey lines indicate 95% confidence intervals; the black line indicates the best model based on the weighted mean age for each depth. c Age-depth plot for core EV08 (ages are expressed as kilo-years before present (ka)). Solid line represents the linear interpolation between tie points (vertical lines) between EV08 and Cheng et al. (2016)
The aligned El Valle-Sanbao chronology required further support as the derived age for 45.98 m was approximately 102 ka, considerably younger than the 134.6 ± 8.2 ka derived from empirical dating. The Quercus pollen curve from EV08 (Cárdenes-Sandí 2015) served as a temperature proxy for glacial/interglacial transitions (Fig. 4). A cooling of 2.5–6 °C from modern temperatures would be necessary to bring Quercus to the upper peaks of the El Valle caldera, as suggested by Bush and Colinvaux (1990). Quercus was consistently present in the samples from from EV08 with the exception of 51.5 to 53 m depth. Only this portion of the core was as warm as modern, and therefore is very likely to align with MIS5e (c. 125–134 ka). We conclude from this observation that at El Valle the 234U/230Th dates give a very general impression of the period of sedimentation, but the large errors on these dates provide a strong warning that they should not be relied on to provide accurate age assessments.
The resulting chronology placed zone EV08-1 at the initiation of the last interglacial period from around 137–119 ka (Fig. 6). Zone EV08-2 was defined as the MIS5e/MIS5d transition from 119 to 108 ka. The final zone, EV08-3, was defined as within MIS5c from 108 to 100 ka.

Diatoms present from 137 to 100 ka in core EV08 at El Valle, Panama. Grey, dashed lines indicate zone separations and white areas indicate areas of sediment lost during the coring process. Species shown are those > 1% abundance. Insolation is plotted from 8°N
Core EV90
Material was not available for dating of EV90, but due to its proximity to EV88, the depths analyzed between 14 and 38 m were defined as glacial aged. Core stratigraphy suggests a late Pleistocene age, originating in the last interglacial, and ending in the last glacial period. The lowermost 15 m from 38 to 53 m core depth span at least a portion of MIS5e; however, the lack of a firm chronology limits the scope of analysis of this core.
Core EV88
A chronology for EV88 was established using five bulk sediment 14C dates shown in Table 1, previously reported by Bush and Colinvaux (1990). An age model for EV88 was created using Bacon v. 2.2 age modeling (Blaauw and Christen 2011) and the IntCal13 calibration curve (Reimer et al. 2013).
In the EV88 core, the deepest meaningful radiocarbon dates end in the deglacial period (c. 14.5 ka), but in the tenth meter reach infinity. Sedimentation remained fairly constant through this record (Fig. 5).
Discussion
Lake El Valle during MIS5
Detailed analysis of MIS5-aged sediments suggests the existence of a nutrient-rich water body from 137 to 100 ka at El Valle. The most striking changes in this record revolve around the abundance of A. granulata, a species common to shallow, turbid, nutrient-rich environments (Haberyan et al. 1997; Kilham et al. 1986). Aulacoseira granulata is also commonly found in rivers, streams, and lakes during periods of turnover (Pérez et al. 1999; Kilham et al. 1986). Additionally, the A. granulata profile of EV08 oscillates regularly with the A. agassizii profile. Aulacoseira agassizii is a large, centric diatom previously reported in deeper water of heavily mixed African lakes with large surface areas (Kilham et al. 1986; Tremarin et al. 2012). Thus A. agassizii is associated in this record with slight increases in lake depth resulting in basin expansion while A. granulata decreases. Aulacoseira agassizii and C. meneghiniana were present in low abundances throughout the record, supporting an inference of a well-mixed system subject to frequent overturn.
Along with A. granulata, the older portions of the core were relatively rich in A. minutissimum, a benthic species that thrives in streams and moving water suggesting likely stream input to the lake. All diatoms present in EV08-1 are tolerant of low alkalinity and occur in nutrient-rich habitats.
A shift in the ecology of the system is evident near the end of EV08-1 at c. 120 ka where the largest peak in benthic diatoms occurs. This peak, composed mainly of N. radiosa, coincides with decreased levels of A. minutissimum. Navicula radiosa thrives in brightly lit waters with low turbidity (McIntire 1968) suggesting a decrease in water level and turbulence, possibly from reduced stream input as A. minutissimum decreases in abundance after c. 128 ka.
Between about 119 and 108 ka (Zone EV08-2), benthic diatoms decreased to approximately 10% until c. 112 ka when they stabilized at < 5% in abundance. At c. 118 ka, A. granulata fell from about 90% representation in the diatom assemblage to oscillate between 35 and 80%. Navicula radiosa declined to < 10% by 118 ka and close to 0% at 112 ka until the end of EV08-2 at 108 ka. The progression toward a wider and slightly deeper water body is marked by peaks of A. agassizii and U. acus (Fritz et al. 1999; Owen et al. 2009). For A. agassizii to reach 20% of the diatom sum suggests a more strongly mixed system. Ulnaria acus is an epiphytic species that increased in abundance with A. agassizii in the last 2 ka of EV08-2 (Fig. 6). The presence of these species along with the decrease in benthic species and increase in aquatic pollen taxa, are associated with higher lake levels resulting in basin expansion (Van der Hammen and Hooghiemstra 2003). Small increases in C. meneghiniana and N. amphibia in EV08-2 may indicate increased organic input to the lake from runoff as both diatoms are characteristic of high concentrations of dissolved organic material (Bradbury et al. 1981). Disappearance of A. minutissimum also suggests a decrease in stream input into the system during this time possibly as a result of lake expansion causing the entry point of the stream to be further removed from the core site. The low species turnover maintained throughout this record, as opposed to more prominent variations between benthic and planktonic species, suggests that lake chemistry may not have experienced marked changes within the 37 ka period analyzed at El Valle.
After 108 ka in EV08-3, diatom abundances return to similar conditions witnessed in the first 5 ka of EV08-1. Consequently, we infer that precipitation at El Valle increased at the onset of glacial conditions (MIS5d). Although the driest time of the EV08 record occurred during MIS5e, no pronounced or prolonged increases in benthic or saline diatoms were sustained within the 37 ka period supporting the continuous existence of a lake at El Valle.
Was the last interglacial wet or dry in Central Panama?
Precipitation in Central and South America is strongly influenced by the position of the ITCZ, linked to Atlantic SST (Peterson and Haug 2006). No other records from Central America exist that reach back to MIS5e; however, the existing records from the Northern Hemispheric Atlantic and South America help reconstruct ITCZ movement between the last interglacial and glacial periods.
Two of the El Valle cores overlapped the last interglacial (MIS5e). First, EV90 showed a low-resolution record rich in benthic species during the inferred last interglacial (possibly 48–52 m). This assemblage represented the shallowest portion of the EV90 core. Second, the EV08 core (located about 0.5 km to the east of EV90) presents an equivalent period that was dominated by Aulacoseira, Navicula, and Achnanthidium. We infer that EV08 represented a body of standing water that was somewhat deeper than that of EV90, and accounts for the presence of the deeper water species A. agassizii in EV08 and not in EV90. The EV08 record also offers far greater detail of the last interglacial than EV90, and further discussion is based on this high-resolution record.
In the Northern Hemisphere, MIS5e has been cited as being warm and wet. The transition to a wetter system after MIS5e in El Valle occurs around the same time as drier conditions emerge on the western coast of Africa in the Gulf of Guinea (Weldeab et al. 2007), Soreq Cave, Israel (Bar-Matthews et al. 2003), and the Cariaco Basin (Yarincik et al. 2000; Fig. 7). Precipitation changes in these records were attributed to precessional forcing of SST which in turn affects the average position of the ITCZ. Therefore, the shift to higher precipitation at the onset of MIS5d suggests an intensification of the influence of the ITCZ over Panama (a more southward average position) as a result of decreasing glacial SSTs.

Regional records featuring hydrological indicators relative to precession at 8°N latitude. a Lake Fúquene, Colombia Arboreal pollen percentage (AP). Increasing AP% indicates warmer temperatures. b El Valle A. agassizii/A. granulata ratio (black). Increasing ratio indicates wetter periods. Planktonic/benthic diatom ratio (gray, dotted). Higher levels indicate drier periods. c Mg/Ca ratio from the Gulf of Guinea (out of phase with El Valle). Higher levels indicate higher riverine input from precipitation to the gulf (Weldeab et al. 2007). d Soreq Cave, Israel δ18O speleothem record (out of phase with El Valle) with more depleted values depicting increased rainfall (Bar-Matthews et al. 2003). e Low resolution Cariaco Ti record reflecting riverine runoff into the basin. Higher Ti levels indicate higher rainfall over Venezuela (Yarincik et al. 2000). f Cariaco Mo data reflecting SST in the Atlantic during MIS5. Increasing levels of Mo indicate more stratification in the water column, and higher SST (Gibson and Peterson 2014). g Seasonality and mean insolation at 8°N. The black line indicates mean summer insolation and the grey line depicts mean winter insolation (Laskar et al. 2004; Paillard et al. 1996)
MIS5e in Colombia (5°N) shows a rapid warming between MIS 6 and 5e with the establishment of humid montane forest at 2540 m elevation about 130 ka (Groot et al. 2011). This peak was relatively short-lived and there was an oscillation toward slightly less arboreal pollen between approximately 125 and 119 ka where the Fúquene record reveals a drop in forest pollen from about 85–60% of the pollen sum. This oscillation could reflect a slightly drier period in Colombia and it aligns with the inferred dry peak from El Valle. A period of cooling in the initiation of the last glacial period resulting in further southern migration of the ITCZ could account for both patterns. The changes of MIS5e and d seen in Panama and Colombia were minor compared with events further south in Bolivia and Peru where Lake Titicaca was reduced by 85% (D’Agostino et al. 2002; Fritz et al. 2012; Gosling et al. 2008; Hanselman et al. 2011).
El Valle in the last glacial period
The EV88 and EV90 fossil diatom records represent slightly different lake environments, while still representing El Valle during glacial times. EV90 represents a slightly deeper lake environment as it was cored 50 m further out from the shoreline than EV88, and therefore presents an A. granulata abundance similar to the EV08 core taken from the center of the basin. EV88 represents El Valle closer to the shoreline accounting for higher abundances of C. meneghiniana, U. delicatissima, and other benthic species. Their subsequent discussion accounts for this variation.
After lake levels began to increase at the end of MIS5e, an open water body was maintained at El Valle until approximately 22 ka. The decline in Ulnaria spp. and increase in C. meneghiniana suggested higher salinity from shallowing and eutrophication of the system. Some drier episodes were suggested by spikes of C. meneghiniana with the appearance of the subaerial diatom Luticola mutica (Kützing) Mann and the benthic diatom Gomphonema affine Kützing at 30 m depth. At other sites, the presence of these diatoms indicated shallow conditions (Vélez et al. 2005), shorelines closer to the coring point, or the increased erosion of wetland soils into the basin (Tibby 2001). At Lake Petén Itzá, a much drier setting than El Valle, there was an overall decrease in moisture availability through the early and mid-glacial period, and evidence of wet-dry cycles (Correa-Metrio et al. 2012; Hodell et al. 2008).
Further analysis of the El Valle glacial record is warranted as the abrupt spikes of the planktonic diatom, D. stelligera, could indicate more stratified conditions. In turn, C. meneghiniana, oscillating occasionally with D. stelligera could indicate drier periods with relatively higher salinity. Abrupt wet-dry cycles such as these have been described from Lake Petén Itzá, Guatemala (Correa-Metrio et al. 2012; Hodell et al. 2008), the Cariaco Basin in Venezuela (Peterson and Haug 2006), and Lake Fúquene, Colombia (Groot et al. 2011). At these locations, such abrupt wet-dry signatures align to Dansgaard–Oeschger cycles or Heinrich events (Dansgaard et al. 1993; Heinrich 1988) driven by Atlantic SST variability. It appears probable that El Valle, with a higher resolution glacial record and improved chronological control, would reveal a similar archive demonstrating the linkage between Central American climates and Atlantic SSTs.
The period between 22 and 15 ka was marked by a sharp increase in benthic diatom species. The fossil pollen record from this core section indicated the presence of a Typha swamp during this phase (Bush and Colinvaux 1990). The upper 5 m in EV88 were the only depths in any El Valle record where benthic species accounted for greater than 50% of the assemblage indicating either a drier climate or a hydrarch succession inducing shallowing within the last glacial maximum in Panama.
The diatom record from the ice-age sediments of El Valle revealed an overall mesic climate system with brief events that suggested substantial shallowing. Such climatic oscillations were consistent with sea-surface temperature-forced precipitation change in Central America rather than the broad characterization of glacial versus interglacial conditions implicit in Haffer’s refugial hypothesis (Haffer 1969).
Conclusions
The long-term dominance of planktonic over benthic diatoms in the Paleolake of El Valle suggests that Central Panama did not experience profound or protracted drought between approximately 137 and 30 ka. Toward the end of the last interglacial, between about 125 and 120 ka, water levels were lowered and El Valle was drier than at any time until the last glacial maximum. Throughout the majority of the ice-age at El Valle, a well-mixed lake existed within the basin. More stratified lake conditions and some lowstands were suggested that could have aligned with Dansgaard–Oeschger cycles or Heinrich Events in the Atlantic Ocean. Though likely, based on other records, a more detailed record is needed before this Atlantic Ocean—tropical climate linkage can be confirmed. The last glacial maximum at El Valle was probably drier than modern conditions, although infilling of the basin as a result of hydrarch succession cannot be ruled out, especially as the basin did not support a Holocene lake.
A more southern position of the ITCZ and cooler Atlantic SST produced moist conditions in the glacial that do not support the long-lasting ice-age aridity proposed by refugialists. The continued presence of a lake at El Valle throughout the last interglacial and glacial periods displays the persistence of humid Panamanian environments and the probable longevity of premontane settings as forested environments. Our data suggest that characterizing the past interglacial as being simply wet or dry is misleading. In Panama, the last interglacial was a period of overall drying, but witnessed considerable climatic heterogeneity.
References
ANSP (2012) Algae image database, phycology section. Patrick Center for Environmental Research, The Academy of Natural Sciences. https://diatom.ansp.org/algae_image/
Baker PA, Fritz SC (2015) Nature and causes of Quaternary climate variation of tropical South America. Quat Sci Rev 124:31–47
Bar-Matthews M, Ayalon A, Gilmour M, Matthews A, Hawkesworth CJ (2003) Sea–land oxygen isotopic relationships from planktonic foraminifera and speleothems in the Eastern Mediterranean region and their implication for paleorainfall during interglacial intervals. Geochim Cosmochim Acta 67:3181–3199
Battarbee RW (1986) Handbook of holocene palaeoecology and palaeohydrology. Wiley, New York
Blaauw M, Christen JA (2011) Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal 6(3):457–474
Bradbury JP, Leyden BW, Salgado-Labouriau ML, Lewis WM Jr, Schubert C, Binford M, Frey DG, Whitehead DR, Weibezahn FH (1981) Late quaternary environmental history of Lake Valencia, Venezuela. Science 214:1299–1305
Bush MB, Colinvaux PA (1990) A long record of climatic and vegetation change in lowland Panama. J Veg Sci 1:105–119
Bush MB, Piperno DR, Colinvaux PA, de Oliveira PE, Krissek LA, Miller MC, Rowe WE (1992) A 14 300-yr paleoecological profile of a lowland tropical lake in Panama. Ecol Monogr 62:251–275
Bush MB, Miller MC, de Oliveira PE, Colinvaux PA (2002) Orbital forcing signal in sediments of two Amazonian lakes. J Paleolimnol 27:341–352
Bush MB, Hanselman JA, Gosling WD (2010) Non-linear climate change and Andean feedbacks: an imminent turning point? Glob Change Biol 16:3223–3232
Bush MB, Gosling WD, Colinvaux PA (2011) Climate change in the lowlands of the Amazon Basin. In: Bush MB, Flenley JR, Gosling WD (eds) Tropical rainforest responses to climate change, 2nd edn. Praxis Springer, Chichester, pp 61–84
CAPE (2006) Last interglacial Arctic warmth confirms polar amplification of climate change. Quat Sci Rev 25:1383–1400
Cárdenes-Sandí GM (2015) Late quaternary vegetation and climate change in Central America. Florida Institute of Technology, Ann Arbor, p 258
Cheng H, Edwards RL, Sinha A, Spötl C, Yi L, Chen S, Kelly M, Kathayat G, Wang X, Li X (2016) The Asian monsoon over the past 640,000 years and ice age terminations. Nature 534:640–646
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18:117–143
Coates AG, Obando JA (1996) The geologic evolution of the Central American Isthmus. In: Jackson JBC, Budd AF, Coates AG (eds) Evolution and environment in tropical America. The University of Chicago Press, pp 21–56
Colinvaux PA, De Oliveira PE (2000) Palaeoecology and climate of the Amazon basin during the last glacial cycle. J Quat Sci 15:347–356
Colinvaux P, Irion G, Räsänen M, Bush M, De Mello JN (2001) A paradigm to be discarded: geological and paleoecological data falsify the Haffer and Prance refuge hypothesis of Amazonian speciation. Amazoniana 16:609–646
Correa-Metrio A, Bush MB, Cabrera KR, Sully S, Brenner M, Hodell DA, Escobar J, Guilderson T (2012) Rapid climate change and no-analog vegetation in lowland Central America during the last 86,000 years. Quat Sci Rev 38:63–75
Cruz FW Jr, Burns SJ, Jercinovic M, Karmann I, Sharp WD, Vuille M (2007) Evidence of rainfall variations in southern Brazil from trace element ratios (Mg/Ca and Sr/Ca) in a Late Pleistocene stalagmite. Geochim Cosmochim Acta 71:2250–2263
D’Agostino K, Seltzer GO, Baker PA, Fritz SC, Dunbar RB (2002) Late-quaternary lowstands of Lake Titicaca: evidence from high-resolution seismic data. Palaeogeogr Palaeocl 179:97–111
Dansgaard W, Johnsen SJ, Clausen HB, Dahl-Jensen D, Gundestrup NS, Hammer CU, Hvidberg CS, Steffensen JP, Sveinbjornsdottir AE, Jouzel J, Bond G (1993) Evidence for general instability of past climate from a 250-kyr ice-core record. Nature 364:218–220
ETESA (2009) Mapas Hidrometeorologicos. Datos Climáticos Históricos. Pixel Media Publicidad. http://www.hidromet.com.pa/clima_historicos.php?sensor=1
Fritz SC, Cumming BF, Gasse F, Laird KR (1999) Diatoms as indicators of hydrologic and climatic change in saline lakes. In: Stoermer EF, Smol JP (eds) The diatoms: applications for the environmental and earth sciences. Cambridge University Press, pp 41–72
Fritz SC, Baker PA, Seltzer GO, Ballantyne A, Tapia PM, Cheng H, Edwards RL (2007) Quaternary glaciation and hydrologic variation in the South American tropics as reconstructed from the Lake Titicaca drilling project. Quat Res 68:410–420
Fritz SC, Baker P, Tapia P, Spanbauer T, Westover K (2012) Evolution of the Lake Titicaca basin and its diatom flora over the last ~ 370,000 years. Palaeogeogr Palaeocl 317:93–103
Gibson KA, Peterson LC (2014) A 0.6 million year record of millennial-scale climate variability in the tropics. Geophys Res Lett 41:969–975
Gosling WD, Bush MB, Hanselman JA, Chepstow-Lusty A (2008) Glacial-interglacial changes in moisture balance and the impact on vegetation in the southern hemisphere tropical Andes (Bolivia/Peru). Palaeogeogr Palaeocl 259:35–50
Grimm EC (1987) CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput Geosci 13:13–35
Groot MHM, Bogotá RG, Lourens LJ, Hooghiemstra H, Vriend M, Berrio JC, Tuenter E, Van der Plicht J, Van Geel B, Ziegler M et al (2011) Ultra-high resolution pollen record from the northern Andes reveals rapid shifts in montane climates within the last two glacial cycle. Clim Past 7:299–316
Guiry MD, Guiry GM (2015) AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. http://www.algaebase.org
Haberyan KA, Horn SP, Cumming BF (1997) Diatom assemblages from Costa Rican lakes: an initial ecological assessment. J Paleolimnol 17:263–294
Haffer J (1969) Speciation in Amazonian forest birds. Science 165:131–137
Haffer J, Prance GT (2001) Climatic forcing of evolution in Amazonia during the Cenozoic: on the refuge theory of biotic differentiation. Amazoniana 16:579–605
Hanselman JA, Gosling WD, Paduano GM, Bush MB (2005) Contrasting pollen histories of MIS 5e and the Holocene from Lake Titicaca (Bolivia/Peru). J Quat Sci 20:663–670
Hanselman JA, Bush MB, Gosling WD, Collins A, Knox C, Baker PA, Fritz SC (2011) A 370,000-year record of vegetation and fire history around Lake Titicaca (Bolivia/Peru). Palaeogeogr Palaeocl 305:201–214
Haug GH, Gunther D, Peterson LC, Sigman DM, Hughen KA, Aeschlimann B (2003) Climate and the collapse of Maya civilization. Science 299:1731–1734
Heinrich H (1988) Origin and consequences of cyclic ice rafting in the northeast Atlantic Ocean during the past 130,000 years. Quat Res 29:142–152
Heiri O, Lotter AF, Lemcke G (2001) Loss on ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results. J Paleolimnol 25:101–110
Hidalgo PJ, Vogel TA, Rooney TO, Currier RM, Layer PW (2011) Origin of silicic volcanism in the Panamanian arc: evidence for a two-stage fractionation process at El Valle volcano. Contrib Mineral Petrol 162:1115–1138
Hill MO (1979) DECORANA—A FORTRAN program for detrended correspondence analysis and reciprocal averaging. Annu Rev Ecol Ebol Syst. Cornell University, New York
Hodell DA, Anselmetti FS, Ariztegui D, Brenner M, Curtis JH, Gilli A, Grzesik DA, Guilderson TJ, Muller AD, Bush MB, Correa-Metrio A, Escobar J, Kutterolf S (2008) An 85-ka record of climate change in lowland Central America. Quat Sci Rev 27:1152–1165
Hooghiemstra H (1984) Vegetational and climatic history of the high plain of Bogota, Colombia. Dissertaciones Botanicae 79, J. Cramer, Vaduz
Hooghiemstra H (2002) The dynamic rainforest ecosystem on geological, quaternary and human time scales. In: Verweij P (ed) Understanding and capturing the multiple values of tropical forest. Tropenbos International, Wageningen, pp 7–19
Hooghiemstra H, van der Hammen T (1998) Neogene and quaternary development of the neotropical rain forest: the refugia hypothesis, and a literature review. Earth Sci Rev 44:147–183
Kawamura K, Parrenin F, Lisiecki L, Uemura R, Vimeux F, Severinghaus JP, Hutterli MA, Nakazawa T, Aoki S, Jouzel J (2007) Northern Hemisphere forcing of climatic cycles in Antarctica over the past 360,000 years. Nature 448:912–916
Kilham P, Kilham SS, Hecky RE (1986) Hypothesized resource relationships among African planktonic diatoms. Limnol Oceanogr 31:1169–1181
Laskar J, Robutel P, Joutel F, Gastineau M, Correia A, Levrard B (2004) A long-term numerical solution for the insolation quantities of the Earth. Astron Astrophys 428:261–285
Ledru M-P, Rousseau DD, Cruz FW Jr., Riccomini C, Karmann I, Martin L (2005) Paleoclimate changes during the last 100,000 yr from a record in the Brazilian Atlantic rainforest region and interhemispheric comparison. Quat Res 64:444–450
Maher BA, Hu M (2006) A high-resolution record of Holocene rainfall variations from the western Chinese Loess Plateau: antiphase behaviour of the African/Indian and East Asian summer monsoons. Holocene 16:309–319
McIntire CD (1968) Structural characteristics of benthic algal communities in laboratory streams. Ecology 49:520–537
Nikolova I, Yin Q, Berger A, Singh UK, Karami M (2013) The last interglacial (Eemian) climate simulated by LOVECLIM and CCSM3. Clim Past 9:1789
Owen RB, Renaut RW, Scott JJ, Potts R, Behrensmeyer AK (2009) Wetland sedimentation and associated diatoms in the Pleistocene Olorgesailie Basin, southern Kenya Rift Valley. Sediment Geol 222:124–137
Paillard D, Labeyrie L, Yiou P (1996) Macintosh program performs time-series analysis. Eos Trans Am Geophys Union 77:379
Patrick R, Reimer CW (1966) The diatoms of the United States, vol 1. The Academy of Natural Sciences of Philadelphia, Philadelphia
Patrick R, Reimer CW (1975) The diatoms of the United States, vol 2. The Academy of Natural Sciences of Philadelphia, Philadelphia
Pennington RT, Lavin M, Prado DE, Pendry CA, Pell SK, Butterworth CA (2004) Historical climate change and speciation: neotropical seasonally dry forest plants show patterns of both Tertiary and Quaternary diversification. Philos Trans R Soc B 359:515–538
Pérez M, Bonilla S, Martínez G (1999) Phytoplankton community of a polymictic reservoir, La Plata River basin, Uruguay. Rev Bras Biol 59:535–541
Peterson LC, Haug GH (2006) Variability in the mean latitude of the Atlantic Intertropical Convergence Zone as recorded by riverine input of sediments to the Cariaco Basin (Venezuela). Palaeogeogr Palaeocl 234:97–113
Reimer PJ, Bard E, Bayliss A, Beck JW, Blackwell PG, Bronk Ramsey C, Buck CE, Cheng H, Edwards RL, Friedrich M (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55:1869–1887
Riboulleau A, Bout-Roumazeilles V, Tribovillard N (2014) Controls on detrital sedimentation in the Cariaco Basin during the last climatic cycle: insight from clay minerals. Quat Sci Rev 94:62–73
Round FE, Crawford RM, Mann DG (1990) The diatoms: biology and morphology of the genera. Cambridge University Press, Cambridge
Seltzer GO, Cross S, Baker P, Dunbar R, Fritz S (1998) High-resolution seismic reflection profiles from Lake Titicaca, Peru/Bolivia. Evidence for Holocene aridity in the tropical Andes. Geology 26:167–170
Spaulding SA, Lubinski DJ, Potapova M (2010) Diatoms of the United States. http://westerndiatoms.colorado.edu
Stebbins GL (1974) Flowering plants: evolution above the species level. The Belknap Press of Harvard University Press, Cambridge
Sublette Mosblech NA, Chepstow-Lusty A, Valencia BG, Bush MB (2012) Anthropogenic control of late-Holocene landscapes in the Cuzco region, Peru. Holocene 22:1361–1372
Tibby J (2001) Diatoms as indicators of sedimentary processes in Burrinjuck reservoir, New South Wales, Australia. Quat Int 83:245–256
Tremarin PI, Ludwig TAV, Torgan LC (2012) Ultrastructure of Aulacoseira brasiliensis sp. nov. (Coscinodiscophyceae) and comparison with related species. Fottea 12:171–188
Tzedakis C (2003) Timing and duration of Last Interglacial conditions in Europe: a chronicle of a changing chronology. Quat Sci Rev 22:763–768
Van der Hammen T (1974) The Pleistocene changes of vegetation and climate in tropical South America. J Biogeogr 1:3–26
Van der Hammen T, Absy ML (1994) Amazonia during the last glacial. Palaeogeogr Palaeocl 109:247–261
Van der Hammen T, Hooghiemstra H (2003) Interglacial–glacial Fuquene-3 pollen record from Colombia: an Eemian to Holocene climate record. Glob Planet Change 36:181–199
Vélez M, Berrio J, Hooghiemstra H, Metcalfe S, Marchant R (2005) Palaeoenvironmental changes during the last ca. 8590 calibrated yr (7800 radiocarbon yr) in the dry forest ecosystem of the Patía Valley, Southern Colombian Andes: a multiproxy approach. Palaeogeogr Palaeocl 216:279–302
Wang X, Edwards RL, Auler AS, Cheng H, Kong X, Wang Y, Cruz FW, Dorale JA, Chiang H-W (2017) Hydroclimate changes across the Amazon lowlands over the past 45,000 years. Nature 541:204–207
Weldeab S, Lea DW, Schneider RR, Andersen N (2007) 155,000 years of West African monsoon and ocean thermal evolution. Science 316:1303–1307
Wiens JJ, Donoghue MJ (2004) Historical biogeography, ecology and species richness. Trends Ecol Evol 19:639–644
Yarincik KM, Murray RW, Lyons TW, Peterson LC, Haug GH (2000) Oxygenation history of bottom waters in the Cariaco Basin, Venezuela, over the past 578,000 years: results from redox-sensitive metals (Mo, V, Mn, and Fe). Paleoceanography 15:593–604
Acknowledgements
Drilling of the 2008 El Valle core was made possible through a grant from the National Science Foundation (NSF-0902864). We thank all those involved in the drilling process of the EV08 core including Richard Wharry, Diana Ochoa, Andrés Gómez and Javier Luque. We thank Carlos Jaramillo for his help in transporting the core. William Gosling, Louise Thomas, and Peter van Calsteren of the Open University, U.K. are thanked for their assistance in providing the initial U/Th age. We would also like to thank the reviewers for their helpful insights into the composition of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
10933_2017_6_MOESM1_ESM.jpg
ESM1: Most common diatom species in core EV08 from El Valle, Panama. Scale bar is equal to 10 μm a) Ulnaria acus (Kützing) Aboal b)Ulnaria ulna (Nitzsch) Compére c) Aulacoseira agassizii Ostenfeld d) Cyclotella meneghiniana Kützing e) Navicula radiosa Kützing f) Nitzschia amphibia Grunow g) Achnanthidium minutissimum (Kützing) Czarnecki h) Aulacoseira granulata (Ehrenberg) Simonsen (JPEG 899 kb)
10933_2017_6_MOESM3_ESM.jpg
ESM3: EV08 Quercus record against the EV88 Quercus record. a) EV08 Quercus curve against depth. The 234U/230Th date for this core is indicated by (*). Quercus absence is indicated by the red section with the new chronology date indicated with and arrow. b) EV88 Quercus curve against depth. The 234U/230Th dates for this core is indicated by (*). Quercus absence is indicated by the red section. Quercus presence from extended counts are indicated by (+). (JPEG 388 kb)
Rights and permissions
About this article

{kind=link}
{kind=link}
Cite this article
Shadik, C.R., Cárdenes-Sandí, G.M., Correa-Metrio, A. et al. Glacial and interglacials in the Neotropics: a 130,000-year diatom record from central Panama. J Paleolimnol 58, 497–510 (2017). https://doi.org/10.1007/s10933-017-0006-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-017-0006-8