
Abstract
A large seabird colony of northern fulmars (Fulmarus glacialis) at Cape Vera on Devon Island in the Canadian high Arctic has been shown to act as an effective biological pump, transporting large quantities of marine-derived nutrients and contaminants to the ponds beneath their nesting sites. Previous research has shown that seabirds also have the potential to elevate radionuclide concentrations in their immediate surroundings. Here, we assess the influence of the Cape Vera seabird colony on the 210Pb budgets of six affected ponds and one control site, and discuss potential implications with respect to 210Pb dating methodologies. In general, fluxes of 210Pb at Cape Vera were similar to, or less than, those measured at other Arctic locations not influenced by seabirds. Plots of unsupported 210Pb activity versus depth showed a number of non-monotonic features in all sediment cores, necessitating the use of the constant-rate-of-supply (CRS) model to derive age-depth profiles. We conclude that seabird activity has not dramatically increased the 210Pb flux to the ponds at Cape Vera. Overall, the low flux of excess 210Pb made accurate dating problematic for some of the study cores, necessitating the use of independent dating markers (e.g., 137Cs and PCB concentrations) to verify the reliability of the chronologies.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The role of biovectors in the global transport of contaminants is receiving increased attention, as research has shown that the behaviours and lifecycles of certain animals can focus industrially produced compounds into foodwebs at concentrations far beyond those contributed by abiotic processes alone (Blais et al. 2007). Recent research at Cape Vera on Devon Island in the Canadian high Arctic has shown that the guano and other ornithogenic inputs generated by a large seabird colony have dramatically altered the physical, chemical and biological characteristics of their immediate surroundings. For example, Blais et al. (2005) found concentrations of contaminants up to 60-fold higher than background levels in the surface sediments of ponds located beneath the seabird nesting sites. At present, our labs are conducting several research projects on the ponds at Cape Vera, which, in addition to the contaminant work, includes examining the impact of seabirds on water quality and aquatic biota (Keatley 2007). In particular, we aim to use paleolimnology to reconstruct seabird population dynamics and their influence on nutrient and contaminant budgets over long-term timescales. Naturally, the development of accurate sediment chronologies is key to interpreting such temporal records.
The radioisotope 210Pb (half-life 22.3 years) is commonly used to date recent sediments (up to ∼150 years), and although its use is commonplace in Arctic regions, it can be problematic due to issues relating to persistent lake ice cover, permafrost suppression of 222Rn (a 210Pb parent nuclide), and reduced atmospheric scavenging due to low precipitation (Wolfe et al. 2004). All of these factors contribute to low initial 210Pb concentrations in lacustrine sediments, which can reduce the time period usable by this geochronological tool and lead to large dating errors.
An issue that must be taken into consideration when using 210Pb radiochronology at Cape Vera is the potential of seabirds to enrich pond sediments with radionuclides, via similar processes by which they concentrate contaminants and nutrients. Although 210Pb is not bioconcentrated in marine systems, it is present in seawater and may be ingested by the prey species of seabirds. Thus, given the naturally low excess 210Pb levels in Arctic regions, a large congregation of seabirds has the potential to increase 210Pb flux to the surrounding environment. For example, Dowdall et al. (2005) recorded enrichment factors of 8 for 137Cs, 5 for 238U, and 2 for 226Ra in soils near a seabird colony at Kongsfjorden, in the high Arctic archipelago of Svalbard. Although it was not clear whether these seabirds were increasing radionuclide concentrations directly through their feces or indirectly by increasing the organic content of the soil thereby enhancing its ability to retain a number of different radionuclides, the impact of the seabirds on the radioecology of the area was unquestionable.
The two most commonly used 210Pb dating models are the constant-rate-of-supply (CRS) and constant-initial-concentration (CIC) models (Appleby 2001), with the former being the most widely used (Binford et al. 1993; Blais et al. 1995). In general, the CIC model assumes a constant initial 210Pb concentration in the sediments regardless of accumulation rates, whereas the CRS model assumes that the rate of deposition of unsupported 210Pb from the atmosphere is constant. The most appropriate model for dating Cape Vera sediments will depend on several factors including whether: (1) sediment accumulation rates are closely tied to the amount of seabird-derived nutrients, (2) seabird subsidies contain significant amounts of 210Pb, and (3) the amount of seabird-derived nutrients and 210Pb reaching the ponds vary in proportion to each other.
Here we investigate the influence of seabird activity on the 210Pb budgets of pond sediments at Cape Vera, and discuss potential implications for 210Pb radiochronology. We address two central questions. First, have seabirds considerably elevated the sedimentary 210Pb flux at Cape Vera relative to other high latitude sites that do not support seabird colonies? Second, which 210Pb-dating model, CRS or CIC, is most appropriate to date the sediment cores from the seabird-influenced ponds? To address the latter question, we examine features in the unsupported 210Pb activity versus depth profiles, such as the presence/absence of exponential declines and non-monotonic changes. Following the selection of the most appropriate 210Pb-dating model, we then objectively evaluate the reliability of the chronology using two independent dating markers, specifically 137Cs and polychlorinated biphenyls (PCBs).
Site description
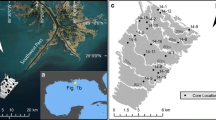
Cape Vera (76°15′ N, 89°15′ W) is located on Devon Island, Nunavut, in the Canadian high Arctic (Fig. 1). The landscape of this region is dominated by dolostone/limestone cliffs that rise abruptly ∼300 m asl. These cliffs provide ideal nesting habitats for ∼20,000 northern fulmars (Fulmarus glacialis) that breed here between May and September each year (Gaston et al. 2006). Our study ponds are located on a small foreland that stretches along the length of the cliffs (Fig. 1). Each breeding season, these ponds receive large amounts of waste products (e.g., guano, regurgitated stomach contents, carcasses) from the nesting birds. The seabirds represent an important link between marine and terrestrial environments because the waste products they generate are ultimately derived from the ocean, which is their only feeding ground. Thus, the seabirds act as an effective biological pump, enriching the ponds beneath their nesting sites with marine-derived nutrients (Keatley 2007) and contaminants (Blais et al. 2005). Detailed descriptions of the study ponds and their limnological characteristics are given in Keatley (2007).

Map showing the location of (a) Cape Vera on Devon Island in the Canadian Arctic archipelago, and (b) the study sites and the main breeding area (i.e., guano source) at the fulmar colony (dark grey-shaded region)
Methods
Sediment core recovery
Sediment cores were recovered from seven ponds during July 2005 and 2006 by pushing plexiglass tubes (3.75 cm diameter) directly into the sediments and plugging the tube bottoms with specially designed bungs, as in standard gravity coring operations (Glew 1989). All cores were sectioned on-site into 0.5-cm intervals (except for CV-5 which was sectioned at 1-cm intervals) using a Glew (1988) extruder. All ponds were assigned unofficial names using the prefix CV and a number (e.g., CV-5, -6, -7, -9, -9a, -20, -22). All sites are located within the immediate vicinity of the seabird breeding site, except for CV-22, which is located at Cape Hawes, a region with no modern seabird activity ∼10 km from Cape Vera (Fig. 1).
210Pb and 137Cs determinations
The sediment subsamples to be 210Pb-dated were lyophilized, ground to a fine powder using a mortar and pestle, and placed in centrifuge tubes (outer diameter = 1.5 cm) up to a height of 2 cm, corresponding to ∼1–2 g dry mass. The sediments were allowed to settle before being sealed with epoxy and left for three weeks to attain radioactive equilibrium between the in-situ 226Ra and its decay products that are used to estimate supported 210Pb activity. The equilibrated samples were counted on a digital, high purity germanium spectrometer (DSPec, Ortec®) with a detector consisting of a germanium crystal with lithium diffused electrodes. All samples were counted for at least 23 h to ensure a sufficient signal, with a minimum of 400–500 counts per radioisotope to obtain a good signal to noise ratio. The resulting spectrum files showed 210Pb activity with a peak at 46.5 keV. 226Ra activity was determined by gamma-ray emissions of its daughter isotope 214Pb at 295 and 352 keV. The activity (A) of a sample was calculated by:
where, N is the number of counts in the peak; e is the detector efficiency; Y is the yield of photons of an energy E; and t is the count time. All spectrum files were analyzed using a DOS-based software program developed and provided by Peter Appleby (University of Liverpool, UK), which included efficiency calculations and corrections for self absorption (Appleby 2001).
137Cs was measured simultaneously with 210Pb, and had an activity peak at 662 keV. 137Cs is an artificial radionuclide associated with above ground testing of nuclear weapons, which in North America began in the late 1940s and peaked in 1963, just prior to the signing of a nuclear test ban treaty. Thus, the onset of detectable 137Cs should correspond to the late-1940s/early-1950s, and its peak activity should denote 1963, the time of its maximum atmospheric fallout (Appleby 2001).
Polychlorinated biphenyls (PCBs)
PCBs were analyzed on a Hewlett-Packard 6890 series II gas chromatograph with a 63Ni micro electron-capture detector, using splitless injection with an inlet temperature of 250°C. Details on sample preparation, instrument specifications and calibration, and compound identification and quantification are given in Krümmel et al. (2005). PCB measurements provide additional chronological markers, as these compounds were first produced in 1927, with widespread industrial usage occurring in the mid-1950s. In spite of this, we do not use the first onset of PCBs to denote 1927, as these compounds are so pervasive in the environment that some degree of contamination is unavoidable (Blais and Muir 2001). Although we perform blank subtractions on our extracts, this only accounts for contamination encountered in the laboratory. Examination of the PCB profiles in all sediment cores reveals trace levels of these compounds even in the deepest (pre-1900s) sediments (Fig. 2e). However, each of the PCB profiles also records a marked rise in concentration at some point in time. Under the assumption that all sediment samples were exposed to the same degree of contamination during field sampling and transport (all sediment cores were processed and stored following identical procedures), we use the timing of the marked rise in PCB concentration to demarcate the early-to-mid-1950s, when their industrial usage became widespread.

Plots showing (a) total and supported 210Pb activity (as 226Ra), (b) unsupported 210Pb activity, (c) CRS-derived age-depth profiles, (d) 137Cs activity, and (e) total PCB concentration for all sediment cores. The dates shown in (d) denote the CRS dates that correspond to the peak 137Cs activities. The arrows shown in (e) indicate the timing of the marked rise in PCB concentrations
Results and discussion
The influence of seabirds
The seabird colony at Cape Vera is clearly transporting large quantities of marine-derived nutrients and contaminants to the ponds beneath their breeding grounds (Blais et al. 2005; Keatley 2007). Seabirds also have the potential to enrich pond sediments with 210Pb, as it is present in seawater and may be ingested by their prey. However, 210Pb fluxes recorded at Cape Vera are amongst the lowest values recorded throughout the Arctic (Table 1). The comparatively low 210Pb fluxes recorded at Cape Vera (range from 2.2 to 12.5 Bq m2 yr−1) indicate that seabirds have only negligible, if any, influence on the lacustrine 210Pb budgets. As further proof, CV-22, the pond located 10 km away from the nearest seabird activity, had the third highest 210Pb flux in our Cape Vera dataset (Table 1). The variations in 210Pb flux among our sites (mean = 7.0 ± 3.8 Bq m2 yr−1) may be related to sediment focusing, as this process is known to influence lacustrine 210Pb flux, both within and among lakes (Fitzgerald et al. 2005). However, given the shallow nature of these waterbodies (<2 m deep), and their gently sloping sides with large flat bottoms, sediment focusing may be less important than other catchment related factors, such as differences in hydrology, drainage area and ground cover.
Our 210Pb flux data contrasts with previous research that showed seabird activity markedly elevated radionuclide levels in their immediate surroundings (Dowdall et al. 2005). One possible explanation for the low 210Pb fluxes recorded in our study ponds is that the increased organic content of the catchment via nutrient inputs from the birds has enhanced the capacity of the surrounding landscape to retain 210Pb. Thus, seabirds may be transporting 210Pb to Cape Vera at levels greater than the natural atmospheric flux, but little of this surplus 210Pb reaches the pond sediments as most of it is retained within the rich moss beds that characterize most of the seabird-impacted zone.
Core chronologies
Total 210Pb near-surface activities ranged from ∼100–160 Bq kg−1, and reached supported levels from near ∼2 to 10 cm depths (Fig. 2a). Mean supported 210Pb activities (measured as 214Pb) ranged from 3 (CV-20) to 33 (CV-7) Bq kg−1. All cores recorded a number of non-monotonic features in profiles of unsupported 210Pb versus depth (Fig. 2b). These stratigraphic reversals are permissible in CRS models, but not in CIC models as they necessarily imply changes in initial concentration. Thus, only the CRS model can be considered valid for establishing age-depth profiles in our study ponds. Although the unsupported/supported 210Pb horizon appears to have been contained in all sediment cores (except CV-20), we typically do not extend our chronologies beyond ∼1920, as the low activities produce dating errors that are unacceptably large (i.e., >30% of the inferred age). Below we discuss the CRS-derived chronologies in relation to independent chronological markers, namely 137Cs and PCBs.
Cores CV-5, CV-6, and CV-9a
The sediment cores from ponds CV-5, CV-6, and CV-9a display peak 137Cs concentrations corresponding to CRS-derived dates of 1976, 1965, and 1977, respectively (Fig. 2c). Thus, the 137Cs peak activities match reasonably well with the known timing of maximum 137Cs discharge to the atmosphere in 1963 (Appleby 2001). The initial onset of 137Cs activity predates 1920 in all three sediment cores (several decades before it was first released into the atmosphere by nuclear testing), indicating some degree of downward diffusion. Although diffusional processes appear to have influenced these 137Cs profiles, they typically do not cause displacement of peak concentrations, and thus peak 137Cs activity can still be used to demarcate 1963.
The marked increases in PCB concentrations recorded in CV-6 and CV-9a (denoted by arrows in Fig. 2e) correspond to CRS-derived dates of 1945 and 1951, respectively. The onset of these marked increases is generally consistent with the timing of widespread industrial usage of PCBs, which began in the early-to-mid-1950s. The PCB profile from CV-5 is more difficult to interpret (Fig. 2e). For example, although there is a slight rise near 7.25 cm depth (corresponding to a CRS date of 1948), there is a much larger increase near 2 cm depth (corresponding to a CRS date of 2002). The uncertainty as to which increase marks the true rise of PCBs at CV-5 makes it difficult to use this profile to independently validate the CRS chronology.
Agreement between 210Pb dates and independent markers within ±15 years may be considered to indicate reliable sediment chronologies, particularly in lake systems that pose challenges to dating (e.g., Schottler and Engstrom 2006). Thus, the generally good conformity between the 210Pb dates and other independent chronological markers indicates that the CRS model provides reliable age-depth profiles in cores CV-5, CV-6, and CV-9a.
Core CV-9
The CRS-derived dates for the CV-9 sediment core appear too young, based on the 137Cs and PCB data. For example, the initial onset and peak activity of the 137Cs profile in the CV-9 core corresponds to CRS dates of 1978 and 1990, respectively, and the marked rise in PCB concentration corresponds to a CRS date of 1978 (Fig. 2d, e). When independent radioisotopes do not validate each other, as in the case of CV-9, it is difficult to determine which one most accurately reflects the true age-depth profile. The 137Cs curve for CV-9 appears to be well resolved, displaying a ‘classic’ 137Cs profile including an initial onset (from zero), distinctive subsurface peak, and decline towards the surface. Likewise, the PCB profile also appears to be well resolved, showing a marked increase from near-zero values. The fact that the 137Cs and PCB data both indicate the CRS-derived chronology underestimates the “true” age-depth profile, gives credence to the use of these markers over the use of the 210Pb dates. Moreover, as discussed earlier, both 137Cs and PCBs are predicted to show initial increases in lake sediments beginning around the early 1950s. The correspondence between the first onset of 137Cs and marked rise in PCB concentration (Fig. 2d, e) lends further support to the reliability of these two markers over 210Pb. Thus, in the case of CV-9 we use the 137Cs peak at 3.25 cm to denote 1963 and the corresponding increases of 137Cs and PCBs at 6.25 cm to demarcate the early-to-mid-1950s.
Cores CV-7, CV-20, and CV-22
The dating markers measured on the cores from CV-7, CV-20, and CV-22 pose some challenges to interpretation of sediment chronology. In CV-7, total 210Pb concentrations reach supported levels within 5 cm depth, indicating very low sedimentation rates (Fig. 2a). Although the 137Cs profile appears well resolved, the peak at 3.25 cm corresponds to a CRS date of 1918, which, of course, is far too old to corroborate the 210Pb chronology. However, the 137Cs activity measured at 2.25 cm depth (corresponding to a CRS date of 1974) is only slightly less than at 3.25 cm (Fig. 2c). Given the slow sedimentation rates at CV-7, it is likely that the true 137Cs peak may lie somewhere between 2.25 and 3.25 cm depth, in which case the 137Cs peak would have good conformity with the 210Pb dates. Thus, despite the apparent offset with the 137Cs profile, the CRS chronology appears to approximate a realistic reflection of the true age-depth profile, although the CRS dates should be used with caution. The PCB profile in CV-7 shows a small rise near 2.75 cm depth, corresponding to a CRS date of 1946 (Fig. 2e), however, this PCB increase is so minor compared to the other cores that it cannot be used reliably to support the CRS chronology.
CV-20 is the only core in which excess 210Pb does not appear to decline to zero, indicating the unsupported/supported 210Pb horizon was not captured within this sediment core (Fig. 2a). We do not suspect mixing, as the diatom stratigraphy shows several small, but distinct, species changes that would not be recorded in a well-mixed core (Keatley 2007). The CRS chronology (Fig. 2c) was obtained by interpolating to background by extending a line from ln 210Pb activity to background as measured by 226Ra. Profiles of 137Cs and PCB concentrations do not show initial increases from near zero values, as in all other cores (Fig. 2d, e), indicating that the CV-20 core probably does not extend beyond ∼1950. Likewise, the 137Cs peak and CRS dates both indicate less than ∼50 years of sediment accumulation within this core; however, the relatively poor conformity between the two radionuclides (the 137Cs peak at 6.25 cm depth corresponds to a CRS date of 1985) suggests that any dates from this core should be interpreted cautiously.
In CV-22, both 210Pb and 137Cs radioisotopes indicate extremely slow sedimentation rates. For example, the maximum 137Cs concentration, demarcating the 1963-dating horizon, occurs in the uppermost sediment interval (Fig. 2d). Although this is not a ‘typical’ 137Cs profile, the minimal activity below 2 cm depth indicates that any diffusional processes are likely negligible. Total 210Pb concentrations appear to reach supported values within ∼2 cm depth (Fig. 2a), which severely limits the number of dates that can be included in the chronology (Fig. 2c). The 210Pb inventory for CV-22 is low compared to the other study ponds, and may be related to low 210Pb retention owing to minimal organic sedimentation. This finding is as expected, given that CV-22 is located 10 km away from the seabird colony at Cape Vera, and thus does not receive abundant seabird-derived nutrients as do the other study ponds. Likewise, the negligible PCB concentrations recorded at CV-22 compared to all other study ponds (Fig. 2e) supports the findings of Blais et al. (2005), that seabirds, not abiotic processes, are the main vectors of PCB transport to the ponds at Cape Vera. Thus, although longer-lived radioisotopes (e.g., 14C) are required to extend the CV-22 chronology beyond 2 cm depth, the 210Pb and 137Cs data both indicate slow sedimentation rates, similar to those recorded in non-seabird influenced ponds elsewhere in the Canadian high Arctic (e.g., Douglas et al. 1994).
Conclusions
We have shown that seabird activity has not greatly affected the 210Pb budgets of the pond sediments at Cape Vera. An area of future research is to measure 210Pb concentrations in mosses and organic material surrounding the affected ponds to determine whether the catchment is acting as a sink for 210Pb, and potentially other radionuclides. Typical of high Arctic regions, the low flux of excess 210Pb made dating problematic, but in combination with independent chronological markers (137Cs and PCBs), chronologies were established for most cores. The undeniable influence of seabird activity on the limnology of the Cape Vera ponds (e.g., Keatley 2007; Blais et al. 2005) suggests that paleolimnology can be used to track past seabird population dynamics and their influence on nutrient and contaminant levels over long-term timescales. The sediment chronologies established here are a crucial first step in that process.
References
Appleby PG (2001) Chronostratigraphic techniques in recent sediments. In: Last WM, Smol JP (eds) Tracking environmental change using lake sediments. Volume 1: Basin analysis, coring, and chronological techniques. Kluwer Acadamic Publishers, Dordrecht, The Netherlands, pp 172–203
Binford MW, Kahl JS, Norton SA (1993) Interpretation of 210Pb profiles and verification of the CRS dating model in the PIRLA project lake sediment cores. J Paleolimnol 9:275–296
Blais JM, Muir DCG (2001) Paleolimnological methods and applications for organic contaminants. In: Last B, Smol JP (eds) Developments in paleoenvironmental research, vol 2: physical and chemical approaches, chap 10. Kluwer Academic Press, Dordrecht, The Netherlands, pp 271–298
Blais JM, Kalff J, Cornett RJ, Evans RD (1995) Evaluation of 210Pb dating in lake sediments using stable Pb, Ambrosia pollen, and 137Cs. J Paleolimnol 13:169–178
Blais JM, Kimpe LE, McMahon D, Keatley B, Mallory ML, Douglas MSV, Smol JP (2005) Arctic seabirds transport marine-derived contaminants. Science 309:445. doi:10.1126/science.1112658
Blais JM, MacDonald RW, Mackay D, Webster E, Harvey C, Smol JP (2007) Biologically mediated transport of contaminants to aquatic systems. Environ Sci Technol 41:1075–1084
Douglas MSV, Smol JP, Blake W Jr (1994) Marked post-18th century environmental change in high-arctic ecosystems. Science 266:416–419
Dowdall M, Gwynn JP, Gabrielsen GW, Lind B (2005) Assessment of elevated radionuclide levels in soils associated with an avian colony in a high arctic environment. Soil Sediment Contam 14:1–11
Fitzgerald WF, Engstrom DR, Lamborg CH, Tseng C-M, Balcom PH, Hammerschmidt CR (2005) Modern and historic atmospheric mercury fluxes in northern Alaska: global sources and Arctic depletion. Environ Sci Technol 39:557–568
Gaston AJ, Mallory ML, Gilchrist HG, O’Donovan K (2006) Status, trends and attendance patterns of the northern fulmar Fulmarus glacialis in Nunavut, Canada. Arctic 59:165–178
Glew JR (1988) A portable extruding device for close interval sectioning of unconsolidated core samples. J Paleolimnol 1:235–239
Glew JR (1989) A new trigger mechanism for sediment samplers. J Paleolimnol 2:241–243
Hermanson MH (1990) 210Pb and 137Cs chronology of sediments from small, shallow Arctic lakes. Geochim Cosmochim Acta 1443–1451
Keatley BE (2007) Limnological and paleolimnological investigations of environmental change in three distinct ecosystem types, Canadian High Arctic. PhD Thesis, Queen’s University at Kingston, Ontario, Canada, 332 pp
Krümmel EM, Gregory-Eaves I, Macdonald RW, Kimpe LE, Demers MJ, Smol JP, Finney B, Blais JM (2005) Concentrations and fluxes of salmon–derived polychlorinated biphenyls (PCBs) in lake sediments. Environ Sci Technol 39:7020–7026
Lockhart WL, Wilkinson P, Billeck BN, Danell RA, Hunt RV, Brunskill GJ, Delaronde J, St Louis V (1998) Fluxes of mercury to lake sediments in central and northern Canada inferred from dated sediment cores. Sci Total Environ 260:171–180
Lockhart WL, Stern G, Outridge P (2000) Mercury and other contaminants in a laminated sediment core from Devon Island. In: Kalhok S (ed) Synopsis of research carried out under the 1999–2000 northern contaminants program. Dept. of Indian Affairs and Northern Development, Ottawa, Canada, pp 104–112
Michelutti N, Wolfe AP, Vinebrooke RD, Rivard B, Briner J (2005) Recent primary production increases in arctic lakes. Geophys Res Lett 32:L19715. doi: 10.1029/2005GL023693
Michelutti N, Wolfe AP, Briner JP, Miller GH (2007) Climatically controlled chemical and biological development in Arctic lakes. J Geophys Res 112:G03002. doi:10.1029/2006JG000396
Schottler SP, Engstrom DR (2006) A chronological assessment of Lake Okeechobee (Florida) sediments using multiple dating markers. J Paleolimnol 36:19–36
Wolfe AP, Miller GH, Olsen CA, Forman SL, Doran PT, Holmgren SU (2004) Geochronology of high latitude lake sediments. In: Pienitz R, Douglas MSV, Smol JP (eds) Long-term environmental change in Arctic and Antarctic Lakes. Springer, The Netherlands, pp 19–52
Acknowledgements
This work was funded by Strategic and Discovery grants from the Natural Science and Engineering Research Council of Canada awarded to JMB, MSVD, and JPS, and a Northern Scientific Training Program (NSTP) grant to BEK. We thank the Polar Continental Shelf Project/Étude du Plateau Continental Polaire (PCSP/ÉPCP), NSTP and Environment Canada (CWS) for financial and logistical support in the field. The quality of this manuscript was greatly improved by two anonymous reviewers. This represents PCSP/ÉPCP contribution No. 03207.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Michelutti, N., Blais, J.M., Liu, H. et al. A test of the possible influence of seabird activity on the 210Pb flux in high Arctic ponds at Cape Vera, Devon Island, Nunavut: implications for radiochronology. J Paleolimnol 40, 783–791 (2008). https://doi.org/10.1007/s10933-008-9198-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-008-9198-2