
Abstract
Water chemistry and surface sediments were analyzed from 41 shallow lakes representing three previously-defined hydrological categories in the Slave River Delta, Northwest Territories, Canada, in order to identify relationships between hydrological and limnological conditions and their associations with recently deposited diatom assemblages. Evaporation-dominated lakes are physically removed from the influence of the Slave River, and are characterized by high alkalinity and high concentrations of nutrients and ions. In contrast, flood-dominated lakes tend to receive a pulse of floodwater from the Slave River during the spring thaw and have low alkalinity and low concentrations of most nutrients and ions. Exchange-dominated lakes are variably influenced by floodwaters from the Slave River and seiche events from Great Slave Lake throughout the spring thaw and open-water season, and are characterized by a broad array of limnological conditions that are largely dependent on the strength of the connection to these sources of floodwater. Specific diatom ‘indicator’ taxa have been identified that can discriminate these three hydrological lake categories. Evaporation-dominated lakes are associated with high relative abundance of common epiphytic diatom taxa, while diatoms indicative of flood- and exchange-dominated lakes span a wide range of habitat types (epiphytic, benthic) but also include unique planktonic diatoms (Stephanodiscus and Cyclostephanos taxa) that were not found in surface sediments of evaporation-dominated lakes. The planktonic diatom taxa originate from the Slave River, and thus are indicative of river influence. In complex, remote, freshwater ecosystems like the Slave River Delta, integration of results from hydrological and limnological approaches provides a necessary foundation to assess present, past and future hydroecological responses to changes in river discharge and climate.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Deltas are highly productive landscapes, and include important aquatic ecosystems that are responsive to spatial and temporal variability of river discharge and flooding (e.g. Lewis et al. 2000; Junk 2005). Due to strong control of river connection on the physical, geochemical and biological conditions of floodplain lakes (English et al. 1997; Hay et al. 1997; Lesack et al. 1998; Prowse and Conly 2001, 2002; Squires and Lesack 2001, 2002, 2003; Squires et al. 2002; Spears and Lesack 2006; Brock et al. 2007; Wolfe et al. 2007b), there is concern regarding the downstream effects of human (Nilsson et al. 2005) and hydro-climatic (Overpeck et al. 1997; Serreze et al. 2000) alteration of river hydrology on the ecological state of floodplain and delta lakes and wetlands (Prowse and Conly 2001; Junk 2005; Schindler and Donahue 2006; Schindler and Smol 2006). Consequently, there is increasing need to develop improved and scientifically rigorous methods for the assessment and prediction of potential impacts of multiple stressors on deltaic environments. This need is particularly acute for remote, northern deltas as there is limited scientific knowledge of ecosystem hydroecology, and the data that exist are usually too short and too sparse to distinguish the relative roles of natural and anthropogenic changes. Such is the case for the Slave River Delta, one of three major deltas within the Mackenzie River Drainage System in northern Canada, which is thought to have undergone considerable drying as a result of regulation of the Peace River upstream (English et al. 1997; Fig. 1). Periodic flooding has created a landscape with high biological productivity and diversity of plant species that provides important feeding, staging, and breeding habitats for a large number of waterfowl, muskrat, and other wildlife (English et al. 1997). The natural resources of this ecosystem are also of central importance to the livelihood and socio-cultural integrity of the indigenous community of Fort Resolution, NWT (Wolfe et al. 2007a).

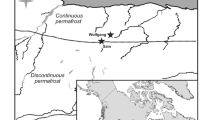
Location of the Slave River Delta and sampling sites, including rivers (R1, R2, R3) and Great Slave Lake (SD42, black triangles), and delta lakes (SD1–41) classified according to the hydrological categories of Brock et al. (2007): flood-dominated (grey circles), exchange-dominated (black circles), and evaporation-dominated (open circles) lakes
The limnological conditions in lakes of the Mackenzie Delta and the Peace-Athabasca Delta, two other major deltaic ecosystems in the Mackenzie River Drainage Basin, are strongly regulated by the frequency of flooding and the degree of connection between floodplain lakes and their associated rivers (Hay et al. 1997; Squires et al. 2002; Wolfe et al. 2007b). Although deltaic sediments may be challenging for paleoenvironmental interpretations due to their highly dynamic nature (e.g. Michelutti et al. 2001), they are providing important records of environmental change (Sakaguchi et al. 2006; Zalat and Servant Vildary 2007) and a wealth of information about past flood events and the dynamic hydrological, geochemical and biotic responses to such events (Hay et al. 1997, 2000; Hall et al. 2004; Wolfe et al. 2005, 2006). Biological communities have been shown to be useful paleo-indicators of hydroecological change in these environments, but developing scientifically robust interpretations requires understanding and quantifying contemporary relationships between basin hydrology, environmental conditions and biological communities of delta lakes (Hall et al. 2004; Wolfe et al. 2005, 2007b). Diatoms are particularly useful biomonitors for such studies, because they are sensitive to changes in physical, chemical and biological conditions and microhabitat availability (Hay et al. 1997; Hall et al. 2004). In the Peace-Athabasca Delta, for example, epiphytic taxa dominate in closed-drainage lakes, whereas planktonic taxa and non-epiphytic taxa (e.g. benthic and tychoplanktonic Fragilaria taxa) proliferate in turbid open-drainage lakes and under flood conditions (Hall et al. 2004). Previous studies in the Mackenzie Delta (Hay et al. 1997, 2000) and the Peace-Athabasca Delta (Hall et al. 2004) have found strong relationships between diatom communities and the hydrolimnological conditions of delta lakes, which are important for accurate interpretations of hydroecological changes from diatom assemblages analyzed in sediment cores. While no such study has been conducted for the Slave River Delta, extensive hydrological studies have been undertaken that have identified three main groups of lakes based on the major hydrological processes that control their water balances (Brock et al. 2007, detailed below).
In this study, we determine if hydrological differences among lakes in the Slave River Delta result in predictable differences in physical and chemical characteristics of lakes, and if these differences are related to the composition of diatom assemblages in recently-deposited surficial lake sediments. Our approach is to: (1) quantify relationships between basin hydrology and limnological conditions, and determine if limnological conditions differ among the three hydrological lake categories, (2) identify the main environmental gradients associated with the composition of diatom assemblages contained in surface sediments and determine whether diatom assemblage composition differs among the hydrological lake categories and (3) identify the diatom taxa that best discriminate among the hydrological lake categories. Results from this study provide a scientific framework for effective biomonitoring of hydroecological changes in the Slave River Delta and for quantifying roles of river hydrology, human impacts and climatic changes over decadal to millennial time scales from ongoing and future paleolimnological analyses.
Site description
The Slave River Delta is located at the mouth of the Slave River in the Northwest Territories (61°15′ N; 113°30′ W) where it enters Great Slave Lake (Fig. 1). The delta consists of a large wetland complex with numerous river channels, marshes, fens, bogs, swamps and forests. Scattered throughout this landscape are a multitude of small, shallow (<5 m) ponds and lakes (hereafter referred to collectively as lakes) that span broad hydrological gradients and support extensive macrophyte growth.
Including non-active areas of sedimentation, the delta is 170 km long by 70 km wide, extending north-northwest from the Slave River rapids at the south end to Great Slave Lake at the north end (Milburn and Prowse 1998). The entire delta covers an area of 8,300 km2, but much of it is no longer active (Vanderburgh and Smith 1988). The active part of the delta is arcuate, spans an area approximately 400 km2, and is prograding into Great Slave Lake through a system of active distributaries (Prowse et al. 2002). Natural levees in this area of the delta are between 0.1–1.5 m above Great Slave Lake low summer water levels, making lakes in the outer delta more susceptible to flooding by the Slave River, whereas the upstream relict portion of the delta has levee heights greater than 2.5 m which impede most river floodwaters (English et al. 1997). The extent and frequency of flooding can vary considerably, but periodic spring ice-jams are the main mechanism that promotes river flooding into Slave River Delta lakes (Prowse et al. 2002).
English et al. (1997) divided the active delta into biogeographical zones based on vegetation patterns and geomorphological differences (e.g. levee height), but this has limited application for assessing hydroecological changes in lakes because it assumes that all lakes within a zone are influenced by similar hydrological processes. More recently, Brock et al. (2007) examined the roles of major hydrological processes (including precipitation, snowmelt runoff, river flooding, Great Slave Lake seiche events and evaporation) on the water balances of lakes in the Slave River Delta using water isotope tracers and identified three distinct hydrological lake categories. The water balances of flood-dominated lakes, located in the active part of the delta, are strongly influenced by Slave River floodwater during the spring melt. Evaporation-dominated lakes, located in the older non-active part of the delta, receive spring snowmelt and evaporation becomes the over-riding process controlling lake water balances during the remainder of the open-water season. Exchange-dominated lakes are located along the Slave River and delta front adjacent to Great Slave Lake and receive periodic inputs throughout the spring and open-water seasons from channel connections with the Slave River or Great Slave Lake water during seiche events, resulting in variable water balances depending on the strength of these connections.
The hydrological classification by Brock et al. (2007) was based on seasonal isotopic data collected during thaw season 2003. Notably, Slave River discharge during spring break-up of 2003 was average compared to 46 years of gauge data at the nearest hydrometric station. This feature is important for our study, because both the hydrological classification (based on water samples collected in 2003), which is strongly influenced by Slave River spring discharge, and surface sediments used in this study (collected in September 2002), which integrate diatoms deposited over a few years, likely reflect average contemporary hydrological and hydroecological conditions of the delta.
Materials and methods
Sample collection and laboratory analyses
The water bodies in this study are the same as those sampled and analyzed by Brock et al. (2007). River sites include the Slave River (site R3) and two of its distributaries (sites R1 [East Channel] and R2 [Jean River]). Water samples from Great Slave Lake (SD42) were collected 100 m offshore of the outer delta to characterize the chemistry of seiche input waters to exchange-dominated basins. The 41 lakes sampled encompass the three hydrological categories (flood-dominated [n = 10], evaporation-dominated [n = 25], and exchange-dominated [n = 6]), as defined by Brock et al. (2007) (Fig. 1).
Water and surface sediment samples were collected with the aid of a helicopter in September 2002. Water samples were collected from the central or deepest part of 41 delta lakes, while three river samples were collected at mid-channel (in all cases from 10 cm below the water surface). In situ limnological measurements (water depth, temperature, pH and conductivity) were recorded at each sampling site and water samples were collected for standard chlorophyll and chemical analyses (dissolved phosphorus (dP), total phosphorus (TP), total nitrogen (TKN), nitrate + nitrite (NO −3 + NO −2 ) dissolved organic carbon (DOC), colour, major ions (Ca2+, Mg2+, K+, Na+, Cl−, SO 2−4 ), dissolved silica (SiO2), and alkalinity). Large particles were removed prior to analyses of water chemistry and chlorophyll by passing water through a 650 μm screen. The water was then filtered and the particles retained by a 0.7 μm filter were frozen and analyzed for chlorophyll-a (chl-a) using standard spectrophotometric methods (Jeffrey et al. 1997). Water samples (1 l) for chemical analyses were kept at 4°C and sent to the Water Chemistry Laboratory, University of Montreal, for analysis using methods described in Environment Canada (1996) within 3–5 days of collection.
Surface sediment samples (0–1 cm) were collected at the time of water sampling from the same 41 delta lakes at the central or deepest part of each basin using a mini-Glew gravity corer (Glew 1991), and were stored in Whirl-pak® bags at 4°C until processed for subsequent analyses. Microscope slides for diatom analysis were prepared from wet sediment samples by acid digestion following standard methods (Hall and Smol 1996). For each sample, a minimum of 400 diatom valves were identified and enumerated along transects using a Zeiss Axioskop II Plus compound microscope with differential interference contrast optics (1000× magnification, numerical aperture = 1.30). Diatom taxonomy followed Krammer and Lange-Bertalot (1986–1991). Diatom data were expressed as taxon relative abundances (%) of the total sum of diatom valves in each sample.
Numerical methods
Prior to numerical analyses, all limnological variables were tested for non-normal distribution by constructing histograms using the computer program SPSS version 12.0, and deviation from normality was assessed visually. To correct for skewed distributions, ln (x + b) transformation (where b is 0.5 × the minimum non-zero value) was applied to NO −3 + NO −2 , TP, colour, SiO2, K and chl-a. Diatom species that were encountered at <3 sites and with a maximum abundance <1% were classified as rare taxa and were omitted from numerical analyses to avoid the influence of taxa whose distributions were poorly characterized by the samples we analyzed. Diatom percent abundance data were square-root transformed to reduce the influence of dominant taxa.
Principal Components Analysis (PCA), an indirect gradient, multivariate ordination technique, was used to assess the main limnological differences among hydrological lake categories. Rivers were included passively and lakes were coded in ordination plots based on Brock et al.’s (2007) hydrological categories (flood-, evaporation- and exchange-dominated). Analysis of variance (ANOVA) was used to assess limnological differences (P ≤ 0.05) among river sites (including Great Slave Lake) and the three hydrological lake categories. ANOVA analyses included post hoc tests (P ≤ 0.05) based on Tukey’s HSD test statistic using SPSS version 12.0.
Detrended Correspondence Analysis (DCA) was performed on the diatom data to quantify the gradient length (in SD units) of the first axis, which was used to determine if linear or unimodal based ordination methods were most appropriate for analyzing the diatom data. All gradient lengths were <2 standard deviation units, therefore, linear ordination techniques (PCA and Redundancy Analysis (RDA)) were employed (Birks 1995). PCA was used to assess distributions of surface sediment diatom assemblages among the hydrological lake categories. Direct gradient ordination by RDA was used to identify relationships between the surface sediment diatom assemblages and the limnological gradients among the lakes. RDA was performed using forward selection (and Monte Carlo tests with 999 permutations) to determine a subset of variables that explained significant (P ≤ 0.05) and independent amounts of variation in the diatom data (ter Braak and Šmilauer 2002). Sample scores (or surface sediment diatom assemblages) in the ordination diagrams were coded a priori according to the three hydrological lake categories to visualize variation in limnological conditions and diatom assemblage composition among the categories. All ordinations were performed using CANOCO version 4.5 (ter Braak and Šmilauer 2002).
Analysis of Similarities (ANOSIM), a non-parametric multivariate test that is analogous to a one-factor ANOVA (Clarke and Warwick 2001), was used to determine if the composition of surface sediment diatom assemblages differed among the three hydrological lake categories. The sample statistic (Global R value) reflects the observed difference among groups of samples contrasted with the difference among replicates within each group of samples. A Global R value of zero indicates that the similarity between and within groups of samples is the same on average. A value of one, on the other hand, indicates that all the replicates within a group of samples are more similar to each other than any replicates from different groups of samples (Clarke and Warwick 2006). Within- and across-group rank Bray–Curtis dissimilarities were computed (5000 permutations) and the distribution of these values was compared to the initial rank dissimilarity and reported as the Global R value (Clarke and Warwick 2006). For example, if a Global R value was significantly greater than zero (P ≤ 0.05), this would indicate that differences in diatom assemblages between hydrological categories are greater than variability within the hydrological categories.
Similarity Percentage tests (SIMPER; Clarke and Warwick 2006) were performed to identify specific diatom taxa that accounted for the greatest observed differences between hydrological categories. Taxa contributing >2% of the average Bray–Curtis similarity within a hydrological category were considered ‘indicator’ taxa that are most representative of that hydrological category. ANOSIM and SIMPER analyses were performed using the program PRIMER version 6.1.5 (Clarke and Warwick 2006). In a final step, Canonical Variates Analysis (CVA), a form of multiple discriminant analysis (Birks et al. 1975), was used to assess if the ‘indicator’ taxa identified by SIMPER analyses significantly discriminate (P ≤ 0.05) the three Slave River Delta hydrological lake categories. The CVA was performed by using the SIMPER-identified diatom taxa as explanatory variables and hydrological lake categories (coded as dummy variables for each site) as the response variables. These data were used in a Canonical Correspondence Analysis with Hill’s scaling focused on inter-species distances and run using Monte Carlo tests with 999 permutations to assess the significance of the CVA axes.
Results
Physical and chemical limnological characteristics
Lakes in the Slave River Delta are shallow (mean maximum depth = 126 cm; range: 30–280 cm), high in nutrients (mean TP = 48.85 μg l−1; range: 15.3–175.3 μg l−1 and mean TKN = 1299.64 μg l−1; range: 308.3–2637.7 μg l−1), alkaline (mean pH = 7.99; range: 7.4–8.4), and contain relatively high concentrations of major ions (Table 1). One exception is SD30 (Ring Lake), a relatively deep (Z max = 10 m) oxbow lake that is continuously connected to the Slave River by a single channel (Fig. 1). Great Slave Lake, the Slave River and its distributaries have high concentrations of TP (mean = 111.79 μg l−1; range: 30.9–285.0 μg l−1) and SO4 (mean = 6.01 mg l−1; range: 5.6–7.0 mg l−1), while most other chemical parameters are relatively low compared to the lakes (Table 1). Although TP concentrations are high at the river sites, dP concentrations are low (mean = 5.06 μg l−1; range: 3.0–8.1 μg l−1), indicating that most of the phosphorus is in particulate forms. Since chl-a concentrations are low (mean = 2.61 μg l−1; range: 2.37–2.84 μg l−1), most of the phosphorus is contained in the suspended mineral sediments rather than phytoplankton and other suspended biota.
Principal Components Analysis of the water chemistry data showed that lakes differed mainly along gradients of concentrations of nutrients (N, P, DOC), ions, alkalinity and chl-a (Fig. 2). Eigenvalues for the first and second PCA axes were 0.787 and 0.213 respectively, and explained all of the variation in the data set. TKN, TP, dP, DOC, Colour, chl-a, K, Mg, alkalinity and conductivity were all strongly and negatively correlated to axis 1, while pH and Na were closely associated with axis 2. Concentrations of SiO2, Ca, Cl, SO4 and NO3 + NO2 contributed nearly equally to the variation along the first and second PCA axes. The main gradient of variation among sites (axis 1) separates lakes that have no river influence (evaporation-dominated lakes) from those that receive periodic inputs of water from the Slave River or Great Slave Lake (flood- and exchange-dominated lakes). Evaporation-dominated lakes tend to have high alkalinity and high concentrations of nutrient and ions. Exceptions are lakes SD8 and SD32, which were classified as evaporation-dominated (Brock et al. 2007) but had water chemistry more typical of flood-dominated lakes (Fig. 2). The PCA plot also shows that evaporation-dominated lakes have the broadest range of limnological conditions. In contrast, flood-dominated lakes cluster more tightly along the main PCA axis (axis 1), characterized by high concentrations of Cl and SO4 and low concentrations of nutrients, whereas exchange-dominated lakes are distributed along axis 1 spanning a broader range of nutrient and ion concentrations. The exchange-dominated lakes SD30, SD39 and SD41 plot close to the rivers indicating their water chemistries are similar to that of river water, characterized by low concentrations of nutrients, ions, chl-a and alkalinity. However, other exchange-dominated lakes possess quite different limnological properties. Lakes SD28 and SD17 plot near the evaporation-dominated lakes due to moderately high nutrient and ion concentrations and alkalinity, while SD10 is chemically similar to flood-dominated lakes, due to relatively high pH and concentrations of Cl and SO4 (Fig. 2).

Principal components analysis (PCA) ordination diagram illustrating the relative position of Slave River Delta lakes (n = 41) with respect to three hydrological lake categories and 17 physical/chemical variables. River sites and Great Slave Lake are plotted passively
Concentrations of several nutrients (TKN, dP, DOC) and major ions (K, SO4) differ significantly (P ≤ 0.05) among flood-, exchange- and evaporation-dominated lake categories (Fig. 3), while some variables (e.g. pH and concentrations of NO3 + NO2, Cl and chl-a [not shown]) did not vary significantly. River sites and Great Slave Lake had significantly lower alkalinity, conductivity and K concentration compared to the three hydrological categories of deltaic lakes. Similar, but non-significant tendencies occurred for concentrations of TKN, dP, DOC, Na and Mg, whereas TP, SiO2 and SO4 were higher in rivers. Generally, concentrations of nutrients and the majority of ions are higher in evaporation-dominated lakes relative to flood- and exchange-dominated lakes. Exchange-dominated lakes have the lowest values and are most similar to river sites compared to lakes in the other hydrological categories (Figs. 2, 3).

Box plots of selected chemical variables for river sites (includes Slave River distributaries and Great Slave Lake) (R; n = 4), flood-dominated (F; n = 10), exchange-dominated (Ex; n = 6), and evaporation-dominated (Ev; n = 25) lakes. Outliers (solid dots) are samples outside the 5th and 95th percentiles. Small case letters (a, b, c, d) are used to identify limnological variables with statistically significant differences (P < 0.05) in mean values between the hydrological categories based on ANOVA tests
Relationships between surface-sediment diatoms and hydroecological conditions
A total of 88 diatom taxa from 23 genera were found in the surface sediment samples collected from the 41 study lakes (Appendix). Most of the diatom taxa encountered typically occupy epiphytic and benthic habitats and have been reported from spatial surveys of shallow lakes and ponds in the Mackenzie and Peace-Athabasca deltas (Hay et al. 2000; Hall et al. 2004). Planktonic diatoms (e.g. Cyclostephanos spp., Stephanodiscus minutulus (Kützing) Cleve & Möller, S. parvus Stoermer & Håkansson) were found at low to modest relative abundance in most flood- and exchange-dominated lakes, but were largely absent from lakes in the evaporation-dominated hydrological category.
Ordination of the diatom data by PCA showed that composition of surface sediment assemblages differs among the hydrological lake categories (Fig. 4). Eigenvalues for the first and second axes were 0.167 and 0.118 respectively, and explained 28.5% of the variation in the data set. The main gradient of variation separated diatom assemblages in the river-influenced lakes (flood- and exchange-dominated; positioned to the left in Fig. 4) from evaporation-dominated lakes (positioned to the right). Surface sediment diatom assemblages in river-influenced lakes (flood- and exchange-dominated categories) included taxa that occupy a wide diversity of habitats (e.g. river-borne planktonic taxa (Cyclostephanos spp. and Stephanodiscus spp.), benthic taxa (Gyrosigma attenuatum (Kützing) Rabenhorst and Rhopalodia gibba (Ehrenberg) O. Müller) and a variety of common epiphytes (Navicula and Nitzschia taxa)). Composition of diatom assemblages overlaps considerably between flood- and exchange-dominated lakes. Surface sediment diatom assemblages of evaporation-dominated lakes generally cluster as a discrete group with positive axis 1 scores, and are almost entirely composed of common epiphytic taxa such as Navicula minima Grunow, Nitzschia amphibia Grunow, and taxa belonging to Achnanthes, Cocconeis and Gomphonema. One exception to this pattern is lake SD32, which has a diatom assemblage that is more similar to those of flood-dominated lakes (i.e. with modest abundance of Cyclostephanos PAD sp. 2, Gyrosigma attenuatum and Rhopalodia gibba).

Principal components analysis (PCA) of sedimentary diatom taxa (n = 88) from lakes in the Slave River Delta (n = 41). Site scores are located in the left panel and diatom taxa scores are located in the right panel. Numbers correspond with taxon names provided in Appendix
Three lakes (SD7, SD13 and SD24) are conspicuous in the PCA because they are positioned high on axis 2, clearly separated from the other sites (Fig. 4). Although these lakes have quite different water chemistry (i.e. they are positioned far apart in the PCA performed on water chemistry variables; see Fig. 2), their diatom assemblages are all dominated by small benthic Fragilaria taxa (e.g. F. pinnata Ehrenberg, F. pinnata var. intercedens Ehrenberg, F. construens (Ehrenberg) Grunow, F. construens var. venter (Ehrenberg) Grunow and F. construens f. binodis (Ehrenberg) Grunow).
Ordination by RDA identified five variables (TKN, TP, K, Mg and SiO2) that explained significant and independent amounts of variation in the surface sediment diatom assemblages among sites (Fig. 5). The eigenvalues of the first two RDA axes (λ1 = 0.129, λ2 = 0.061) were both significant (P ≤ 0.05) and captured 46.2% and 21.8% of the variation in the species-environment relationships, respectively. Concentrations of TKN, TP and K are highly correlated with the first RDA axis (intraset correlations = 0.781, 0.735 and 0.700, respectively) and SiO2 shows the strongest correlation with the second axis (intraset correlation = 0.575), while Mg contributes nearly equally to the variation along both the first and second axes (intraset correlations = 0.252 and 0.275, respectively). The first RDA axis mainly separated diatom assemblages typical of evaporation-dominated lakes (higher nutrient and ion concentrations and dominated by Navicula minima, Nitzschia amphibia and Achnanthes taxa) from assemblages typical of river-influenced (flood- and exchange-dominated) lakes (lower concentrations of nutrients and ions that were dominated by Gyrosigma attenuatum, Rhopalodia gibba, and taxa of the genera Navicula, Nitzschia, Cyclostephanos and Stephanodiscus). To a lesser degree, the second RDA axis separated the diatom assemblages of flood-dominated lakes (positioned in the lower left quadrant, associated with relatively low concentrations of SiO2 and relatively abundant Navicula libonensis Schoeman and Cocconeis taxa) from exchange-dominated lakes (positioned in the upper left quadrant, associated with higher concentrations of SiO2 and abundant Eunotia bilunaris (Ehrenberg) Mills, Fragilaria capucina var. gracilis (Oestrup) Hustedt and Cymbella taxa).

Redundancy analysis (RDA) of sedimentary diatom taxa (n = 88) and forward-selected environmental variables from Slave River Delta lakes (n = 41). Site scores are located in the left panel and diatom taxa scores are located in the right panel. Numbers correspond with taxon names provided in Appendix
In terms of diatom assemblage composition (Fig. 4) and relationships between water chemistry and diatom assemblage composition (Fig. 5), lake SD32 shares more affinity with lakes in the flood-dominated hydrological category than the evaporation-dominated lakes. This appears to be a singular inconsistency in relating the limno-ecological data to the hydrological classification of lakes developed by Brock et al. (2007) from analysis of stable isotopes in point-in-time lake water samples. Consequently, lake SD32 was placed in the flood-dominated hydrological category for subsequent ecological analyses.
Composition of surface sediment diatom assemblages differed significantly among all three hydrological lake categories of the Slave River Delta, as assessed by ANOSIM tests (Table 2). Analysis by SIMPER identified a number of unique ‘indicator’ diatom taxa that can discriminate among hydrological lake categories (i.e. the taxa that accounted for >2% of the average Bray–Curtis similarity within groups; Fig. 6a). Diatom taxa identified as indicators of a particular hydrological lake category account for 98% of the similarity within that category although several diatom taxa, mainly those occupying epiphytic habitats, were identified as indicators of more than one hydrological lake category (Fig. 6b). Navicula minima and Nitzschia amphibia were identified as unique indicators of evaporation-dominated lakes, whereas Eunotia bilunaris (Ehrenberg) Mills, Fragilaria capucina var. gracilis, Nitzschia perminuta (Grunow) M. Peragallo, Cyclostephanos cf. tholiformis Stoermer, Håkansson & Theriot and Stephanodiscus minutulus were uniquely associated with exchange-dominated lakes. Navicula libonensis, Gyrosigma attenuatum, Rhopalodia gibba and Cyclostephanos PAD sp. 2 were identified as unique indicators of flood-dominated lakes. Most of the ‘indicator’ diatom taxa occupy epiphytic and benthic habitats, with the exception of the centric diatoms Cyclostephanos cf. tholiformis, Stephanodiscus minutulus (exchange-dominated) and Cyclostephanos PAD sp. 2 (flood-dominated), which originate from the Slave River and Great Slave Lake (M.A. Sokal, personal observation).

Relative abundance (%) profiles of sedimentary diatom taxa contributing >2% of the Bray–Curtis similarity in the assemblages within a hydrological category. Panel (a) includes diatom taxa meeting this criterion for only one hydrological category, whereas panel (b) includes taxa meeting the criterion for more than one hydrological category
Canonical Variates Analysis provided evidence that the ‘indicator’ taxa identified by the SIMPER analysis account for significant (p ≤ 0.05) discrimination among the hydrological lake categories. Eigenvalues of the first two CVA axes are 0.876 and 0.832, respectively, and explain 85.3% of the variance among hydrological lake categories. Thus, the threshold criteria used in our SIMPER analyses (i.e., diatom taxa contributing >2% of the average Bray–Curtis similarity within groups) successfully identified ‘indicator’ taxa that distinguish hydroecological conditions of lakes within the Slave River Delta.
Discussion
The hydrological settings of lakes in the Slave River Delta are associated with distinctly different limnological conditions and composition of diatom assemblages in surface sediments, as summarized in Table 3. Differences are most pronounced between lakes that are influenced by the Slave River or Great Slave Lake (flood- and exchange-dominated lakes) and lakes that are removed from the influence of the river (evaporation-dominated lakes). This is not surprising given the fact that a strong influence of river flood water on the limnology and ecology of floodplain lakes has been observed in both the Mackenzie Delta (Hay et al. 1997, 2000; Squires and Lesack 2002; Squires et al. 2002) and the Peace-Athabasca Delta (Hall et al. 2004; Wolfe et al. 2007b). Evaporation-dominated lakes possess water balances controlled by precipitation, snowmelt runoff and evaporation (Table 3). As a result, they are limnologically most dissimilar to the rivers and are highly dominated by epiphytic diatoms. In contrast, flood- and exchange-dominated lakes receive inputs of river or Great Slave Lake water which lower their concentrations of nutrients, chl-a, the majority of ions and alkalinity, and their diatom assemblages include a variety of epiphytic diatoms, but most notably include unique benthic and planktonic taxa (Table 3).
A strong relationship exists between the hydrological lake category and water chemistry, indicating the hydrological setting strongly regulates limnological properties (Fig. 2; Table 3). The diatom assemblages preserved in the surface sediments also show strong and statistically significant differences in hydro-limnological conditions among all three hydrological lake categories (Figs. 4 and 5). Thus, the isotope-based classification by Brock et al. (2007) captures major limnological and hydroecological differences among lakes of the Slave River Delta. The only consistent exception is lake SD32 which had a diatom composition typical of lakes with flood-dominated hydrology, even though it was categorized by Brock et al (2007) as an evaporation-dominated lake. This suggests that the isotope-based criteria used to classify this lake were strongly influenced by short-term effects of summer evaporation as this lake did not flood in the spring of 2003. However, diatoms incorporated in surface sediments indicate that over longer timescales this lake behaves more like flood-dominated lakes. All of the other evaporation-dominated lakes tend to have the highest concentrations of nutrients and several ions, which can be best explained by increased evaporative enrichment of ions and internal recycling of nutrients in the absence of river flooding (e.g. Hay et al. 1997). The diatom assemblages in evaporation-dominated lakes have high relative abundances of common epiphytes, a feature that is consistent with observed clear water conditions of low phytoplankton abundance and extensive macrophyte coverage throughout all or most of these lake basins. These diatom assemblages are associated with high concentrations of nutrients (TKN and TP) and ions (K and Mg), similar to findings by Hall et al. (2004) for closed-drainage lakes in the Peace-Athabasca Delta which are analogous to evaporation-dominated lakes in the Slave River Delta. Hay et al. (2000) also identified that similar epiphytic diatom assemblages typified the analogous high-closure lakes in the Mackenzie Delta that support dense macrophyte beds in the absence of strong flood influence.
In flood-dominated lakes, flooding from rivers is typically restricted to the spring melt season, but exchange-dominated lakes periodically re-connect to the river or are inundated with water from Great Slave Lake during seiche events throughout the open-water season (Brock et al. 2007). Consequently, limnological conditions of exchange-dominated lakes tend to be more variable compared to lakes of the other hydrological categories. Additionally, local geographic factors (e.g. distance to river and Great Slave Lake, sill elevation, presence of channel connections) likely influence the frequency and magnitude of flooding of lakes in the exchange-dominated category. Thus, it is not surprising that lakes in the exchange-dominated category encompass a broad range of limnological conditions that overlaps to some extent with lakes of hydrological categories at the two extremes (evaporation- and flood-dominated). For example, lakes SD30, SD39 and SD41 have limnological characteristics similar to river sites, suggesting that at the time of sampling, the Slave River was flowing into SD30 and water from Great Slave Lake was entering SD39 and SD41 at the outer portion of the delta. Contrary to this, SD17 and SD28 (both of which have channel connections to the Slave River) were limnologically more similar to evaporation-dominated lakes at the time of sampling, suggesting their channels were acting as outflows or disconnected entirely. Diatom assemblages in exchange-dominated lakes are associated with high levels of SiO2 which is consistent with prolonged river connection and high concentrations of SiO2 in river water. Although flood-dominated lakes receive SiO2-rich flood waters in the spring, the effects appear to be short-lived as is suggested by the separation of flood- from exchange-dominated lakes along a SiO2 gradient (Fig. 5).
Increased river influence often results in decreased light transparency, macrophyte coverage and percent abundance of epiphytic diatoms in open-drainage lakes of the Peace-Athabasca Delta and no-closure lakes of the Mackenzie Delta, which are continuously flooded by rivers and thus highly turbid (Hay et al. 2000; Squires et al. 2002; Hall et al. 2004). However, this situation does not appear to be the case in the Slave River Delta, and such lakes do not exist or at least were not sampled in our study. Flood- and exchange-dominated lakes remain dominated by macrophytes and epiphytic diatom taxa (as are evaporation-dominated basins), in spite of high minerogenic turbidity, and are more similar to restricted-drainage lakes in the Peace-Athabasca Delta and low-closure lakes in the Mackenzie Delta which receive periodic flooding (Hay 1997; Hall et al. 2004).
Diatom assemblages in most lakes in this study are dominated by epiphytic taxa, reflecting the large habitat space (macrophyte surface area) available in these shallow deltaic ponds that support prolific macrophyte growth. Many of the epiphytic diatom taxa were distributed broadly among lakes of more than one hydrological category, and thus these taxa are likely unable to distinguish changes in hydrological status of lakes in the Slave River Delta. Importantly, however, we have identified specific ‘indicator’ taxa that can discriminate the three hydrological lake categories (Table 3). The epiphytic diatoms Navicula minima and Nitzschia amphibia are associated with evaporation-dominated lakes and are almost non-existent in flood- or exchange-dominated lakes. Both of these species are relatively abundant in high-closure lakes of the Mackenzie Delta (Hay et al. 2000) and are indicators of closed-basin lakes in the Peace-Athabasca Delta (Hall et al. 2004). Diatoms indicative of flood- and exchange-dominated lakes span a much wider range of habitat types including epiphytic, planktonic, tychoplanktonic and motile benthic taxa (Fig. 6). The planktonic ‘indicator’ taxa Cyclostephanos cf. tholiformis, Stephanodiscus minutulus (exchange-dominated) and Cyclostephanos PAD sp. 2 (flood-dominated) are important indicators of connectivity to rivers and Great Slave Lake, and similarly they have been associated with high river connectivity in the Peace-Athabasca Delta (Hall et al. 2004). Curiously, Cyclostephanos cf. tholiformis is found in lakes with exchange-dominated hydrology but not in lakes with flood-dominated hydrology, while the opposite pattern occurs for Cyclostephanos PAD sp. 2, but it is difficult to identify a mechanism responsible for this apparent phenomenon because both taxa are commonly found in Slave River plankton samples.
Conclusions
We demonstrate that variations in basin hydrology correspond with differences in limnological conditions and composition of diatom assemblages in the surface sediments of lakes in the Slave River Delta. The distinct composition of water chemistry and diatom assemblages among hydrological categories indicates that the lake classification scheme of Brock et al. (2007) based on stable isotope results from the open-water season of 2003 also captures major limnological and ecological differences (Table 3). In the absence of river flooding, evaporation-dominated lakes have the highest concentrations of several ions and nutrients and these clear water, macrophyte dominant systems have high relative abundances of common epiphytic diatoms such as Navicula minima and Nitzschia amphibia. In contrast, flood-dominated lakes have relatively low concentrations of most ions and nutrients, while exchange-dominated lakes have the lowest average concentrations of most ions and nutrients, but tend to be more variable as a result of varying influence by the Slave River and Great Slave Lake. Sedimentary diatom taxa indicative of flood- and exchange-dominated lakes span a wider range of habitat types when compared to evaporation-dominated lakes, and include the planktonic taxa Cyclostephanos cf. tholiformis, Stephanodiscus minutulus (exchange-dominated) and Cyclostephanos PAD sp. 2 (flood-dominated) which are important indicators of river connectivity in the Slave River Delta.
Our approach has integrated the use of water isotope tracers and chemistry to characterize present hydrolimnological conditions, and demonstrated the potential of sedimentary diatom assemblages as bioindicators of hydrological changes and as proxy indicators of past hydroecological conditions in the Slave River Delta. Knowledge gained from this formative study is being used in biomonitoring and paleolimnological investigations of the Slave River Delta to further understanding of the complex interactions among hydrology, limnology, and aquatic ecology in this freshwater ecosystem. In particular, ongoing multi-proxy (stable isotopes, diatoms, plant macrofossils) paleolimnological studies at several lakes situated throughout the delta will use the information provided by this study to assess the relative roles of river discharge, frequency of ice-jam floods, climatic variability and geomorphic change on the Slave River Delta ecosystem. This is necessary for anticipating responses to climate change and human modifications of upstream river systems, including effects of river regulation by the WAC Bennett Dam and consumptive water use by the Athabasca Oil Sands operations.
References
Birks HJB (1995) Quantitative paleoenvironmental reconstructions. In: Maddy D and Brew JS (eds) Statistical modeling of Quaternary science data. Quaternary Research Association Technical Guide No. 5, Cambridge, pp 161–254
Birks HJB, Webb III T, Berti AA (1975) Numerical analysis of pollen samples from central Canada: a comparison of methods. Rev Palaeobot Palynol 20:133–169
Brock BE, Wolfe BB and Edwards TWD (2007) Characterizing the hydrology of shallow floodplain lakes in the Slave River Delta, NWT, using water isotope tracers. Arct Antarct Alp Res (in press)
Clarke KR, Warwick RM (2001) Changes in marine communities: an approach to statistical analysis and interpretation (2nd edn). Plymouth Marine Laboratory, Plymouth, UK, 144 pp
English MC, Hill RB, Stone MA, Ormson R (1997) Geomorphological and botanical change on the outer Slave River Delta, NWT, before and after impoundment of the Peace River. Hydrol Proc 11:1707–1724
Environment Canada (1996) Manual of analytical methods. The national laboratory of environmental testing, Canada Centre for Inland Waters, Burlington
Glew JR (1991) A miniature gravity corer for recovering short sediment cores. J Paleolimnol 5:285–287
Hall RI, Smol JP (1996) Paleolimnological assessment of long-term water-quality changes in south-central Ontario lakes affected by cottage development and acidification. Can J Fish Aquat Sci 53:1–17
Hall RI, Wolfe BB, Edwards TWD, Karst-Riddoch TL, Vardy SR, McGowan S, Sjunneskog C, Paterson A, Last WM, English M, Sylvestre F, Leavitt PR, Warner BG, Boots B, Palmini R, Clogg-Wright KP, Sokal MA, Falcone M, van Driel P and Asada T (2004) A multi-century flood, climatic, and ecological history of the Peace-Athabasca Delta, northern Alberta, Canada. Final report. Published by BC Hydro, pp 163 + Appendices
Hay MB, Smol JP, Pipke KJ, Lesack LWF (1997) A diatom-based paleohydrological model for the Mackenzie Delta, Northwest Territories, Canada. Arct Alp Res 29:430–444
Hay MB, Michelutti N, Smol JP (2000) Ecological patterns of diatom assemblages from Mackenzie Delta lakes, Northwest Territories, Canada. Can J Bot 78:19–33
Hill MO (1973) Diversity and evenness: a unifying notation and its consequences. Ecology 54:427–432
Jeffrey SW, Mantoura RFC, Wright SW (1997) Phytoplankton Pigments in Oceanography: guidelines to Modern Methods. UNESCO Publishing, Paris, pp 661
Junk WJ (2005) Flood pulsing and the linkages between terrestrial, aquatic and wetland systems. Verh Internat Verein Limnol 29:11–38
Krammer K, Lange-Beralot H (1986–1991) Bacillariophyceae. In: Ettl H, Gerloff J, Heynig H, Mollenhauer D (eds) Sϋβwasserflora von mitteleuropa band 2/1–4. Gustav Fischer Verlag, Stuttgart
Lesack LFW, Marsh P, Hecky RE (1998) Spatial and temporal dynamics of major solute chemistry among Mackenzie Delta lakes. Limnol Oceanogr 43:1530–1543
Lewis Jr WM, Hamilton SK, Lasi MA, Rodriguez M, Saunders III JF (2000) Ecological determinism on the Orinoco floodplain. BioScience 50:681–692
Michelutti N, Hay MB, Lesack LFW, Smol JP (2001) Diatom changes in lake sediments from the Mackenzie Delta, N.W.T., Canada: paleohydrological applications. Arc Antarc Alp Res 33:1–12
Milburn D, Prowse TD (1998) Sediment bound contaminants in a remote northern delta. Nord Hydrol 29:397–414
Nilsson C, Reidy CA, Dynesius M, Revenga C (2005) Fragmentation and flow regulation of the world’s large river systems. Science 308:405–408
Overpeck J, Hughen K, Hardy D, Bradley R, Case R, Douglas M, Finney B, Gajewski K, Jacoby G, Jennings A, Lamoureux S, Lasca A, MacDonald G, Moore J, Retelle M, Smith S, Wolfe A, Zielinski G (1997) Arctic environmental change of the last four centuries. Science 278:1251–1256
Prowse TD, Conly FM (2001) Multiple-hydrologic stressors of a northern delta ecosystem. J Aquat Ecosyst Stress Recovery 8:17–26
Prowse TD, Conly FM (2002) A review of hydroecological results of the northern river basins study, Canada. Part 2. Peace-Athabasca Delta. Riv Res Appl 18:447–460
Prowse TD, Conly FM, Church M, English MC (2002) A review of hydrological results of the northern river basins study, Canada. Part 1. Peace and Slave River. Riv Res Appl 18:429–446
Sakaguchi A, Yamomoto M, Sasaki K, Kashiwaya K (2006) Uranium and thorium isotope distribution in an offshore bottom sediment core of the Selenga Delta, Lake Baikal, Siberia. J Paleolimnol 35:807–818
Schindler DW, Donahue WF (2006) An impending water crisis in Canada’s western prairie provinces. Proc Natl Acad Sci USA 109:7210–7216
Schindler DW, Smol JP (2006) Cumulative effects of climate warming and other human activities on freshwaters of arctic and subarctic North America. Ambio 35:160–168
Serreze MC, Walsh JE, Chapin III FS, Osterkamp T, Dyurgerov M, Romanovsky V, Oechel WC, Morison J, Zhang T, Barry RG (2000) Observational evidence of recent change in the northern high-latitude environment. Clim Change 46:159–207
Spears BM, Lesack LFW (2006) Bacterioplankton production, abundance, and nutrient limitation among lakes of the Mackenzie Delta (western Canadian arctic). Can J Fish Aquat Sci 63:845–857
Squires MM, Lesack LFW (2001) Benthic algal response to pulsed versus distributed inputs of sediments and nutrients in a Mackenzie Delta lake. J North Am Benthol Soc 20:369–384
Squires MM, Lesack LFW (2002) Water transparaency and nutrients as controls on phytoplankton along a flood-frequency gradient among lakes of the Mackenzie Delta, western Canadian Arctic. Can J Fish Aquat Sci 59:1339–1349
Squires MM, Lesack LFW (2003) The relation between sediment nutrient content and macrophyte biomass and community structure along a water transparency gradient among lakes of the Mackenie Delta. Can J Fish Aquat Sci 60:333–343
Squires MM, Lesack LFW, Huebert D (2002) The influence of water transparency on the distribution and abundance of macrophytes among lakes of the Mackenzie Delta, Western Canadian Arctic. Freshwater Biol 47:123–135
ter Braak CJF, Šmilauer P (2002) CANOCO Reference manual and CanoDraw for Windows user’s guide: software for Canonical Community Ordination (Version 4.5). Microcomputer Power, Ithaca, New York, pp 500
Vanderburgh S, Smith DG (1988) Slave River Delta: geomorphology, sedimentology, and Holocene reconstruction. Can J Earth Sci 25:1990–2004
Wolfe BB, Karst-Riddoch TL, Vardy SR, Falcone MD, Hall RI, Edwards TWD (2005) Impacts of climate and river flooding on the hydro-ecology of a floodplain basin, Peace-Athabasca Delta, Canada since A.D. 1700. Quat Res 64:147–162
Wolfe BB, Hall RI, Last WM, Edwards TWD, English MC, Karst-Riddoch TL, Paterson AM, Palmini R (2006) Reconstruction of multi-century flood histories from oxbow lake sediments, Peace-Athabasca Delta, Canada. Hydrol Proc 20:4131–4153
Wolfe BB, Armitage D, Wesche S, Brock BE, Sokal MA, Clogg-Wright KP, Mongeon CL, Adam ME, Hall RI, Edwards TWD (2007a) From isotopes to TK interviews: towards interdisciplinary research in Fort Resolution and the Slave River Delta, NWT. Arctic 60:75–87
Wolfe BB, Karst-Riddoch TL, Hall RI, Edwards TWD, English MC, Palmini R, McGowan S, Leavitt PR, Vardy SR (2007b) Classification of hydrological regimes of northern floodplain basins (Peace-Athabasca Delta, Canada) from analysis of stable isotopes (δ18O, δ2H) and water chemistry. Hydrol Proc 21:151–168
Zalat A, Servant Vildary S (2007) Environmental change in Northern Egyptian delta lakes during the late Holocene, based on diatom analysis. J Paleolimnol 37:273–299
Acknowledgements
We thank the Deninu Kue First Nation, Deninu School (especially the Principal, Moh Odeen), K.P. Clogg-Wright, T.W.D. Edwards, M.C. English, Mr. G. Lafferty of Fort Resolution, and M. Morin of Wood Buffalo Helicopters for field and laboratory assistance and logistical services. Water chemistry analyses were conducted by Richard Carignan and his staff at the Water Chemistry Laboratory, University of Montreal. Funding for this study was provided by a NSERC Northern Research Chair and a Premier’s Research Excellence Award to B.B. Wolfe, a NSERC Discovery grant to R.I. Hall, Northern Scientific Training Program grant and Ontario Graduate Scholarships to M.A. Sokal, and BC Hydro.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sokal, M.A., Hall, R.I. & Wolfe, B.B. Relationships between hydrological and limnological conditions in lakes of the Slave River Delta (NWT, Canada) and quantification of their roles on sedimentary diatom assemblages. J Paleolimnol 39, 533–550 (2008). https://doi.org/10.1007/s10933-007-9128-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-007-9128-8