
Abstract
The presence of osteoderms in the skin of some extinct sloths and in cingulates (armadillos, pampatheres, and glyptodonts) has often been considered a pleisomorphic character of the Xenarthra. While osteoderms are known from the earliest cingulates, they are absent in most sloths including the two extant taxa and only appear late in their fossil record. Osteoderms are currently only reported from five genera of mylodonts and two megatheres, out of the over 100 currently recognized genera of sloths. Consequently, rather than a plesiomorphic character of the Xenarthra, which has been secondarily lost in sloths, it is more likely that osteoderms in sloths are the result of parallel evolution to the cingulates that independently evolved in one, possibly two different sloth clades.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Within placental mammals, ossifications in the skin, osteoderms, are only present within members of the Xenarthra. Historically, reviews of the evolution of the Xenarthra have considered the presence of dermal ossicles or osteoderms a plesiomorphic character of the order (Romer 1966). While this distinctive anatomical character clearly defines one of the subgroups, the Cingulata, its presence in the other major clade, the Pilosa is restricted to only a few genera. I provide a general review of the presence of osteoderms within the sloths and the implications for this limited taxonomic distribution. Reported sloth osteoderms from the literature along with personal examination of unpublished records of specimens in museum collections are used to examine the relative robustness of the fossil record in documenting the presence of osteoderms in fossil sloths. The recent blossoming of research in bone histology of fossil specimens including studies of the osteoderms of both cingulates and pilosans has contributed valuable new information on the anatomical differences in the osteoderms of the two groups. The recent literature on xenarthran osteoderm histology is relatively extensive and only selected references are utilized here to provide a general overview of the major anatomical differences in the osteoderms of cingulates and pilosans. For more specific details the reader is urged to utilize the primary sources. Data sharing is not applicable to this article as no datasets were generated or analyzed during the study.
Geological History of Osteoderms in Xenarthrans
Dermal ossifications or osteoderms are common in many groups of tetrapods, including some amphibians (dissorophid temnospondyls), placodonts, chelonia, phytosaurs, lepidosaurs, crocodylians, and ornithischian (stegosaurs and ankylosaurs) and sauropodomorph dinosaurs. One major group in which they do not occur with any frequency is the Synapsida, with the exception of two varanopsids from the Late Permian of South Africa, Heleosaurus scholtzi (Reisz and Modesto 2007) and Elliotsmithia longiceps (Dilkes and Reisz 1996). The arrangement of the osteoderms in Heleosaurus is rectilinear and arranged in longitudinal and transverse rows, so does not resemble the hexagonal pattern seen in the carapace of glyptodonts and scapular and buckler shields of pampatheres and the Dasypodinae, or in the Euphractinae, where the morphology is variable or the mosaic pattern formed by sloth osteoderms. Based on the current fossil record there is no evidence of osteoderms in the therapsid lineage leading to mammals. While xenarthrans are considered one of the earliest placental mammal groups to diverge (Delsuc et al. 2001), there is no evidence for the presence of osteoderms in any of the other basal mammals, suggesting their presence is not plesiomorphic for mammals, although it has been proposed that osteoderms are an example of what has been termed deep homology: a latent but plesiomorphic ability (genetic, cellular, developmental, and structural) to form structures and organs (Hill 2006; Vickaryous and Hall 2008).
In placentals, osteoderms are only present in a single clade, the Xenarthra. The presence of an external carapace formed by articulated osteoderms is one of the major defining features that divide the Xenarthra into two major clades, the armored forms, Cingulata, which includes the living and extinct armadillos and the extinct pampatheres and glyptodonts and the non-armored clade, the Pilosa, “hairy forms,” which includes the living and extinct sloths and New World anteaters (Vermilingua). Contrary to the widespread perception that osteoderms are present in most sloths, their presence has only been documented in a few fossil genera and they are absent from the two living genera, Bradypus and Choloepus. The limited occurrence of sloth taxa with osteoderms suggests they are instead a derived feature limited to only one or two sloth lineages rather than a primitive or plesiomorphic feature of the Xenarthra (e.g., Vickaryous and Hall 2006; Krmpotic et al. 2009, 2014).
The earliest record of the Xenarthra is from the upper Paleocene. The primary record is based on isolated osteoderms from the Sao José de Itaboraí Basin, Brazil (Oliveira and Bergqvist 1998; Bergqvist et al. 2004). In fact, if it was not for the preservation of these osteoderms, the earliest record for the Xenarthra might be significantly younger, as very little of the rest of the skeleton is known for these early taxa. It is noteworthy that even in these earliest cingulate taxa the osteoderms are already quite derived and already articulate with each other along fibrous joints that resemble the sutures between bones of the skull, tend to have consistent sizes and shapes in different parts of the carapace, have a complex ornamentation on the external surface and most likely covered with keratin (scute), and already include both immobile and movable (imbricating) morphologies.
The earliest ascertained fossil sloths in contrast do not appear until the late Oligocene as represented by the genera Orophodon, Octodontotherium, Paroctodontotherium, and Chubutherium (Cattoi 1962; Pujos and De Iuliis 2007). None of these taxa have had osteoderms reported for them. The first major diversification of sloths occurred in the Miocene, during the Santacrucian SALMA, with numerous described genera representing the four major families, Megalonychidae, Nothrotheriidae, Megatheriidae, and Mylodontidae. Despite the intensive study of the Santracucian sloths by Ameghino (1887) and Scott (1903–1904) as well as subsequent researchers (e.g., Bargo et al. 2012), osteoderms have not been reported from any of the Santacrucian sloth taxa, despite the recovery of numerous articulated skeletons. It is not until the Pliocene that osteoderms are described for any sloth taxa (see discussion below). It should also be noted that while the fossil record of the Vermilingua (South American anteaters) is poor (McDonald et al. 2008), osteoderms are not present in the living taxa and have not been reported for any fossil taxa.
In contrast to the osteoderms of the cingulates, the osteoderms of sloths never articulate with each other but are separated by connective tissue in the skin. Consequently, when the skin decays they often become disassociated unlike the cingulates in which articulated carapaces are commonly preserved, although as discussed below occasional osteoderm pavements have been found. See historical background. Within a single individual, the osteoderms are variable in both size and morphology and do not form two distinct functional morphs as in the cingulates. Osteoderms in sloths are also distinguished from those of cingulates in that they are embedded within the skin and are not exposed externally so do not develop a keratinous covering and lack any form of ornamentation. The earliest known osteoderms of cingulates are already more derived in their morphology than any known sloth osteoderms (Hill 2006).
Osteoderm Histology
Histologically the osteoderms of cingulates and sloths are also quite different (Hill 2006; Wolf et al. 2012). Hill (2006) considered the simple isolated osteoderms of sloths plesiomorphic compared to the more highly derived and complex osteoderms of glyptodonts, pampatheres, and armadillos. Based on osteoderms referred to Glossotherium chapadmalense from the Blancan NALMA Haile 15A fauna from Florida and Glossotherium cf. G. harlani from the late Blancan NALMA Inglis 1A fauna of Florida, Hill (2006) described the surface of the osteoderms as typically rough and irregularly pitted. Internally, near the deep surface there is an interweaving network of mineralized fibers oriented in three orthogonal planes with one set perpendicular to the surface. The fibers towards the center were more diffuse with no preferred orientation. The primary bone in the center of the sloth osteoderm is formed by small primary vascular canals enclosed by concentric bone lamellae. In contrast, osteoderms of cingulates have a distinct surficial ornamentation that makes it possible to attribute even isolated osteoderms to a specific taxon. Internally, the osteoderms have a superficial layer composed of collagen fibers oriented perpendicular to the surface. Deeper towards the center of the osteoderm, there is less organization of the collagen fibers and the radially arranged neurovascular channels are enclosed by concentric bone lamellae that exhibit some remodeling. Cingulate osteoderms are highly vascularized with narrow radially oriented channels and a large central cavity (Hill 2006). The imbricating osteoderms have vacuities filled with bone marrow. In pampatheres, the buckler osteoderms have a diplöe or “sandwich” structure, formed by a region of trabecular bone between superficial and deep layers of compact bone similar to that in the membranous bone of the skull and shell of turtles (Wolf et al. 2012; Ciancio et al. in press).
Historical Background
The earliest description of osteoderms attributed to extinct sloths is by Lund (1840). The description of the presence of osteoderms was made in conjunction with specimens that Lund referred to the genera Platyonyx and Megalonyx. However, as many of the generic names utilized by Lund are either invalid because they were preoccupied or the species have since been assigned to other sloth genera, it is difficult to determine to which taxon his description of osteoderms should be assigned, because Lund (1840) did not include specimen numbers in his papers and the specimens were not illustrated. Mylodonts sensu stricto, (excluding scelidotheres), are present in many of the faunas described by Lund so the presence of osteoderms would not be unexpected. It is also possible that some of the osteoderms he interpreted as sloth were from the limbs of cingulates, which are also common members of the fauna. Ameghino (1907), referencing Lund, reported that osteoderms were present in the nothrothere sloth, Nothrotherium, from Lagoa Santa, but Cartelle and Fonseca (1983) in their description of a complete skeleton of this animal determined it did not possess osteoderms.
Burmeister (1867:173, pl. 5, fig. 8) was the first to both describe and illustrate osteoderms attributed to sloths. Osteoderms in dried skin of a sloth from the late Pleistocene Cueva del Milodon (Mylodon Cave), Chile were described and illustrated by Moreno and Woodward (1899), Woodward (1899), and Lönnberg (1899) as either Neomylodon or Grypotherium. Both of these genera and their included species are now considered junior synonyms of Mylodon darwinii. Although numerous pieces of skin of Mylodon darwinii (Fig. 1a, b) have been recovered from the cave, none were associated with any parts of the skeleton so it is not possible to determine where on the body the piece of skin was located. As Mylodon darwinii is the only sloth present in the cave based on both cranial and post-cranial parts of the skeleton, it is not unreasonable to assign the skin fragments to this taxon. However, despite this limitation, the skin fragments from Mylodon Cave are used as the primary reference for the recognition of sloth osteoderms. Isolated osteoderms not preserved in skin referred to Mylodon darwinii have been reported from cave sites in Chile (López-Mendoza and Mena-Larraín 2011).

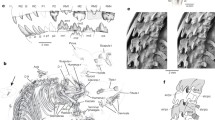
Mummified skin of Mylodon darwinii from Cueva del Milodon, Chile in the Natural History Museum, London. a, external view; b, internal view showing osteoderms. Photo courtesy of John Simmons
Branco (1906) x-rayed portions of the skin of Mylodon darwinii from Mylodon Cave. The x-ray (Fig. 2a, b) showed areas of high osteoderm density with a gradation to areas with lower concentrations of osteoderms to areas where they are absent in the skin. While this does show that the distribution of osteoderms was not uniform and osteoderms were absent in some areas of the skin, as with all other pieces of isolated skin from the cave this piece cannot be positioned on the animal or its orientation determined. Based on the x-rays of portions of the skin illustrated by Branco (1906: pl. 2), three random 10 cm squares were marked and the number of osteoderms per square was counted. The counts were 95, 88, and 83 osteoderms per 10 cm square. In these areas of high density, the osteoderms are in close proximity to each other and form a continuous mosaic but still remain separated by a narrow band of skin. In areas with a lower density of osteoderms, there is more skin between, so they are spaced farther apart. The preserved specimens of sloth skin also show that as in the cingulates, osteoderms in sloths form a single layer. Discoveries of fossil osteoderm layers, in which the osteoderms are stacked on top of each other, probably reflect folds in the skin at the time of burial and should be considered a taphonomic artifact. This is also seen in the large slab of associated osteoderm of Paramylodon harlani (ABDSP (LACM) 1568/V77700) from the Pleistocene of Anza-Borrego State Park, California (McDaniel et al. 2001) (Fig. 3a).

X-rays of mummified skin of Mylodon darwinii from Cueva del Milodon, Chile showing the distribution of osteoderms. a, x-ray of total piece of skin; b, closeup showing details of osteoderms, from Branco (1906)

Comparison of articulated osteoderms of a, Paramylodon harlani (ABDSP(LACM) 1568/V77700) from the Irvingtonian of Anza Borrego Desert State Park, California; and b, mylodontid cf. Glossotherium from the Ensenadan of Tarija, Bolivia (FM P14217). Photos courtesy of Lyndon K. Murray and Bill Simpson, respectively
Cartelle (1991) proposed that Mylodonopsis from the Pleistocene of Brazil is closely related to Mylodon. Although not mentioned in the original description, the partial skeleton has associated osteoderms (pers. observation). Hoffstetter (1952) described the presence of osteoderms of Glossotherium (Oreomylodon) wegneri from the late Pleistocene of Ecuador, which may be a distinct genus Dechaseaux (1971). Unfortunately, he did not provide either measurements or illustrations of the specimens. Osteoderms have been recovered from the asphalt deposits of Talara, Peru (ROM 41973) and three sloths, Glossotherium, Catonyx, and Eremotherium, are known from the fauna. The osteoderms were recovered from screened matrix, so there is no specific association with other skeletal material. Assignment to any of these taxa is tenuous but presumably they are from Glossotherium, although the possibility that they come from Eremotherium cannot be ruled out (see discussion below).
Osteoderms have been recovered from the middle Pleistocene (Ensenadan) of Tarija, Bolivia. Boule and Thévenin (1920:219) mentioned but did not describe or illustrate the specimens they referred to Mylodon robustus var. tarijensis. A specimen from Tarija in the Field Museum (FMNH P14217) is an articulated mosaic of osteoderms (Fig. 3b), so is similar to the Anza-Borrego specimen in that it represents a large piece of skin that was buried. Unfortunately, no other skeletal material was associated with the specimen, so assignment to genus is not possible. The Tarija fauna includes the sloths Megatherium, Nothropus, Catonyx, Lestodon, and Glossotherium. The last three genera are mylodonts, but osteoderms have never been reported for the first two genera and are known only from Glossotherium; it is presumed that these osteoderms can be tentatively referred to this genus. This is the oldest sample of sloth dermal osteoderms in South America. Collins (1933) reported osteoderms from a mylodont sloth from Colombia, South America, which he referred to Mylodon robustus var. tarijensis suggesting it is similar in age to Tarija.
While osteoderms are associated with a skeleton referred to Glossotherium chapadmalense from the Blancan NALMA of Florida (Robertson 1976), none were reported for the type specimen from Argentina (Kraglievich 1925 ). Arzani et al. (2014) described a nearly complete skeleton of Glossotherium robustum from the late Pleistocene of Argentina with well-preserved osteoderms including a layer that was embedded in the skin on top of the head.
Osteoderms in the North American genus Paramylodon were first described by Merriam (1908) based on material recovered from the late Pleistocene asphalt deposits of Rancho La Brea, California. The osteoderms from the Rancho La Brea mylodont were also described by Sinclair (1910) and Stock (1925). The osteoderms overlying a scapula of Paramylodon harlani from Rancho La Brea were described by Sinclair (1910) as larger and often squarish to rhomboid in outline compared to others recovered from the site. Allen (1913) described but did not illustrate osteoderms from the type of Mylodon garmani (= Paramylodon harlani). Shuler (1918:26) mentioned isolated osteoderms found in the sand pits of late Pleistocene age in Dallas County, Texas. Concentrations of osteoderms not associated with other skeletal elements were found at the late Pleistocene Kimmswick Site in Missouri (Graham and Kay 1988). As the osteoderms were recovered by screen washing, it is not possible to determine their original spatial relationships. Graham and Kay (1988) interpreted the concentrations of osteoderms as indicative of in situ decay of skin. They interpreted the lack of associated sloth skeletal elements as suggestive that a piece of skin was brought to the site by the human occupants. However, dried pieces of skin can be transported as well as skeletal elements and other modes for its burial and preservation are also possible. Burial of large pieces of skin resulting in the preservation of the relative position of the osteoderms is uncommon but a few examples are known such as a large set of associated osteoderms from Tarija, Bolivia, and Anza-Borrego Desert State Park, California (McDaniel et al. 2001) previously mentioned.
The earliest documented association of osteoderms with identifiable bones of a sloth skeleton is from two Blancan NALMA localities in North America. Haile 15A, a sinkhole in Florida containing a late Blancan fauna (ca. 2.5 Ma), preserved a single partial skeleton of a mylodont sloth referred to Glossotherium chapadmalense (Robertson 1976). Osteoderms were also present in the type of Glossotherium garbanii from Arroyo El Tanque, Municipio de San Miguel de Allende, Guanajuato, Mexico, also Blancan (ca. 4.4 Ma) (Montellano and Carranza-Castañeda 1981; Montellano-Ballesteros and Carranza-Castañeda 1986; Carranza-Castañeda 2006). They noted that the osteoderms varied in form from rectangular to round and the size varied from 8.2 by 10 mm for the smallest to 26.0 by 12.7 mm for the largest.
Although there are a reasonable number of records of osteoderms that can assigned to a specific sloth taxon, it should be emphasized that there still are many taxa for which they have not been reported. Osteoderms are unknown from the mylodont subfamilies Scelidotheriinae and Urumacotheriinae, indicating that their presence is not a defining character of the family Mylodontidae. Unlike the cingulates in which the distinctive ornamentation of the osteoderms is often utilized for taxonomic identification (Carlini et al. 2009; Krmpotic et al. 2009; Yané 2013), the much simpler morphology of osteoderms in sloths has precluded easy taxonomic identification and it is not possible at this time to refer isolated osteoderms to a specific taxon if the fauna from which they are recovered includes multiple sloth taxa. If consistent morphological features exist that permit the assignment of isolated sloth osteoderms to a genus or species, they have not yet been determined by any comprehensive comparative study. In many cases, osteoderms have been found in isolation but because they were not directly associated with skeletons (Shuler 1918:26) identification beyond sloth was not possible. Therefore, for the present, an association with other parts of the skeleton (e.g., Arzani et al. 2014) is needed or they may not be assignable to a specific taxon, or may be simply overlooked and not reported. While working from negative evidence is never desirable, i.e., absence of reported osteoderms morphologically similar to those seen in other sloths means a taxon did not have osteoderms, in some cases there are enough records to infer they are absent in a specific sloth is a reasonable extrapolation. For example, the middle Miocene La Venta fauna has a diverse array of well-preserved sloths, including two mylodonts Pseudoprepotherium and Glossotheriopsis, that have been sufficiently studied (Hirschfeld 1985; McDonald 1997; Villarroel 1998, 2000) and no osteoderms have been reported, so it is most likely these sloth taxa did not have osteoderms. Given their small size, it could be argued that osteoderms could be easily transported prior to the rest of the skeleton but given the large number present in an individual it is likely that at least a small subset would remain with an associated skeleton.
While most of the sloth genera with osteoderms are mylodonts, Cartelle and Bohórquez (1986) described osteoderms from the late Pleistocene megathere, Eremotherium laurillardi. Eremotherium laurillardi, the Panamerican sloth, has the largest known distribution for any extinct sloth and is known from numerous skeletons (Cartelle and De Iuliis 1995). While this single record is intriguing as osteoderms have never been reported for any other specimens, the presence of a mylodont sloth has been reported from the same locality, so it is possible that this is the source of the osteoderms. Politis and Messineo (2008) reported but did not illustrate any of the 142 osteoderms they assigned to Megatherium americanum, the only sloth from the late Pleistocene (Lujanian SALMA) Campo Laborde site, Argentina, which is also represented by other parts of the skeleton. Given that two glyptodonts, Neosclerocalyptus sp. and Doedicurus sp., are also present in the fauna it cannot be ruled out that these osteoderms are possibly from the legs of either of these two taxa as they are also present in skin of the limbs of Glyptodon (Rinderknecht 2000). If it can be definitely demonstrated that osteoderms are present in these distantly related sloths, mylodonts and megatheres, it further supports the contention of separate acquisition of this feature in sloths, or if plesiomorphic for the Xenarthra it requires the loss of this feature in all other sloths, including the immediate ancestors to the Pliocene and Pleistocene genera in which osteoderms are present.
Phylogenetic Implications
A complete phylogenetic analysis of the relationships of the mylodontid sloths has not been done, although some mylodont taxa have been included in broader studies of sloth relationships (e.g., Gaudin 2004). Based on characters of the mandible for seven South American genera from the subfamily Mylodontinae, Perea (1992) presented their possible relationships but did not include the North American genera, Thinobadistes and Paramylodon. Gaudin (2004) in a more inclusive analysis of sloth relationships based on craniodental characters included 12 genera including scelidotheres, mylodontines, and lestodontines. While neither of these studies included all mylodont genera, what is interesting is that the genera included known to have osteoderms do not cluster together closely in either of these studies. In Perea’s study, Mylodon and Glossotherium do not group together but are in separate clades after the first major branch point. In Gaudin’s study, the three genera with osteoderms, Mylodon, Paramylodon, and Glossotherium, are in a clade that includes three genera for which osteoderms have not yet been documented. However, based on Gaudin’s phylogeny, it is possible that osteoderms might yet be documented for Pleurolestodon, if it is actually a distinct genus from Glossotherium.
Osteoderms are absent in virtually all other sloths including numerous genera within the Mylodontidae. As defined by the craniodental characters of Gaudin (2004), the presence of osteoderms appear to be restricted to a single clade within the subfamily Mylodontinae. This suggests the presence of osteoderms in these related taxa is also a synapomorphy of this clade (Fig. 4). This requires a reexamination of the current taxonomy because three of these genera with osteoderms, Glossotherium, Paramylodon, and Mylodonopsis, are included in the Lestodontinae, while the fourth genus with osteoderms, Mylodon, is placed in a separate subfamily Mylodontinae (McKenna and Bell 1997). McAfee (2009) determined that Glossotherium and Paramylodon share a closer relationship to each other than is indicated by their placement in Gaudin’s cladogram. While the presence of osteoderms is only a single character, and should be only one of many used in a phylogenetic analysis of the mylodonts, a closer examination of its phylogenetic distribution should provide some insight as to whether their presence in different mylodont sloths is a single evolutionary event or was independently acquired in multiple lineages.

Distribution of osteoderms in sloths and suggested relationships. Modified from Gaudin (2004). * indicates genera in which osteoderms have been reported
Conclusions
Based on the above overview I propose that the presence of osteoderms in sloths is not a plesiomorphic character characteristic of the Xenarthra, but rather is an independent acquisition in a small clade of mylodontine sloths. It may also have evolved independently in the megatheres, but the few examples referred to megatheres should be reexamined. The presence of osteoderms in a limited number of sloth taxa in one, possibly two clades indicates the possibility of parallel evolution of this trait in the Cingulata and some pilosans and should be considered a synapomorphy for these sloth taxa in which it is present.
Summary
Osteoderms are present in a wide diversity of tetrapods including varonopsid synapsids, but currently they do not appear to have been present in non-mammalian therapsids, so it is not possible at the moment to trace the presence of osteoderms in the lineage leading to placental mammals.
While xenarthrans are considered one of the earliest placental mammal groups to diverge, there is no evidence for the presence of osteoderms in any of the other basal mammals suggesting that osteoderms are not plesiomorphic for Placentals.
There exist hiatuses between the first appearance of the Xenarthra, as represented by the Cingulata in the late Paleocene, the earliest sloths in the late Oligocene, and the first appearance of osteoderms in sloths in the Pliocene. If osteoderms are plesiomorphic for xenarthrans, there is the inconsistency that they are only present in geologically younger sloths and based on our current state of knowledge absent in the earliest members of the Pilosa. If a plesiomorphic feature, osteoderms should be present in the earliest members of the Megalonychidae, Megatheriidae, and Nothrotheriidae as well as the Mylodontidae, and also in the Vermilingua, and then secondarily lost in geologically younger members of each these clades.
Despite appearing later in the fossil record, the osteoderms in sloths have a simpler and less derived morphology and lack the same level of morphological complexity as osteoderms of the cingulates.
While often considered characteristic of the Mylodontidae, osteoderms are currently only documented in a small number of genera within the family, which despite inclusion in different subfamilies by some studies, are probably closely related and form a single clade within the mylodontines.
The presence of osteoderms in the late Pleistocene megatheres, Eremotherium and possibly Megatherium, members of the Megatheriidae, which is not considered to be closely related to the mylodonts, is probably also independently acquired, but requires more specimens to confirm their presence in these taxa.
Osteoderms in the Pilosa were independently acquired in one lineage of mylodontines, and possibly two genera of megatheres, and are not a plesiomorphic character of the Pilosa.
Abbreviations
- NALMA:
-
North American Land Mammal Age
- SALMA:
-
South American Land Mammal Age
- ABDSP(LACM):
-
Anza Borrego Desert State Park, Borrego Springs, California (specimen originally in the Natural History Museum of Los Angeles County)
- FMNH:
-
Field Museum of Natural History, Chicago, Illinois
References
Allen GM (1913) A new Mylodon. Memoirs of the Museum of Comparative Zoology at Harvard College 40:319-346
Ameghino F (1887) Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de la Patagonia austras y depositados en el Museo La Plata. Boletim Museo La Plata 1:1-26
Ameghino F (1907) Notas sobre una pequeña colección de huesos de mamíferos procedentes de las grutas calcáreas de Iporanga en el Estado de São Paulo, Brasil. Revista do Museu Paulista 7:59-124
Arzani H, Lanzelotti SL, Acuña Suárez GE, Novo NM (2014) Primer registro de pelos fósiles en Glossotherium robustum (Xenarthra, Mylodontidae), Pleistoceno tardío, Mercedes, Provincia de Buenos Aires, Argentina. Ameghinana 51:585-590
Bargo MS, Toledo N, Vizcaíno SF (2012) Paleobiology of the Santacrucian sloths and anteaters (Xenarthra, Pilosa). In: Vizcaíno SF, Kay RF, Bargo MS (eds) Early Miocene Paleobiology in Patagonia. Cambridge University Press, pp 216-242
Bergqvist LP, Abrantes ÉPL, Avilla LDS (2004) The Xenarthra (Mammalia) of Sao José de Itaboraı Basin (upper Paleocene, Itaboraian), Rio de Janeiro, Brazil. Geodiversitas 26:323-337
Boule M, Thévenin (1920) Mammifères fossils de Tarija. Librairie H. le Soudier, Paris, 256 pp
Branco W (1906) Die Anwendung der Röntgenstrahlen in der Paläontologie. Abhandlungen der Königlich Preussischen Akademie der Wissenschaften Berlin:1-55
Burmeister H (1867) Fauna Argentina. I. Mamiferos fósiles. Lista de los mamiferos fósiles del terreno diluviano. Anales del Museo Público de Buenos Aires 1:87-300
Carlini AA, Ciancio MR, Flynn JJ, Scillato-Yané GJ, Wyss AR (2009) The phylogenetic and biostratigraphic significance of new armadillos (Mammalia, Xenarthra, Dasypodidae, Euphractinae) from the Tinguirirican (early Oligocene) of Chile. J Syst Palaeontol 7:489-503
Carranza-Castañeda O (2006) Late Tertiary fossil localities in central Mexico, between 19°-23°N. In: Carranza-Castañeda O, Lindsay EH (eds) Advances in Late Tertiary Vertebrate Paleontology in Mexico and the Great American Biotic Interchange. Universidad Nacional Autónoma de México, Instituto de Geología and Centro de Geociencias, Publicación. Especial 4:45-60
Cartelle C (1991) Um novo Mylodontinae (Edentata, Xenarthra) do Pleistoceno final de região intertropical brasileira. An Acad Bras Ciênc 63:161-170
Cartelle C, Bohórquez GA (1986) Presença de ossículos dérmicos em Eremotherium laurillardi (Lund) Cartelle & Bohorquez, 1982. (Edentata, Megatheriidae). Iheringia 11:3-8
Cartelle C, De Iuliis G (1995) Eremotherium laurillardi: the Panamerican late Pleistocene megatheriid sloth. J Vertebr Paleontol 15:830-841
Cartelle C, Fonseca S (1983) Contribuição ao melhor conhecimento da pequeña preguiça terrícola Nothrotherium maquinense (Lund) Lydekker, 1998. Lundiana 2:127-181
Cattoi N (1962) Un nuevo (Xenarthra) del Terciario de Patagonia, Chubutherium ferelloii gen. et. sp. nov. (Megalonychoidea, Mylodontidae). Revista Museo Argent Ciencias Nacional “Bernardino Rivadavia” Zool 8:123-133
Ciancio MR, Krmpotic CM, Scarano AC, Epele MB (In Press) Internal morphology of osteoderms of extinct armadillos and its relationship with environmental conditions. Journal of Mammalian Evolution
Collins RL (1933) Mylodont (ground sloth) dermal ossicles from Colombia, South America. J Wash Acad Sci 23:426-429
Dechaseaux C (1971) Oreomylodon wegneri, édenté gravigrade du Pléistocène de l'Equateur - Crâne et moulage endocrânien. Ann Paléontol 57:243-285
Delsuc F, Catzeflis FM, Stanhope M J, Douzery EJP (2001) The evolution of armadillos, anteaters and sloths depicted by nuclear and mitochondrial phylogenies: implications for the status of the enigmatic fossil Eurotamandua. Proc Roy Soc B 268:1605-1615
Dilkes DW, Reisz RR (1996) First record of a basal synapsid (‘mammal-like reptile’) in Gondwana. Proc Roy Soc Lond B 263:1165-1170
Gaudin TJ (2004) Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. Zool J Linn Soc 140:255-305
Graham RW, Kay M (1988) Taphonomic comparisons of cultural and noncultural faunal deposits at the Kimmswick and Barnhart Sites, Jefferson County, Missouri. In: Laub RS, Miller NG, Steadman DW (eds) Late Pleistocene and Early Holocene Paleoecology and Archeology of the Eastern Great Lakes Region. Bulletin of the Buffalo Society of Natural Science 33:227-240
Hill RV (2006) Comparative anatomy and histology of xenarthran osteoderms. J Morphol 267:1441-1460
Hirschfeld SE (1985) Ground sloths from the Friasian La Venta fauna, with additions to the Pre-Friasian Coyaima fauna of Colombia, South America. Univ Calif Publ Geol Sci 128:1-91
Hoffstetter R (1952) Les mammifères Pléistocènes de la République de L’Équateur. Mémoires de la Société Géologique de France 66:1-391
Kraglievich L (1925) Cuatro nuevos gravigrados de la Araucana chapadmalense. Anales del Museo Nacional de Historia Natural de Buenos Aires 33:215-235
Krmpotic CM, Ciancio MR, Barbeito C, Mario RC, Carlini AA (2009) Osteoderm morphology in recent and fossil euphractine xenarthrans. Acta Zool 90:339-351
Krmpotic CM, Carlini AA, Galliari FC, Favaron P, Miglino MA, Scarano AC, Barbeito CG (2014) Ontogenetic variation in the stratum granulosum of the epidermis of Chaetophractus vellerosus (Xenarthra, Dasypodidae) in relation to the development of cornified scales. Zoology117:392-397
Lönnberg E (1899) On some remains of “Neomylodon listai” Ameghino brought home by the Swedish Expedition to Tierra del Fuego 1895-1897. Svenska Expeditionen Till Magellansländerna 2:149-170
López-Mendoza P, Mena-Larraín F (2011) Extinct ground sloth dermal bones and their role in the taphonomic research of caves: the case of Baño Nuevo-1 (Andean Central Patagonia, Chile). Revista Mexicana de Ciencias Geológicas 28:519-532
Lund PW (1840) Blik paa Brasiliens Dyreverden för Sidste Jordomvaeltning. Tredie. Afhandling. Fortseattelse af Pattedyrene. Det Kongelige Danske Videnskabernes. Selskabs Naturvidenskabr\elige og Mathematiske Afhandlinger 8:217-272
McAfee RK (2009) Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae). Zool J Linn Soc 155:885-903
McDaniel GE Jr, Jefferson GT, McDonald HG (2001) A large Paramylodon harlani osteoderm layer from the Irvingtonian of Anza-Borrego Desert State Park, California. J Vertebr Paleontol 21:79A
McDonald HG (1997) Xenarthrans: Pilosans. In: Kay RF, Madden RH, Cifelli RL, Flynn JJ (eds) Vertebrate Paleontology in the Neotropics: The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, D.C., pp 231-243
McDonald HG, Vizcaíno SF, Bargo MS (2008) Fossil vermilinguas – an overview. In: Vizcaíno SF, Loughry J (eds) The Biology of the Xenarthra. University of Florida Press, Gainesville, pp 64-78
McKenna MC, Bell SK (1997) Classification of Mammals Above the Species level. Columbia University Press, New York
Merriam JC (1908) Death trap of the ages. Sunset Magazine 21:465-475
Montellano M, Carranza-Castañeda O (1981) Edentados Pliocenicos de la region central de Mexico. Anais II Congresso Latino-Americano Paleontologia, Porto Alegre:683-695
Montellano-Ballesteros M, Carranza-Castañeda O (1986) Descripción de un milodontido del Blancan temprano de la mesa central de Mexico. Universidad Nacional Autónoma de México, Instituto de Geología Revista 6:193-203
Moreno FP, Woodward AS (1899) On a portion of mammalian skin, named Neomylodon listai, from a Cavern near Consuelo Cove, Last Hope Inlet, Patagonia. Proc Zool Soc Lond 67:144-156
Oliveira EV, Bergqvist LP (1998) A new Paleocene armadillo (Mammalia, Dasypodoidea) from the Itaboraí Basin, Brazil. Paleógeno de América del Sur y de la Península Antártica. Asociación Paleontológica Argentina, Publicación Especial 5:35-40
Perea D (1992) Aproximacion a la sistematica filogenetica de Mylodontinae (Xenarthra). Boletín de la Sociedad Zoológica del Uruguay 7:33-34
Politis GG, Messineo PG (2008) The Campo Laborde site: new evidence for the Holocene survival of Pleistocene megafauna in the Argentine Pampas. Quaternary Internatl 191:98-114
Pujos F, De Iuliis G (2007) Late Oligocene Megatherioidea fauna (Mammalia: Xenarthra) from Salla-Luribay (Bolivia): new data on basal sloth radiation and Cingulata-Phyllophaga split. J Vertebr Paleontol 27:132-144
Reisz RR, Modesto SP (2007) Heleosaurus scholtzi from the Permian of South Africa: a varanopid synapsid, not a diapsid reptile. J Vertebr Paleontol 27:734-739
Rinderknecht A (2002) La presencia de osteodermos en las extremidades posteriores de Glyptodon clavipes Owen, 1839 (Mammalia: Cingulata). Ameghiniana 37:369-373
Robertson JS (1976) Latest Pliocene mammals from Haile XVA, Alachua County, Florida. Bull Florida St Mus - Biol Sci 20:111-186
Romer AS (1966) Vertebrate Paleontology. 3rd Edition. University of Chicago Press, Chicago
Scott WB (1903–1904) Mammalia of the Santa Cruz beds. Part 1: Edentata. Report of the Princeton Expeditions to Patagonia 5:1-364
Shuler EW (1918) The geology of Dallas County. University of Texas Bulletin 1818:1-54
Sinclair WJ (1910) Dermal bones of Paramylodon from the asphaltum deposits of Rancho La Brea, near Los Angeles, California. Proc Am Phil Soc 49:191-195
Stock C (1925) Cenozoic gravigrade edentates of western North America with special reference to the Pleistocene Megalonychidae and Mylodontidae of Rancho La Brea. Carnegie Instit Wash Publ 331:1-206
Vickaryous MK, Hall BK (2006) Osteoderm morphology and development in the nine-banded armadillo, Dasypus novemcinctus (Mammalia, Xenarthra, Cingulata). J Morphol 267(11):1273-1283
Vickaryous MK, Hall BK (2008) Development of the dermal skeleton in Alligator mississippiensis (Archosauria, Crocodylia) with comments on the homology of osteoderms. J Morphol 269:398-422
Villarroel CA (1998) Los Nothrotheriinae (Megatheriidae, Gravigrada) del Mioceno de La Venta (Colombia). Descripción de Huilabradys magdaleniensis, nuevos genero y especie. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 22:497-506
Villarroel C (2000) Un nuevo Mylodontinae (Xenarthra, Tardigrada) en la fauna de La Venta, Mioceno de Colombia: el estado actual de la familia Orophodontidae. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 24:117-127
Wolf D, Kalthoff DC, Sander PM (2012) Osteoderm histology of the Pampatheriidae (Cingulata, Xenarthra, Mammalia): implications for systematics, osteoderm growth, and biomechanical adaptation. J Morphol 273:388-404
Woodward AS (1899) On some remains of Grypotherium (Neomylodon) listai and associated mammals from a cavern near Consuelo Cove, Last Hope Inlet, Patagonia. Proc Zool Soc Lond 69:64-78
Yané GJS (2013) Notas sobre los Dasypodidae (Mammalia, Edentata) del Plioceno del Territorio Argentino I. Los restos de edad Chasiquense (Plioceno Inferior) del sur del provincial de Buenos Aires. Ameghiniana 14:133-144
Acknowledgements
Bill Simpson kindly provided the images of the mylodont osteoderms from Tarija at the Field Museum of Natural History, Chicago. John Simmons kindly provided the photos of the mummified skin of Mylodon darwinii housed at the Natural History Museum, London, which granted permission to use the images. I thank the three anonymous reviewers for their comments and suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
McDonald, H.G. An Overview of the Presence of Osteoderms in Sloths: Implications for Osteoderms as a Plesiomorphic Character of the Xenarthra. J Mammal Evol 25, 485–493 (2018). https://doi.org/10.1007/s10914-017-9415-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10914-017-9415-8