
Abstract
Harvester ants have long been known to exhibit interspecific seed preference and this preference has been thought to be associated with distance in a manner analogous with optimal foraging theory. However, little attention has been given to how intraspecific seed preference changes or how microhabitat (i.e. the composition of the terrain that the ants are moving through) impacts seed harvesting preference. We addressed these questions by conducting seed harvesting experiments in three different Ephedra viridis populations that contain harvester ants (Pogonomyrmex occidentalis) by using only E. viridis seeds and conducting trials over multiple distances and varying degrees of microhabitat cover. We found that increased microhabitat cover and increased seed mass decrease the likelihood of E. viridis seeds being harvested much more dramatically than distance. However, we found no effect of distance or microhabitat cover on which E. viridis seeds were harvested. We conclude that harvester ant E. viridis seed preference is distance and microhabitat independent. However, increases in microhabitat cover negatively impacts the likelihood of P. occidentalis harvesting E. viridis seeds of any size. Our findings suggest that harvester ant foraging behavior is influenced by structure of the microhabitat more than by distance. This provides a new context on how harvester ant foraging behavior and effectiveness should be considered.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Within arid environments, seed harvesting by ants is prevalent because of the high nutrition content of seeds and the lack of alternative forms of nutrients (Brown et al. 1979). Harvester ants (Pogonomyrmex spp.) have been shown to significantly impact the seed bank of preferred plant species, removing up to 100 % of preferred seed types (Crist and MacMahon 1992). Although harvester ants may passively disperse seeds by accidentally dropping them or discarding seeds outside of their nest (Detrain and Tasse 2000; Retana et al. 2004; Arnan et al. 2010), the overwhelming majority of seeds collected by harvester ants in the Pogonomyrmex genus are destroyed by being taken deep into their nest where they are either eaten or stored too deep underground to emerge (MacMahon et al. 2000), making this a primarily predatory interaction.
Several studies have shown that harvester ants preferentially harvest bigger seeds (Holder and Polis 1987), seeds with higher nutritional content (Kelrick et al. 1986), weaker seed coats (Rodgerson 1998), and seeds that are not infected by endophytic fungi (Knoch et al. 1993). Although ants preferentially select larger seeds, they have been shown to select seeds in proportion to their body size (Davidson 1977; Crist and MacMahon 1991). Furthermore, ants have been shown to be more selective of seed type over longer distances (Crist and MacMahon 1992), but not over shorter distances (Holder and Polis 1987), in accordance with optimal foraging theory (Schoener 1971).
Ant seed harvesting rate has been shown to be affected by distance and habitat structure. As distance from the mound increases seed removal rates decrease (Kelrick et al. 1986) and seed densities in the soil increase (Crist and MacMahon 1992). Vegetation influences such as the presence of cheatgrass (Ostoja et al. 2013) and shortgrass (Fewell 1988) have been shown to lower foraging rates in harvester ants, presumably due to the greater levels of interference associated with movement across densely vegetated habitat. Conversely, (i.e. the ground cover that impacts an ants journey to and from its harvesting area) with intermediate levels of vegetative cover have been shown to increase harvesting rates (Crist and Wiens 1994).
Despite this research, no studies to our knowledge have examined the effect of microhabitat on intraspecific seed preference in harvester ants. Furthermore, of the studies that have tested the effects of microhabitat structure on seed harvesting rates (Kelrick et al. 1986; Fewell 1988; Crist and Wiens 1994; Ostoja et al. 2013), none have quantified microhabitat structure to determine the extent to which ground cover affects seed selection during harvesting.
Ground coverage in a colonies foraging area may influence ants to adopt a marginal value theorem approach (Charnov 1976) to seed harvesting rather than just an optimal foraging theory (Schoener 1971) one. Under a marginal value model, harvester ant colonies in areas with greater microhabitat cover would have greater difficulty moving larger seeds than smaller seeds across their environment. Ants harvesting in environments with greater microhabitat cover would experience diminishing returns from the resistance associated with moving larger seeds compared to smaller seeds when attempting to harvest. Ants in these types of environments may opt to harvest intermediate or small sized seeds over larger seeds to increase their foraging efficiency. Conversely, areas with less microhabitat cover may exhibit behavior consistent with optimal foraging theory and preferentially harvest larger seeds because movement of larger seeds would not be hindered by resistance from their microhabitat.
Here, we studied how intraspecific seed preference of harvester ants (Pogonomyrmex occidentalis) changed with changes in microenvironment and distance under optimal foraging and marginal value theorem hypotheses. To do this, we conducted seed removal experiments using Ephedra viridis seeds in three harvester ant populations that had ranges overlapping with E. viridis. We chose E. viridis because it has an overlapping range with harvester ants and its seeds are known to be harvested by harvester ants in the Great Basin desert of the western United States. We made two hypothesis and predictions based upon distance and microhabitat cover:
-
1.
We hypothesized that harvester ants with lower microhabitat cover would harvest seeds in a manner concordant with optimal foraging theory. We predicted that as distance from the colony increased, more massive seeds would be selected. At shorter distance all seed phenotypes would be removed.
-
2.
We hypothesized that as microhabitat cover increased, harvester ants would harvest seeds in a manner concordant with marginal value theorem. We predicted that as microhabitat cover increased the mass of seeds harvested by harvest ants would decrease due to the difficulty associated with transporting larger seeds through a more resistant landscape.
Material and Methods
Study Sites
We conducted harvesting experiments in three different E. viridis populations in the western portion of the Great Basin of Nevada and collected seeds from five different populations (Fig. 1). Each study area was a sagebrush steppe community primarily dominated by the shrubs Artemisia tridentata, Ephedra viridis, Chrysothamnus viscidiflorus, Prunus andersonii, and Ericameria nauseosa. Cheatgrass (Bromus tectorum) and several species of bunchgrass were also present at each of the sites. The average monthly precipitation in the communities ranged from 17 to 30 mm.

Locations of sampled Ephedra viridis populations and locations of ant seed harvesting experiments. Locations in California and Nevada. Circles on the map above represent E. viridis populations that we sampled seeds from. Stars represent E. viridis populations that we both sampled seeds from and conducted the harvesting experiments
Seed Measurements and Preparation
We measured E. viridis seeds for mass, length, width, and height. We then labeled and numbered each seed with a color dot system using Uni-Posca paint pens. We labeled the seeds with a colored line to represent their population and two colored dots immediately after the line to represent the seed number. After we labeled and numbered seeds, we measured the length, width, and height using SigmaScan Pro 5. We used these measurements to estimate the volume of each seed by using the equation for an ellipsoid. We used the estimated volume of the seeds as a visual indicator of size in two of our figures.
Seed Harvesting Experiments
We conducted four trials per distance category (1, 2, 3, 4, 5, 6, 7, 8, 9, and 10-m), per population, for a total of 120 trials. Each trial was conducted on a unique ant colony (i.e. no ant colony was used twice). We chose 10-m as the farthest distance because P. occidentalis have been shown to harvest seeds to 10-m from their colonies (MacMahon et al. 2000). Despite observations that harvester ants will move farther in areas with lower vegetative cover (Fewell 1988), we still conducted trials in areas with high vegetation cover covering the 0–10 m range for the experiment. For each trial, we randomly placed measured and labeled seeds under a rodent exclosure made of fine steel mesh (6.35 mm diameter holes) at one of the ten distance categories. We poured the seeds through the top of the rodent exclosure to ensure all seeds fit through the wire mesh. This mesh encapsulated the seeds that were placed on the ground and allowed harvester ants easy access through holes while preventing rodents from accessing the seeds. We surveyed the area around each rodent exclosure to ensure that the ant mound under investigation was the closest one to the seeds in each trial.
We used 60 seeds per trial, with twenty seeds coming from three different E. viridis plants. Seeds from a particular E. viridis were only used once in the experiment. Each plant in a trial was from a different E. viridis population (i.e. there were never two plants from the same population in the same trial). We used seeds from different populations because seeds could only be identified by the population that they came from due to the labeling method used (i.e. if two plants from the same population were used, we would not be able to identify which plant the seed came from because they were marked similarly). We sorted the E. viridis seeds into a small, medium, or large seed category based on the average mass of the seeds (small <12.9 mg; medium 12.9–17.4 mg; large >17.4 mg). We used one plant from the small, medium, and large mass classes in each experiment to ensure a wide range of seed phenotypes in each trial.
We estimated microhabitat cover by measuring the total coverage in a linear transect from the colony to the rodent exclosure. We considered microhabitat cover to be any object (biotic or abiotic) that inhibited movement along a straight path from the ant mound to the location of the seeds (e.g. twigs, leaves, grass, rocks, etc.). We only measured vegetation coming directly out of the ground or objects in direct contact with the ground along the transect, since these could impede the ant’s ability to travel directly to and from its colony.
We collected any remaining seeds after four harvesting periods (48 h) had passed in each trial. We checked for remaining seeds both under the rodent exclosure and along the trunk trail back to the ant mound. We identified the seeds remaining after each trial and marked them as unharvested. We marked all seeds that were not found as harvested. We conducted all trials within each population simultaneously. We conducted all trials in each population over a two-week period from June 28, 2014 through July 6, 2014 to ensure minimal temporal effects on harvesting. We thoroughly surveyed the regions surrounding each ant colony to ensure that the colony being used for a trial was the closest colony to the placed seeds.
Analyses
Statistical analyses were done using the stats package in R, version 2.15 (R Core Team 2016) in concordance with RStudio, version 0.98.1091 (RStudio 2015). Figures were created using ArcMap 10 and ggplot2 (Wickham 2009). All statistical analyses were conducted under a mixed-effects modeling framework using the lme4 package (Bates et al. 2015) with population as a random factor to correct for the spatial autocorrelation and pseudoreplication associated with conducting the experiments in three populations. In order to determine how intraspecific seed preference of harvester ants changed with changes in microenvironment and distance we conducted two different analyses: 1) We tested the effect of seed mass, distance, and microhabitat cover on the likelihood of a seed being harvested, and 2) We tested the effect of distance, and microhabitat cover on the mass of seeds chosen within the trial.
We used logistic regression to assess the effect of seed mass, distance, and microhabitat cover on the likelihood of seeds being harvested. We used simple and multiple regression techniques to assess the effect of distance, and microhabitat cover on the mass of seeds chosen within trials. For this second analysis, we only used harvested seeds from each trial. For both analyses, we first regressed each variable independently to measure their individual effect size and significance. We then developed multiple regression models using the variables from the simple regressions with meaningful effect sizes and statistical significance. Regression models from the analyses were compared using Akaike information criterion (AIC) (Akaike 1974) to aid in assessing the quality of the regression models. We used standardized coefficients for all analyses due to the wide scale of units used to measure the predictor variables.
Results
Effects of Seed Mass, Distance, and Microhabitat Cover on the Likelihood of Seed Harvesting
We found a strong relationship between the mass of harvested and unharvested seeds in the experiment, with smaller seeds being preferentially harvested over larger seeds (Fig. 2). The odds of a seed being harvested were lowered significantly by seed mass, total ground cover, and percent ground cover (Table 1).

The average mass and volume of harvested and unharvested seeds from each trial conducted in the harvesting experiment. Blue triangles represent the average mass and volume of harvested seeds from each trial. Red circles represent the average mass and volume of unharvested seeds from each trial
We developed several simple and multiple logistic regression models to determine the best candidate models for explaining ant seed harvesting preference regarding E. viridis seed mass, harvesting distance, total and percent ground cover (Table 1). Of 17 plausible models, the two best models both contained seed mass as well as either percent cover or total cover, respectively (Table 1).
Effect of Distance and Microhabitat Cover on the Mass of Seeds Harvested
We found no relationship between the mass of harvested seeds with respect to distance, total ground cover, or percent ground cover (Fig. 3). Of the 10 models we developed none were plausible explanations for changes in seed preference based on distance, total ground cover, or percentage of ground cover (Table 2).

Average mass of harvested seeds with respect to distance, total ground cover, and percent ground cover. Mass of seeds were not found to change significantly over distance, total ground cover, or percent ground cover
Discussion
The effects of landscape and distance have long been focal points of harvester ant foraging behavior (MacMahon et al. 2000). However, the historical focus on the effects of landscape has primarily been associated with harvesting rate (Kelrick et al. 1986; Fewell 1988; Crist and Wiens 1994; Ostoja et al. 2013). Furthermore, of the studies that have investigated harvester ant seed preference, the majority of those have focused on interspecific seed preference (Kelrick et al. 1986; Crist and MacMahon 1992; Knoch et al. 1993; Rodgerson 1998) rather than intraspecific preference (Holder and Polis 1987). In this experiment, we used two different analyses to assess the intraspecific seed preference of harvester ants over varying degrees of microhabitat cover using E. viridis seeds. We found that harvester ants preferentially harvest smaller seeds over larger seeds. We concluded that this relationship was a harvesting preference rather than a physiological limitation because a wide range of seed phenotypes were both harvested and unharvested in the experiment (Fig. 4). We also found that the likelihood of a seed being harvested decreases significantly with greater seed mass and ground cover (Table 1). We did not find a significant relationship between distance and the likelihood of seed harvesting (Table 1). Furthermore, we did not find any evidence of harvester ant seed preference when exclusively considering only harvested seeds (Fig. 3; Table 2).

Mass and volume of all harvested and unharvested seeds in the experiment. As in Fig. 2, blue triangles represent harvested seeds and red circles represent unharvested seeds. There were a wide range of both harvested and unharvested phenotypes
Effects of Distance, Percent Ground Cover, Total Ground Cover, and Seed Mass on the Likelihood of E. viridis Seeds Being Harvested
In contrast with the Holder and Polis (1987) study, we found that an increase in seed mass resulted in a decrease in the likelihood of a seed being harvested. The observed preference for larger sized seeds in the Holder and Polis study could have occurred for two reasons: 1) All seed choice experiments were conducted 10 cm from the ant colony, and 2) Seed size classes used in the study were made from milling oats (Avena sp.) into three separate size classes. A 10 cm distance does not reflect the harvesting range of harvester ants (MacMahon et al. 2000) and may mitigate the effects of microhabitat cover. Milling of oats into three separate size classes may also result in a larger size preference because the original seed morphology has been compromised. If we had milled E. viridis seeds into specific size classes, the seed morphology would be altered by artificially increasing the number of edges that a harvester ant could grab on to, potentially allowing them to more easily transport seeds with larger masses.
Our results suggest that the likelihood of E. viridis seeds being harvested is not significantly affected by distance, which is contrary to the results found by Crist and MacMahon (1992). In the Crist and MacMahon (1992) study, the number of seeds present in the soil increased with an increasing distance from the ant colony. However, the Crist and MacMahon (1992) study was conducted in a single locality with a single ant population, while we had a much wider range of microhabitat cover scenarios using three populations (Fig. 5). Variation in microhabitat cover allowed us to test the role landscape resistance may have in foraging decisions by harvester ants.

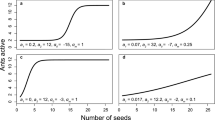
The average percentage of ground cover in each population and the respective mean proportion of seeds harvested given distance. The upper left graph represents the percentage of ground cover that was measured in each study population. When the percentage of ground cover is very large the mean proportion of seeds harvested in the study mimics the expectation that the likelihood of seeds being harvested decreases with distance from the mound. However, when percentage of ground cover is low, the relationship no longer is present
An increase in microhabitat cover and percentage of microhabitat cover were found to significantly decrease the likelihood of E. viridis seeds being harvested and had a much more significant impact than distance (Table 1), which makes sense because of the way that harvester ants move across an open versus a covered microhabitat (Supplementary video, https://www.youtube.com/watch?v=iJCoYLU28mQ). Harvester ants move much faster over open microhabitat than microhabitat that is covered, which would explain why a lower proportion of seeds were harvested with increases in total microhabitat cover as well as percentage microhabitat cover, but not distance (Fig. 6). Interestingly, we found an increase in proportion of seeds removed at intermediate percentages of microhabitat cover, similar to Crist and Wiens (1994).

Proportion of seeds harvested in all experiments given distance, total microhabitat cover, and percentage of microhabitat cover. Distance by itself shows no relationship with proportion of seeds removed. Both total and percentage microhabitat cover show a negative relationship with proportion of seeds harvested
The top two candidate models contained both mass and either total microhabitat cover or percentage microhabitat cover (Table 1). However, the model containing seed mass and total microhabitat cover is likely the better explanation for seed selection in harvester ants because total microhabitat cover still provides a context of distance, while percent microhabitat cover does not.
Effects of Distance and Microhabitat Cover on the Mass of Seeds Harvested
Although smaller seeds were preferentially harvested over larger seeds in our experiment (Figs. 1 and 2), we did not detect a change in seed mass preference under different distance and microhabitat cover regimes (Table 2). This uniform lack of significance and effect size in our analyses suggests that harvester ants prefer smaller E. viridis seeds regardless of the environmental factors being experienced. This is in direct contrast to both optimal foraging theory and marginal value theory hypotheses because both of these predict that there would be changes in seed preference with changes in distance or microhabitat.
Conclusions
Our results show that harvester ants prefer smaller E. viridis seeds. However, there is no evidence to suggest that harvester ants change their seed preference over increased distances or microhabitat cover as would be suggested under an optimal foraging theory or marginal value theorem framework. Since our best statistical model for predicting the likelihood of seed harvesting includes both mass and total microhabitat cover and there is no evidence that increased microhabitat cover results in changes in E. viridis seed choice, we conclude that the size of the seed selected is a behavioral choice by harvester ants and is distance/microhabitat independent. Furthermore, increased microhabitat cover only affects the likelihood of harvester ants harvesting seeds, not seed selection behavior.
We further conclude that when considering animal behavior from an ecological perspective it is important to conduct experiments in multiple populations. Because we took a multi-population approach to addressing questions about intraspecific harvester ant seed preference, we found that microhabitat cover explains seed harvesting behavior much more than distance alone. This most likely would not have been the case if we would have conducted the experiment in a single population.
Lastly, we think that harvester ant seed preference is contingent upon the scale at which the preference is being considered. There has been ample evidence of interspecific seed preference (Kelrick et al. 1986; Crist and MacMahon 1992; Knoch et al. 1993; Rodgerson 1998) but very little information on intraspecific seed preference outside of our study (Holder and Polis 1987). Due to the scarcity of information on intraspecific seed preference and the contradictory conclusions associated with those studies, we think that a greater number of studies focused on intraspecific seed preference of harvester ants would further refine the understanding of harvester ant seed preference.
References
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19:716–723
Arnan X, Retana J, Rodrigo A, Cerdá X (2010) Foraging behaviour of harvesting ants determines seed removal and dispersal. Insect Soc 57:421–430. doi:10.1007/s00040-010-0100-7
Bates D, Maechler M, Bolker B et al (2015) lme4: Linear Mixed-Effects Models using “Eigen” and S4. https://cran.r-project.org/web/packages/lme4/index.html
Brown JH, Reichman OJ, Davidson DW (1979) Granivory in desert ecosystems. Annu Rev Ecol Syst:201–227
Charnov EL (1976) Optimal foraging, the marginal value theorem. Theor Popul Biol 9:129–136
Crist TO, MacMahon JA (1991) Individual foraging components of harvester ants: movement patterns and seed patch fidelity. Insect Soc 38:379–396
Crist TO, MacMahon JA (1992) Harvester ant foraging and shrub-steppe seeds: interactions of seed resources and seed use. Ecology:1768–1779
Crist TO, Wiens JA (1994) Scale effects of vegetation on forager movement and seed harvesting by ants. Oikos:37–46
Davidson DW (1977) Species diversity and community organization in desert seed-eating ants. Ecology:712–724
Detrain C, Tasse O (2000) Seed drops and caches by the harvester ant Messor barbarus: do they contribute to seed dispersal in Mediterranean grasslands? Naturwissenschaften 87:373–376. doi:10.1007/s001140050744
Fewell JH (1988) Energetic and time costs of foraging in harvester ants, Pogonomyrmex occidentalis. Behav Ecol Sociobiol 22:401–408
Holder K, Polis GA (1987) Optimal and central-place foraging theory applied to a desert harvester ant, Pogonomyrmex californicus. Oecologia 72:440–448
Kelrick MI, MacMahon JA, Parmenter RR, Sisson DV (1986) Native seed preferences of shrub-steppe rodents, birds and ants: the relationships of seed attributes and seed use. Oecologia 68:327–337
Knoch TR, Faeth SH, Arnott DL (1993) Endophytic fungi alter foraging and dispersal by desert seed-harvesting ants. Oecologia 95:470–473
MacMahon JA, Mull JF, Crist TO (2000) Harvester ants (Pogonomyrmex spp.): their community and ecosystem influences. Annu Rev Ecol Syst:265–291
Ostoja SM, Schupp EW, Klinger R (2013) Seed harvesting by a generalist consumer is context-dependent: interactive effects across multiple spatial scales. Oikos 122:563–574
R Core Team (2016) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Retana J, Xavier Picó F, Rodrigo A (2004) Dual role of harvesting ants as seed predators and dispersers of a non-myrmechorous Mediterranean perennial herb. Oikos 105:377–385. doi:10.1111/j.0030-1299.2004.12854.x
Rodgerson L (1998) Mechanical defense in seeds adapted for ant dispersal. Ecology 79:1669–1677
RStudio (2015) RStudio: Integrated development environment for R. Boston, MA
Schoener TW (1971) Theory of feeding strategies. Annu Rev Ecol Syst:369–404
Wickham H (2009) ggplot2: elegant graphics for data analysis. Springer, New York
Acknowledgments
We would like to thank the University of Nevada, Reno for providing us with the opportunity to conduct this research. We would like to thank all the undergraduate lab assistants who helped weigh and measure all of the seeds in this experiment. We would also like to thank the reviewers who provided valuable feedback in improving the quality of this manuscript. Lastly, we would like to thank all of the individuals at the University of Nevada, Reno who provided valuable feedback during the development of this project. This work was supported by the National Science Foundation GK-12 Program, Grant DGE No. 1045584 [AH]. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation; This was supported by the University of Nevada, Reno Ecology, Evolution, and Conservation Biology department summer research grant [AH].
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hickey, A., Hollander, J. & Peacock, M. Assessing Microhabitat Cover and Distance Effects on Harvester Ant Intraspecific Seed Preference. J Insect Behav 29, 535–547 (2016). https://doi.org/10.1007/s10905-016-9580-2
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10905-016-9580-2