
Abstract
In this study we investigate associative learning related to food location in Vespula germanica social wasps. Wasps were trained to associate a certain location with food, and their behaviour after food displacement was recorded. The effect of a different number of previous feeding experiences on the detection of the novel food source was evaluated. When these wasps associated a certain location with food, they returned to the original site despite the fact that food was no longer available. This occurred even when a new food source was placed very close to the learned location. The results demonstrate that this learning seems to temporarily impair the detection of new food sources. We discuss how previous experience seems to condition perception, delaying the detection of more rewarding contexts.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Vespula germanica (F.) (Hymenoptera: Vespidae) is a highly invasive social wasp that has become established in a great variety of ecological environments (Archer 1998) It has inhabited North-western Patagonia, Argentina, since 1980, where it has become notoriously widespread in the space of 30 years (Farji-Brener and Corley 1998; D’Adamo et al. 2002). V. germanica is an opportunistic predator and has a highly diverse diet, since it consumes carrion, live arthropods, fruit, honeydew and garbage (Akre 1982). Both environmental variety and eclectic diet facilitate the association of specific contextual conditions with diverse sensory-motor patterns. It has been proposed that phenotypic plasticity might favour invasion, since invasive species often deal with dynamic and changeable contexts (Sakai et al. 2001). In this sense, as V. germanica is exposed to a large and unpredictable range of environmental conditions, cognitive plasticity might play a significant role in their colonization process.
Social wasps usually revisit non depleted food sources (Takagi et al. 1980; Raveret Richter and Jeanne 1985). They learn global and local landmarks for navigating from their nest to the feeding place (Collett and Zeil 1996). By means of “learning flights” wasps acquire landmark information relating to the site they will revisit (Zeil et al. 1996).
As scavengers, V. germanica foragers make several trips between the food source and the nest, where larvae are fed. Thus, relocating a previously discovered food source is a frequent behaviour pattern in their lives. This relocating behaviour has been thoroughly studied under different environmental conditions, supporting evidence of diverse cognitive mechanisms (D’Adamo and Lozada 2003, 2007a, b; Lozada and D’Adamo 2006, 2009). We propose that the recurrent experience of relocating a particular food source implies the strengthening of certain sensory-motor circuits, evidenced in perception and action patterns.
Associative learning is a ubiquitous capacity amongst animals, which enables them to establish predictive relationships between contingent events, thus reducing uncertainty as a result of individual experience. Honeybee associative learning mechanisms have been thoroughly studied in the context of food collection (e.g. Menzel 2009; Giurfa 2007; Cheng and Wignall 2006; Cheng 2005). It has been shown that bees learn both specific links between events in their environment and perform different forms of elemental and non-elemental learning such as categorization, contextual learning, and rule abstraction (review in Menzel 2009; Giurfa 2007). Cognitive abilities in V. germanica have been less studied (e.g. Lozada and D’Adamo 2006), however. Experiments with V. germanica free-flying wasps have shown that they associate food reward with certain visual, spatial and odour cues (D’Adamo and Lozada 2003; Moreyra et al. 2006; D’Adamo and Lozada 2008, 2009; Lozada and D’Adamo 2006, 2009). Moreover, they persist in visiting a feeder after food removal, but rapidly extinguish associations that are no longer rewarding (Lozada and D’Adamo 2006, 2009). Interestingly, the length of time wasps continue their search depends on the number of feeding visits wasps had previously experienced (e.g. Lozada and D’Adamo 2006).
In this study we explore how previous experience conditions perception, by investigating how learning experience interferes with detecting new food sources. We trained wasps to associate a certain location with food, and recorded wasp behaviour after food displacement. We evaluated the effect of a difference in the number of feeding experiences in the detection of the novel food source, which was displaced 60 cm from the original learned feeder location. We predict that wasps will more frequently visit the learned location, no longer rewarding, when a higher number of feeding trials were experienced.
Materials and Methods
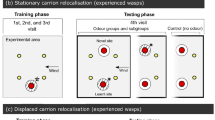
The experiments were conducted in natural, outdoor environments, near San Carlos de Bariloche (41°S, 71°W), Argentina, during the V. germanica wasp’s most active period (March–April) in 2010. All experiments involved feeding individual wasps from a white plastic dish (diameter = 7 cm) containing 20 g of minced bovine meat. This dish was placed on pebbled ground and surrounded by four cylinders of the same colour, 2 cm in diameter and 60 cm in height. The cylinders were arranged as the corners of a square of 30 cm side length, and four dishes were placed at the four edges of the square (Fig. 1). One of these dishes contained food (Fig. 2). When a forager spontaneously arrived at the dish with food, it was distinguishably marked with a dot of washable paint on the abdomen, for future identification. This procedure disturbed wasps only slightly, as they were not captured for marking. Any other wasp visiting the dish was removed, in order to work with only one individual per experiment. The wasp collected a piece of meat, then flew away and returned a few minutes later. Training consisted of either one or four feeding visits, depending on treatment. After these learning trials, the dish with food was displaced from the original feeding location to the opposite point on the array (Fig. 2), i.e. if during training food was located to the West, in the testing phase it was placed to the East. Therefore, when the wasp returned on its second or fifth visit (depending on treatment) it encountered a scenario which was different to the one it had learned, as meat was placed in a different location and a clean, empty dish replaced the baited one. The testing scenario thus consisted of three empty dishes and a baited one (Fig. 2). We operationally define the location where food was originally placed as the “learned” location, the location with food during testing, as the “non learned” location, and the other two, never baited and never learned, as “never rewarded” locations. For control wasps, the dish opposite the baited one is named the “analogous” location, as it resembles the learned location for experimental wasps. During testing, wasp behaviour was scored by recording the number of visits made to each empty dish, until the wasp reached the baited dish and collected a piece of meat. Wasp visits consisted of hovering and landing responses. A hovering episode occurred when the flying wasp remained in the same place, beating its wings for some seconds, over any clean dish but did not land on it. Landing, on the other hand, occurred when the wasp touched, with its six legs, any one of the three clean dishes. An observer who sat at approximately 50 cm from the experimental array recorded wasp responses. One wasp was used for only one experiment. In scoring the data, each location was given 1 point for a hover and 2 points for a landing (Cheng and Wignall 2006; Lozada and D’Adamo 2009). The control group consisted of the same experimental set up with wasps that had no previous learning experience with this array. In this setup, therefore, wasps had to choose between three empty dishes, never rewarded, and one dish with food. As with the experimental group, wasp hovering and landing responses to each dish were recorded. Thus, the comparison between the experimental and control groups reflects the effects of learning experience. We also recorded the time a wasp took from returning to the previously learned location until discovering the new food supply.

Photo of the array utilized for training and testing wasps

Experimental design: Individual wasps were trained to feed from one of four dishes. These dishes were placed around an array composed of four coloured cylinders. During training, wasps fed at a certain location during one or four consecutive visits. During testing, food was displaced to the opposite location
Statistical comparisons on the proportion of wasp visits to the learned and non learned locations were conducted with the Wilcoxon matched pair test within each treatment. Comparisons of wasp visits between all treatments were done using Kruskall Wallis test. Comparisons of data between experimental groups (one or four learning trials) and the control group were conducted with the Mann Whitney U test.
Results
We found significant differences between the number of visits to the empty dish placed at the learned location (where experimental wasps had previously learned to find food) and the number of visits to the analogous location of control wasps (X 2 = 37.67, df = 2, p < 0.0001). Wasps visited the learned location more frequently when trained with one or four learning trials than the analogous location in control wasps (Z = −4.96, N1,2 = 22, 19, p < 0.0001; Z = −5.38, N1,2 = 22, 13, p < 0.0001 respectively). Moreover, significant differences were found between wasps trained with one learning trial and those trained with four (Z = 2.15, N1,2 = 19, 13, p < 0.03) (Fig. 3).

Mean number of wasp visits to the learned location during the testing phase (Mean ± standard error). The three treatments refer to wasps that experienced no learning trials (control), one, or four learning trials
Wasps visited the learned location significantly more frequently than the never rewarded locations, when trained with both one and four learning trials (Z = 3.51, N = 19, p < 0.0005; Z = 3.18, N = 13, p < 0.002 respectively). However, control wasps showed non significant differences between the number of visits to the three empty dishes, as these had never been previously associated with food (Z = 0.53, N = 22, p > 0.59).
Those wasps that had experienced one or four learning treatments visited the never rewarded locations more frequently than control wasps (Z = 4.26, N1, 2 = 19, 22, p < 0.0001; Z = 4.12, N1,2 = 13, 22, p < 0.0007 respectively). This reflects the wasps’ confusion at the disappearance of food from the learned location, something that control wasps did not experience, hence they hardly visited these empty dishes.
Furthermore, significant differences were found in the time spent in finding the novel food source between the three treatments (X 2 = 73.25, df = 2, p < 0.0001) (Fig. 4). Wasps that had fed, either once or four times, spent significantly more time in finding the novel food source than those wasps that had never fed from the array before (Z = 6.75, N1, 2 = 13, 44, p < 0.0001; Z = 8.1, N1,2 = 40, 45 p < 0.0001 respectively). Moreover wasps trained with four learning trials spent significantly more time than those trained with only one (Z = 2.07, N1, 2 = 44, 20, p < 0.04).

Time lapse (in seconds) from wasp’s arrival at the array to discovery of novel food source
Discussion
The present study shows that when V. germanica wasps associate a certain location with food they return to this learned location even if food is no longer there, but is placed very close by. This illustrates the weight of past experience, showing how this can preclude more favourable novel experiences. Thus, learning in this situation could delay the discovery of new food sources, as previous experience seems to condition visual and olfactory food perception. Results from the present study show that when wasps fed from the array only once, they took longer to find the novel dish of meat than those which had never visited the feeder before. So wasps with no previous learning experience found the food source directly, landing straight on the dish with meat, without visiting any other dish. As previous data have shown, food odour is highly attractive, and elicits noticeable landing responses in V. germanica foragers (Moreyra et al. 2006). It is remarkable how just one feeding experience is enough to hinder odour attractiveness, such a salient cue for this species. Furthermore, when wasps had visited the feeder four consecutive times, they took longer to find the novel food source than those that had visited it only once.
Previous studies have shown that the disappearance of food from the learned location leads to extinction of the learned pattern (Lozada and D’Adamo 2006). It has been proposed that the acquisition of a learned response is a function of the number of reinforcements (e.g. Hall 1994; Gallistel et al. 2004). Lozada and D’Adamo (2006) found that V. germanica wasps exploiting a rich food source that suddenly disappears continue visiting the site for a period of time which is related to the number of feeding visits wasps had previously experienced, so wasps that had visited the rewarding dish three times persisted in their search longer than those that had visited it only once (Lozada and D’Adamo 2006). Flexibility could be linked to their capacity to release past experience rather rapidly when facing changes. It is interesting to highlight how one-trial learning is sufficient for this species to establish an association between diverse cues and food reward in different contexts (e.g. D’Adamo and Lozada 2009; Lozada and D’Adamo 2009). However, as demonstrated in the present study, when wasps experience fewer learning trials, the strength of the association is weaker, favouring a quicker response to environmental change. This could be a reflection of the cognitive plasticity of this invasive species, which is continuously dealing with novelty and uncertainty. Flexible mechanisms while foraging have been addressed, as these wasps integrate old and new experiences after very few learning episodes (e.g. D’Adamo and Lozada 2009; Lozada and D’Adamo 2009). In relation to this, Lozada and D’Adamo (2009) have shown that V. germanica is capable of pondering the relative importance of learned patterns and iterative rewarded experiences, with findings in line with results from the present study. To sum up, this evidence indicates that wasps have a vast repertoire of cognitive mechanisms related to the exploitation of an undepleted food source. Being able to return to a relatively constant location for food implies important learning and memory capacities as insects have to memorize resource characteristics, the route to the goal and specific spatial location of the food source with respect to local landmarks (Giurfa 2007). Returning to a large amount of meat may entail beneficial effects given that wasps utilize previous learning, thus reducing food searching time and increasing cost-benefit trade offs (Raveret-Richter 2000). The persistence of an abundant food source provides greater opportunities for learning experience, increasing memory consolidation (Lozada and D’Adamo 2006) and the efficiency of food exploitation and manipulation (unpublished data).
Sensory motor learning capacities have been shown in V. germanica wasps, which can associate different visual stimuli with certain motor responses (D’Adamo and Lozada 2009). For example, if a wasp learns to find food to the west of a blue landmark, and to the east of a yellow landmark, then when presented with the blue landmark, the wasp will direct its search to the west of the landmark, as this learned motor response prevails (D’Adamo and Lozada 2009). These results are in accordance with findings from the present study, as wasps persist in visiting the learned location even when food is located very close by, and odour is strong in the area. This might indicate that certain sensory motor circuits, configured during learning, are also active while trying to relocate the previously visited food source, which could probably delay the activation of novel perceptual-action patterns. Considering all this evidence, we hypothesize that wasps’ previous experience could be reducing perceptual openness, directing certain perception-action processes. Future experiments can be done to elucidate the nature of this interference, given that different possible mechanisms could underlie this process. Wasps could be detecting but ignoring perceived olfactory information, or perception pathways could be turned off while returning to previously learned productive site.
The cognitive flexibility of this social wasp might be related to its ability to release itself from past experience, which could be useful in invasion processes. When evaluating decision-making associated with foraging behaviour, it might be important to take into account that a wasp’s choice implies the consideration of both current and remote experience. We consider free-flying wasp behaviour in changing contexts in natural conditions worthy of study. As an invasive species, V. germanica wasps could constitute a suitable model for analysing cognitive abilities related to environmental uncertainty. Furthermore, it would be useful to analyse cognitive plasticity in other Vespula species which have not shown success in extending their ranges, as this might provide important insights related to the role of learning in invasion processes. Going beyond Vespula, it could be interesting to consider in a general way, how previous experience can have a paradoxical effect, as it can be a limiting factor in the perception of new situations and subsequent favourable choices.
References
Akre RD (1982) Social wasps. In: Hermann HR (ed) Social insects. Academic, New York, pp 1–105
Archer ME (1998) The world distribution of the Euro- Asian species of Paravespula (Hym., Vespinae). Entomol Mon Mag 134:279–284
Cheng K (2005) Context cues eliminate retroactive interference effects in honeybees Apis mellifera. J Exp Biol 208:1019–1024
Cheng K, Wignall AE (2006) Honeybees (Apis mellifera) holding on to memories: response competition causes retroactive interference effects. Anim Cogn 9:141–150
Collett TS, Zeil J (1996) Flights of learning. Curr Dir Psychol Sci 5:149–155
D’Adamo P, Lozada M (2003) The importance of location and visual cues during foraging in Vespula germanica wasps. N Z J Zool 30:171–174
D’Adamo P, Lozada M (2007a) Foraging behaviour related to habitat characteristics in the invasive wasp Vespula germanica. Insect Sci 14:383–388
D’Adamo P, Lozada M (2007b) Foraging behaviour in Vespula germanica wasps when re-locating a food source. N Z J Zool 35:9–17
D’Adamo P, Lozada M (2008) Foraging behavior in Vespula germanica wasps when re-locating a food source. N Z J Zool 35:9–17
D’Adamo P, Lozada M (2009) Flexible foraging behavior in the invasive social wasp Vespula germanica (Hymenoptera: Vespidae). Ann Entomol Soc Am 102(6):1109–1115
D’Adamo P, Sackman P, Rabinovich M, Corley JC (2002) The potential distribution of German wasps (Vespula germanica) in Argentina. N Z J Zool 29:79–85
Farji-Brener A, Corley JC (1998) Successful invasions of hymenopteran insects into NW Patagonia. Ecol Austral 8:237–249
Gallistel CR, Fairhurst S, Balsam P (2004) The learning curve: implications of a quantitative analysis. PNAS 101(36):13124–13131
Giurfa M (2007) Behavioral and neural analysis of associative learning in the honeybee: a taste from the magic well. J Comp Physiol A 193:801–824
Hall G (1994) Pavlovian conditioning: laws of association. In: Mackintosh NJ (ed) Animal learning and cognition. Academic, San Diego, pp 15–43
Lozada M, D’Adamo P (2006) How long do Vespula germanica wasps search for a food source that is no longer available? J Insect Behav 19(5):591–600
Lozada M, D’Adamo P (2009) How does an invasive social wasp deal with changing contextual cues while foraging? Environ Entomol 38(3):803–808
Menzel R (2009) Learning and memory in invertebrates: honey bee. In: Squire LR (ed) Encyclopedia of neuroscience, vol 5. Academic, Oxford, pp 435–439
Moreyra S, D’Adamo P, Lozada M (2006) Odour and visual cues utilized by German yellow-jackets (Vespula germanica) while re-locating protein or carbohydrate resources. Aust J Zool 54:393–397
Raveret-Richter M (2000) Social wasp (Hymenoptera: Vespidae) foraging behaviour. Annu Rev Entomol 45:121–150
Raveret Richter MA, Jeanne RL (1985) Predatory behaviour of Polybia sericea (Olivier), a tropical social wasp (Hymenoptera: Vespidae). Behav Ecol Sociobiol 16:165–170
Sakai AK, Allendorf FW, Holt JS, Lodge DM, Molofsky J, With KA, Baughman S, Cabin RJ, Cohen JE, Ellstrand NC (2001) The population biology of invasive species. Annu Rev Ecol Syst 32:305–332
Takagi M, Hirose Y, Yamasaki M (1980) Prey-location learning in Polistes jadwigae Dalla Torre (Hymenoptera: Vespidae), field experiments on orientation. Kontyu 48(1):53–58
Zeil J, Kelber A, Voss R (1996) Structure and function of learning flights in bees and wasps. J Exp Biol 199:245–252
Acknowledgements
We are very grateful to Audrey Shaw for the revision of the manuscript. This study was partially funded through a grant from CONICET (PIP Nº 114200080100207). We also acknowledge support from the CRUB, Universidad Nacional del Comahue.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lozada, M., D’Adamo, P. Past Experience: a Help or a Hindrance to Vespula germanica Foragers?. J Insect Behav 24, 159–166 (2011). https://doi.org/10.1007/s10905-010-9244-6
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10905-010-9244-6