
Abstract
X-linked agammaglobulinemia (XLA) is clinically characterized by reduced number of peripheral B cells and diminished levels of serum immunoglobulins, and caused by a mutation in the Bruton’s tyrosine kinase (BTK) gene, which play a pivotal role in signal transduction of pre-B-cell receptor (BCR) and BCR. B-cell precursor acute lymphoblastic leukemia (BCP-ALL) is the most common malignancy in children, and it may be associated with gene alterations that regulate B-cell development. Here we described a first case of XLA associated BCP-ALL. The whole-exome sequencing revealed a somatic mutation in MLL2 in the sample from the onset of BCP-ALL. This study suggests that the alterations of BTK and MLL2 synergistically function as leukemogenesis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
X-linked agammaglobulinemia (XLA) is a primary immunodeficiency disease characterized by lack of peripheral B cells and low levels of serum immunoglobulins; it is caused by a mutation of the gene encoding Burton’s tyrosine kinase (BTK) [1]. BTK functions downstream of multiple receptors in various hematopoietic cells. It also plays a pivotal role in signal transduction of pre-B-cell receptor (BCR) and BCR [1]. The activation of pre-BCR and BCR, which results in signal cascade of downstream molecules, is required for the development of B cells, and BTK alterations impede the development of pro-B cells beyond the early pre-B-cell stage to the later stage of pre-B cells [2]. Indeed, XLA patients show severely reduced numbers of mature B cells, whereas pro-B cells are present in the bone marrow of patients with XLA in normal or increased numbers [2]. B-cell precursor acute lymphoblastic leukemia (BCP-ALL) is the most common malignancy in children, and its pathogenesis is explained by the impairment of lymphoid development and normal cellular functions, including cell cycle regulation, tumor suppression and lymphoid signaling [3]. Genome-wide analysis has revealed alterations in genes that regulate B-cell development in more than two-thirds of BCP-ALL patients, including PAX5, TCF3, EBF1, LEF1 and IKZF1 [4]. Recently, germline PAX5 mutations were identified in familial leukemias [5]. Given that impairment of lymphoid development is related to leukemogenesis, there is a possibility that XLA patients have increased susceptibility to BCP-ALL. This is an initial report of a BCP-ALL patient associated with XLA, for whom whole-exome sequencing revealed an acquired mutation in MLL2. This study was approved by the ethics board of the University of Toyama.
Material and Methods
Patient Report
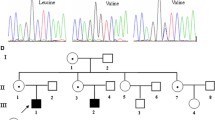
The patient was a 10-year-old boy, who had contracted pneumococcal meningitis at the age of 3 years and had suffered from recurrent bacterial infections including otitis media. The family history was unremarkable. The percentage of B cells and serum immunoglobulin levels were markedly reduced (B cells 0.3 %, IgG 7 mg/dl, IgA 3 mg/dl, IgM 30 mg/dl). At the age of 6 years, the patient was diagnosed as having XLA with a BTK c.1051A > G (p.R307G) missense mutation (Fig. 1a), the pathogenicity of which has been reported in a public database (SH2base: http://bioinf.uta.fi/SH2base/BTK.html). Regular intravenous immunoglobulin (IVIG) replacement therapy was started. When he was 10 years old, a routine hematological study incidentally disclosed lymphoblasts in the peripheral blood. His white blood cell count was 7.0 × 109/L with 15.0 % lymphoblasts, hemoglobin was 12.0 g/dL and platelet count was 185,000/μL. Bone marrow aspiration revealed hypercellular marrow, with 96.2 % lymphoblasts. Flow cytometry revealed that the lymphoblasts were positive for CD10 (99.7 %), CD19 (99.7 %), CD22 (97.9 %), cy-CD79a (96.4 %), CD34 (99.8 %) and cy-TdT (95.4 %) and HLA-DR (98.8 %) and negative for CD20, cy-Igμ chain, Igμ chain, Igκ chain, Igλ chain, and all of the T-lineage and myeloid markers except CD2 (99.2 %). The karyotype was normal, and real-time polymerase chain reaction detected no major BCR-ABL1, minor bcr-abl1, TCF3-PBX, MLL-AF4, MLL-AF9, MLL-ENL, ETV6-RUNX1 and SIL-TAL1 fusion. The patient was diagnosed with BCP-ALL, and received chemotherapy according to the Tokyo Children’s Cancer Study Group (TCCSG) ALL L04-16 high-risk group protocol. He had a good clinical course, and the chemotherapy was just finished. No severe complications occurred except for one episode of febrile neutropenia.

BTK expression and its role in leukemogenesis. a Intracellular BTK expression in CD14+ monocytes evaluated by flow cytometry (BD LSRFortessaTM Cell Analyzer, BD Biosciences, San Diego, CA). Black histograms indicate BTK expression (clone 10E10, OriGene Technologies, Inc., Rockville, MD), and Gray histograms indicate isotype control. BTK expression in the patient was reduced compared with that in a normal control. b Two different roles of BTK in the development of B-cell malignancies. Absence of BTK-dependent BCR signals may promote the development of poorly differentiated B-cell malignancies. Activated BTK-dependent BCR signals promote the development of well-differentiated B-cell malignancies
Whole-Exome Sequencing
Whole-exome sequencing was performed to identify secondary genetic events. Paired leukemia cell DNA from bone marrow cells at diagnosis and control DNA from peripheral blood mononuclear cells in remission were analyzed. Exome capture was carried out using a TruSeq Exome Enrichment Kit (Illumina) according to the manufacturer’s protocol. Massively-parallel sequencing was performed using a HiSeq 1500 platform (Illumina) with 100 bp-paired-end reads. Sequence reads were aligned to the human reference genome using Burrows-Wheeler Aligner. SAMtools, Genome Analysis Toolkit (GATK) and Picard Tools were used for downstream processing and variant calling. Calls with a read coverage < 5 × were filtered out, resulting in > 96 % coverage for the entire target region. Detected tumor-specific candidate variants were further filtered using dbSNP137, Human Genetic Variation Database (HGVD; http://www.genome.med.kyoto-u.ac.jp/SnpDB/) and our in-house SNP database. As a result, six tumor-specific variants were identified (Table 1). Among these variants, a MLL2 c.8740delC (p.H2914fs) mutation was considered a driver mutation, because it is an inactivating mutation and the gene is reported to be recurrently inactivated in diffuse large B-cell lymphoma (DLBCL) [6, 7]. For the other five variants, we performed an extensive literature search but could not find reports supporting their driver role. We also evaluated 237 novel germline nonsynonymous variants that were not registered in SNP databases. However, even with knowledge from genome-wide association studies of ALL and the Catalogue of Somatic Mutations in Cancer (COSMIC) database [8], we could not identify possibly pathogenic variants except the BTK c.1051A > G mutation.
Discussion
It is tempting to hypothesize that a germline BTK mutation and an acquired MLL2 mutation are sufficient to cause a leukemic transformation in B lymphocyte precursors with defective maturation. The development of leukemia cells was arrested just before the stage that required BTK. Previous studies identified that several BCP-ALL cells have aberrant expression of BTK caused by somatic mutations or aberrant transcription of BTK [9]. It is noteworthy that many of these observations were identified in MLL-rearranged ALL [9]. MLL2 is a histone methyltransferase and its impaired function results in aberrant DNA methylation. Haploinsufficiency of MLL2 is sufficient to drive molecular pathogenesis. A heterozygous germline mutation results in a characteristic systemic disorder named Kabuki syndrome [10], and the somatic MLL2 mutation observed in DLBCL is also heterozygous [6, 7]. However, the fact that our current knowledge about gene mutations remains markedly limited and the possibility of the existence of a fusion gene that we have not tested weaken the hypothesis.
Of greatest interest was that BCP-ALL with an MLL2 mutation occurred in a BTK-deficient patient whereas BTK inhibitor is effective in some DLBCL patients [11], most likely having MLL2 mutation. According to our findings and previous reports, we suggest two different roles of BTK in the pathogenesis of B-cell malignancies according to the degree of their differentiation (Fig. 1b). Firstly, constitutively or aberrantly activated BTK-dependent signals from BCR can promote the development of well-differentiated B-cell malignancies [12, 13]. Secondly, absence of BTK-dependent signals, which result in developmental arrest of B cells, can promote the development of poorly differentiated B-cell malignancies such as BCP-ALL. Our report provides the first evidence of a germline BTK mutation and an acquired MLL2 mutation affecting leukemogenesis, as well as a new insight into germline mutations in leukemia development.
References
Tsukada S, Saffran DC, Rawlings DJ, Parolini O, Allen RC, Klisak I, et al. Deficient expression of a B cell cytoplasmic tyrosine kinase in human X-linked agammaglobulinemia. Cell. 1993;72:279–90.
Nomura K, Kanegane H, Karasuyama H, Tsukada S, Agematsu K, Murakami G, et al. Genetic defect in human X-linked agammaglobulinemia impedes a maturational evolution of pro-B cells into a later stage of pre-B cells in the B-cell differentiation pathway. Blood. 2000;96:610–7.
Inaba H, Greaves M, Mullighan CG. Acute lymphoblastic leukaemia. Lancet. 2013;381:1943–55.
Mullighan CG, Su X, Zhang J, Radtke I, Phillips LA, Miller CB, et al. Deletion of IKZF1 and prognosis in acute lymphoblastic leukemia. N Engl J Med. 2009;360:470–80.
Shah S, Schrader KA, Waanders E, Timms AE, Vijai J, Miething C, et al. A recurrent germline PAX5 mutation confers susceptibility to pre-B cell acute lymphoblastic leukemia. Nat Genet. 2013;45:1226–31.
Morin RD, Mendez-Lago M, Mungall AJ, Goya R, Mungall KL, Corbett RD, et al. Frequent mutation of histone-modifying genes in non-Hodgkin lymphoma. Nature. 2011;476:298–303.
Pasqualucci L, Trifonov V, Fabbri G, Ma J, Rossi D, Chiarenza A, et al. Analysis of the coding genome of diffuse large B-cell lymphoma. Nat Genet. 2011;43:830–7.
Urayama KY, Chokkalingam AP, Manabe A, Mizutani S. Current evidence for an inherited genetic basis of childhood acute lymphoblastic leukemia. Int J Hematol. 2013;97:3–19.
Feldhahn N, Río P, Soh BN, Liedtke S, Sprangers M, Klein F, et al. Deficiency of Bruton’s tyrosine kinase in B cell precursor leukemia cells. Proc Natl Acad Sci U S A. 2005;102:13266–71.
Ng SB, Bigham AW, Buckingham KJ, Hannibal MC, McMillin MJ, Gildersleeve HI, et al. Exome sequencing identifies MLL2 mutations as a cause of Kabuki syndrome. Nat Genet. 2010;42:790–3.
Wilson WH, Gerecitano JF, Goy A, de Vos S, Kenkre VP, Barr PM, et al. The Bruton’s tyrosine kinase (BTK) inhibitor, ibrutinib (PCI-32765), has preferential activity in the ABC subtype of relapsed/refractory de novo diffuse large B-cell lymphoma (DLBCL): interim results of a multicenter, open-label, phase 2 study. Blood. 2012;120:686 (abstract).
Aalipour A, Advani RH. Bruton tyrosine kinase inhibitors: a promising novel targeted treatment for B cell lymphomas. Br J Haematol. 2013;163:436–43.
de Rooij MF, Kuil A, Geest CR, Eldering E, Chang BY, Buggy JJ, et al. The clinically active BTK inhibitor PCI-32765 targets B-cell receptor- and chemokine-controlled adhesion and migration in chronic lymphocytic leukemia. Blood. 2012;119:2590–4.
Acknowledgments
This study was supported by grants from the Ministry of Health, Labour, Welfare of Japan. We thank Chikako Sakai and Hitoshi Moriuchi for their excellent technical assistance. This study is dedicated to the late Dr. Toshio Miyawaki.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hoshino, A., Okuno, Y., Migita, M. et al. X-Linked Agammaglobulinemia Associated with B-Precursor Acute Lymphoblastic Leukemia. J Clin Immunol 35, 108–111 (2015). https://doi.org/10.1007/s10875-015-0127-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10875-015-0127-7