
Abstract
Careful regulation of the body’s immunoglobulin G (IgG) and albumin concentrations is necessitated by the importance of their respective functions. As such, the neonatal Fc receptor (FcRn), as a single receptor, is capable of regulating both of these molecules and has become an important focus of investigation. In addition to these essential protection functions, FcRn possesses a number of other functions that are equally as critical and are increasingly coming to attention. During the very first stages of life, FcRn mediates the passive transfer of IgG from mother to offspring both before and after birth. In the adult, FcRn regulates the persistence of both IgG and albumin in the serum as well as the movement of IgG, and any bound cargo, between different compartments of the body via transcytosis across polarized cells. FcRn is also expressed by hematopoietic cells; consistent with this, FcRn regulates MHC class II presentation and MHC class I cross-presentation by dendritic cells. As such, FcRn plays an important role in immune surveillance throughout adult life. The increasing appreciation for FcRn in both homeostatic and pathological conditions is generating an intense interest in the potential for therapeutic modulation of FcRn binding to IgG and albumin.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The neonatal Fc receptor for immunoglobulin (FcRn) was first isolated from rodent gut as a heterodimer between a 12 kDa and a 40-50 kDa protein [1, 2] and subsequently cloned in 1989 by Simister and Mostov [3]. These now-classic studies identified FcRn as the receptor responsible for the previously recognized pathway associated with passive acquisition of immunoglobulin G (IgG) by rodents upon breast-feeding during neonatal life and were the culmination of nearly two decades of investigation by a number of scientists, beginning with Francis Brambell, who examined this immunologic phenomenon [4, 5]. With the cloning and subsequent crystallographic resolution of FcRn structure, it was quickly recognized that the molecule responsible for these physiologic properties possessed a major histocompatibility complex (MHC) class I-like structure that was in non-covalent association with ß2-microglobulin (ß2m) [6, 7]. The identification of FcRn has led to a rapid expansion in our appreciation for this interesting molecule; its function is now recognized to include a number of important activities that extend well beyond the neonatal life of rodents and to involve numerous different organ systems in a wide variety of animal systems [8]. This review briefly summarizes the physiologic and pathophysiologic functions of FcRn and the opportunities that this knowledge provides for therapeutic manipulation.
FcRn is an IgG Binding Molecule that Functions throughout Life
FcRn consists of an approximately 50-kDa heavy chain that is similar in structure to MHC class 1 and is in non-covalent association with the 12-kDa ß2m light chain. X-ray crystallographic studies of rodent and human FcRn have shown that, in contrast to classical MHC class 1 molecules (HLA-A, B, and C of human and H-2 K in mouse), the potential groove formed by the α1 and α2 domains of the heavy chain that serves as a peptide binding site in MHC class 1 is narrowed in FcRn [9]. Consistent with this, biochemical studies and co-crystallographic studies have revealed that FcRn has adopted a structure that allows it to function in the binding of both IgG and albumin [10, 11]. These properties of FcRn are due to ionic interactions between FcRn and IgG or albumin on opposite faces of the FcRn heavy chain [10]. In the case of IgG, this involves a variety of residues on the α1 and α2 domains of FcRn, including leucine 135, aspartic acid 130, tryptophan 131, glutamic acid 116, glutamic acid 115, and isoleucine 114; these interact with critical amino acid residues within the CH2 and CH3 regions of the Fc domain of IgG (Fig. 1). In terms of cross species differences, rat and mouse FcRn are 91 % identical in their peptide sequence [12] whereas human FcRn has only 65 % similarity in its amino acid sequence compared to rat FcRn [13]. Generally, human FcRn shows very limited interspecies IgG binding, extending only to rabbit IgG. Rodent FcRn, however, is known to be promiscuous by binding to IgG molecules from a variety of species including human, rabbit and bovine IgG [14].

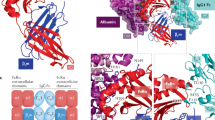
Crystal structure of human FcRn. FcRn is a heterodimer consisting of a MHC class I-like heavy chain (blue) that is non-covalently associated with ß2-microglobulin (ß2m, green). Critical residues for the interaction with IgG molecules are indicated in yellow. The binding of albumin to FcRn (marked in red) occurs at a distinct site from that of IgG, with the conserved histidine 166 as a critical residue for pH-dependent binding to albumin. Adapted from [22]
FcRn binding amino acid residues in the IgG Fc domain specifically include isoleucine 253, histidine 310 and histidine 435 [15–20]. Apart from these, the residue 436, corresponding to histidine in mouse IgG1 and tyrosine in human IgG1, has been shown to be involved in binding of IgG to FcRn, although to a lesser degree than the previously noted residues [20, 21]. In comparison, albumin interactions with FcRn likely involve histidine 161 and histidine 166 on FcRn and three highly conserved histidine residues (histidine 464, histidine 510, and histidine 535) within domain 3 of albumin [10, 22, 23]. Binding of FcRn to albumin and IgG is strictly pH dependent. Binding occurs at acidic pH—typically less than pH 6 (optimally at pH 5), but not at neutral pH or above pH 7. Whereas FcRn binds IgG at a 2:1 ratio, it binds albumin with a stoichiometry of 1:1 [7, 10, 24, 25]. Consistent with this pH dependency, FcRn binding to IgG is abrogated by mutation of these critical residues within the IgG molecule. This pH-dependent binding is consistent with the predominance of FcRn within an intracellular locale—namely, in endosomes—and the interaction between FcRn and its ligands within an intracellular milieu [26–28].
Although FcRn was originally viewed as a neonatally restricted molecule based upon the physiology that was initially linked to it, as discussed above, it is now well accepted that FcRn has a much broader distribution of expression and function, both throughout life and in many cell types within multiple organ systems [29]. FcRn is now recognized to be expressed in both parenchymal cells (e.g., the epithelium of the intestine, lung, and liver) and hematopoietic cells (e.g., monocytes, macrophages, dendritic cells, polymorphonuclear leukocytes, and B cells) in both mouse and human throughout life [29]. Developmental regulation of FcRn exists, but to date it is known to occur only in the intestinal epithelium of the rodent, where FcRn expression is decreased 1000-fold at the time of weaning; and in the placenta, where FcRn functions to transport IgG from mother to fetus antenatally [2, 30, 31]. In contrast, FcRn in the epithelium of other mammals continues to be expressed at moderate levels throughout life [32]. FcRn homologues have been described in mouse, rat, human, non-human primates, cow, pig, sheep, possum, and horse [33–36]. An orthologous molecule has been described in chicken; this molecule is more similar to the phospholipase A2 receptor than to FcRn [37]. These characteristics of FcRn are consistent with the broad range of functions that is now recognized to be associated with this molecule.
FcRn Functions in the Bidirectional Transport of IgG
Originally, FcRn was shown to be responsible for the transfer of maternal IgG across the intestinal epithelium of neonatal rodents into the bloodstream to provide passively acquired immunity [3, 4]. This transfer process has also been linked to the regulation of mucosal tolerance, including that associated with distant sites such as the lung [38]. In a similar fashion, FcRn is recognized to be responsible for the transfer of maternal IgG to a human fetus via transport across the placenta [30, 39, 40].
Physiologically, such transport mechanisms are due to the transcytosis of IgG across polarized epithelial cells, which is now known to be a bidirectional process. As such, FcRn mediates the bidirectional transport (or transcytosis) of IgG across all polarized epithelial barriers in which it has been examined. These include in vitro model systems for a variety of polarized epithelial types that model functions in the intestines, lung, placenta, genitourinary system, and kidney [39, 41–46]. Moreover, such characteristics of FcRn have been documented in model systems derived from human, rat, mouse, pig, and non-human primates. Thus, expression of FcRn in a variety of polarized epithelia confers upon the epithelial barrier an ability to transport IgG, but not polymeric IgA or IgM, in an apical-to-basal or basal-to-apical direction. In this manner, FcRn can deliver IgG into the lumen from the tissue space and transport it back to the lamina propria of a variety of tissues. This system is active in all organ systems examined to date, as noted, but is likely to be especially active in certain portions of the gastrointestinal tract (e.g., rectum), upper airway, and genitourinary system, where it accounts for the high concentrations of IgG relative to IgA and IgM.
These in vitro observations have also been demonstrated in vivo. For example, it has been shown that FcRn is expressed in the upper airway epithelial cells of mice, where it is able to transport Fc fusion proteins in an apical-to-basal direction—that is, from the lumen to the bloodstream [38]. This movement is FcRn dependent, as mutation of the FcRn binding site in the Fc fragment of IgG (I253, H435, and H436) abolishes this transport [38]. In a similar fashion, FcRn has been demonstrated to specifically transport IgG from the bloodstream into the intestines in a mouse model that was forced transgenically to express FcRn in the intestinal epithelium but nowhere else in the host [47]. These studies demonstrate that in rodents, bidirectional transport of IgG can take place in vivo.
Corroboration of this has been provided by studies in non-human primates and in humans. For example, studies have shown that FcRn is expressed at significant levels in the upper airways (e.g., bronchi and trachea) of non-human primates and humans. Application of Fc fusion proteins of human IgG1 as a chimeric molecule with erythropoietin (EPO) is associated with transport across the upper airway epithelium of non-human primates into the bloodstream [48, 49]. As shown in mouse models, mutation of the same FcRn binding site within the Fc domain of the fusion proteins disrupts this transport. Confirmation that this occurs in humans has been demonstrated by successful completion of phase 1 clinical studies assessing the efficacy of delivery of an Fc-EPO fusion protein into the bloodstream of normal healthy volunteers; transport of the Fc-EPO fusion proteins in the upper airways was mediated by FcRn in a dose-dependent fashion [50]. These studies not only demonstrate functional FcRn expression in the epithelia of the lung during adult life, but also have laid the basis for the possibility for transepithelial delivery of different types of macromolecular cargo when chimerized to the Fc fragment of IgG. This has indeed been confirmed for Fc fusion proteins associated with not only EPO, but also interferon-α, interferon-β, and follicle-stimulating hormone [49, 51]. This demonstrates the potential utility of FcRn in the transmucosal delivery of therapeutic macromolecules.
FcRn Functions in the IgG-mediated Transport and Retrieval of Luminal Antigens
The ability of FcRn to bidirectionally transport IgG from the tissues in the intestinal lumen and back to the lamina propria is associated with the retrieval of specific antigens from the lumen [52]. Studies have shown that this ability to transcytose IgG-antigen complexes is bidirectional [43, 47, 52]. Such observations have been demonstrated not only with in vitro systems, but also through the use of a novel humanized mouse model in which a human FcRn transgene is expressed under the control of its own endogenous promoter, together with transgenic expression of human ß2m in a mouse that is devoid of mouse FcRn expression [27, 52]. Such humanized mice express levels of FcRn within the intestinal epithelium similar to that observed in an adult human small intestine. This mouse model system has shown that antigen within the lumen can be retrieved by intravenously administered immunoglobulin molecules specific for that antigen. These IgG molecules then form immune complexes with the corresponding antigen within the lumen, are retrieved by the epithelial cell, and are transported into the lamina propria. There they can be internalized by antigen-presenting cells such as dendritic cells [52]. Such transport mechanisms are capable of promoting a T-cell–specific response in distant lymphoid structures such as mesenteric lymph nodes and spleen (Fig. 2). A similar pathway has been demonstrated in the upper airway, indicating that this is a general function of FcRn in multiple different mucosal sites.

FcRn-mediated IgG transport and antigen retrieval across the intestinal epithelial barrier. FcRn in enterocytes mediates the transcytosis of IgG molecules from the basolateral side into the gastrointestinal lumen. Following formation of antigen-IgG immune complexes, FcRn is then able to bind these immune complexes and transcytose them back to the basolateral side, where the IgG-antigen complex is delivered to antigen-presenting cells (APC) in the lamina propria. Antigen-loaded APCs then migrate to draining lymph nodes and initiate a T-cell response
This transport pathway is physiologically relevant, since this process can be linked to the protection of mice from infection with an epithelial-specific pathogen—Citrobacter rodentium, which models enteropathogenic Escherichia coli infection [47]. Only when FcRn is expressed in the epithelium and in the presence of C. rodentium-specific IgG is the rodent able to mount an effective immune response against this epithelial pathogen [47]. The fact that expression of FcRn within the epithelium, together with specific expression of antigen-specific IgG molecules, is able to promote the activation of antigen-specific T cells within regional lymphoid structures and associated peripheral tissues provides evidence that epithelial expression of FcRn is able to link luminal and/or epithelial infectious exposures with systemic immune activation [47, 52]. Consistent with this, FcRn transport of IgG across the gastric, genitourinary, and lung epithelium is associated with protection from viral infections (e.g., influenza and herpes simplex virus) and Helicobacter heilmanii at these sites [42, 46, 53]. In a similar manner, the application of IgG-antigen complexes to the mucosal surfaces of the lung can promote an antigen-specific immune response consistent with the prediction that FcRn-mediated transport processes have the potential for being applied to vaccination strategies [54].
FcRn Functions in IgG and Albumin Protection from Catabolism
Consistent with previous predictions by Brambell more than 40 years ago, FcRn is now known to be the factor responsible for the long half-life of IgG and albumin [11, 26, 27, 55–57]. In this process, FcRn has been shown in in vitro model systems to internalize these macromolecules by fluid phase endocytosis into an acidified endosome wherein FcRn can bind IgG and protect it from degradation by trafficking IgG (and albumin) away from a degradative fate in lysosomes and recycling them to the cell surface [26, 58–60]. As such, IgG and albumin possess half-lives of 21 and 16 days, respectively, in humans. Proof of these processes has come from studies in mice genetically deficient in FcRn expression wherein both hypogammaglobulinemia and hypoalbuminemia are observed, with a commensurate decrease in serum half-life for each of these molecules [11, 26, 27, 55–57]. Although parenchymal cells such as the endothelium likely play a significant role in IgG protection—as shown by conditional deletion of FcRn in this cell type in vivo [61]—it is now clear that a significant fraction of IgG protection is due to the activities of hematopoietic cells. This is shown by studies using bone marrow chimeras with FcRn-deficient and FcRn-sufficient cells [55, 62]. The latter is consistent with the functional expression of FcRn in a wide variety of hematopoetic cell types in mouse and human, as discussed above.
These observations have important therapeutic implications. Specifically, they raise the possibility that engineering IgG molecules with enhanced FcRn binding can increase IgG half-life and thus improve the pharmacokinetic and pharmacodynamic properties of therapeutic antibodies (as reviewed in detail by Presta [63] and Kuo [64]), Fc fusion proteins, and potentially albumin-based fusion proteins. This has indeed been accomplished, as discussed below, through the generation of monomeric Fc fusion proteins. On the other hand, blockade of FcRn binding of IgG potentially enables the degradation of pathogenic antibodies. This can be accomplished by administration of intravenous immunoglobulins (IVIG), which work in part through this mechanism by saturating FcRn with irrelevant antibodies [65–68]. Alternatively, antibodies and peptides have been developed that can block FcRn-IgG interactions and increase IgG catabolism [69–72]. Such approaches have been effective, with demonstrable activity in autoimmune and IgG-mediated disease models [65, 67, 68, 73, 74].
FcRn Functions in Antigen Presentation by Professional Antigen-Presenting Cells
The expression of FcRn in professional antigen-presenting cells (APC) has prompted an investigation of the role that FcRn plays not only in the protection of IgG from catabolism, but also in antigen presentation. As such, it has been demonstrated that FcRn in mouse and human dendritic cells regulates MHC class II-restricted antigen presentation of model antigens such as ovalbumin, in the case of mouse, and human antigens such as gliadin, which is associated with celiac sprue [59, 75]. Dendritic cells that are deficient in FcRn, or that are provided with IgG-containing immune complexes unable to bind FcRn through site-directed mutagenesis of the FcRn binding sites within the Fc fragment of IgG, have proven that FcRn plays a significant role in MHC class II-mediated antigen presentation to CD4+ T lymphocytes both in vitro and in vivo [59].
Similarly, FcRn recently has been demonstrated to play a role in antigen presentation to CD8+ T cells by dendritic cells [76]. This pathway is called cross-presentation and involves the crossover of extracellular antigens into an MHC class I-dependent pathway. Classically, extracellular antigens are internalized, processed, and presented via MHC class II to CD4+ T cells. However, the antigens generated for presentation by MHC class I molecules to CD8+ T cells are derived from the processing of cytosolic proteins by proteasomes and the transport of these nominal peptides into the endoplasmic reticulum (ER) by the transporter associated with antigen processing (TAP) [77, 78]. In the process of cross-presentation, however, extracellular antigens gain access to the MHC class I-dependent pathway through internalization of the antigen into specialized vacuoles. These vacuoles exhibit less vigorous degradation of internalized proteins, and loading of the generated peptides onto the MHC I molecule occurs either within the internalized vacuole (so-called vacuolar pathway) or, after transport of the internalized polypeptide from the vacuole into the cytosol for processing and presentation, by the classical pathway associated with proteasomes (so-called cytosolic pathway) [78].
New studies by Baker and colleagues have shown that FcRn plays a critical role in cross-presentation and is especially important in the activities of a subset of dendritic cells that were previously not recognized to play a significant role in such processes [76]. Previous studies have shown that the majority of cross-presentation related to the processing and presentation of extracellular antigens is conferred by the activities of dendritic cells that express CD8 [79, 80]. In contrast, when antigen is provided as an IgG-containing immune complex, dendritic cells that are CD8−CD11b+CD11c+ (or inflammatory dendritic cells) display significant cross-presentation at low-antigen doses in a pathway highly dependent upon FcRn expression [76]. This pathway involves the internalization of the IgG-containing immune complexes by Fcγ receptors into an acidic endosome, leading to the hand-off of the immune complex to FcRn, which is competent to bind IgG and IgG-containing immune complexes within this milieu [76]. Crosslinked FcRn, in turn, is able to direct the transport of the internalized immune complex into a compartment characterized by significant levels of vacuolar ATPase (responsible for endosomal acidification), gp91phox (NOX2; responsible for oxidation and buffering endosomal acidification), Sec61 (responsible for transport of the internalized polypeptide from the vesicle to the cytosol), TAP (the transporter associated with peptide transport from the cytosol to the vesicle or to the ER), MHC class I, and Rab27 (an adaptor molecule responsible for gp91phox mobilization) [76]. In the absence of FcRn expression or the ability of the IgG to bind to FcRn, so-called inflammatory dendritic cells are disabled in their ability to properly participate in cross-presentation [76] (Fig. 3). FcRn-dependent antigen presentation is likely to be physiologically relevant, since antibacterial-specific immunoglobulin molecules are only pathogenic in vivo during colitis when FcRn functionally exists in APC [62]. Moreover, inflammatory dendritic cells that are able to function in FcRn-dependent cross-presentation are observed to accumulate in the intestines during colitis [76]. These studies in their totality demonstrate an important functional role of FcRn in antigen presentation and cross-presentation.

FcRn-dependent cross-presentation of immune complexes. Following internalization by surface Fcγ receptors, FcRn regulates the intracellular sorting of IgG ICs into a Rab27a, vacuolar ATPase, Sec61, TAP, gp91phox, and MHC class I-containing acidic phagosome that allows for cross-presentation. In the absence of FcRn expression or the ability of the IgG to bind to FcRn, CD8-CD11b+CD11c+ inflammatory dendritic cells are disabled in their ability to properly participate in cross-presentation. Adapted from [76]
IgG-FcRn Fc Engineering: Fc Mutations that Increase FcRn Binding
Those properties that relate to the intimate relationship that exists between FcRn and IgG and its association with a variety of different physiologic and potentially pathophysiologic pathways have opened up certain possibilities: to engineer and develop new forms of therapeutic intervention (e.g., transmucosal delivery of macromolecular therapeutics); to ameliorate autoimmunity (e.g., administration of IVIG or agents that are able to interfere with FcRn-IgG binding); or to engineer antibody-based therapeutics to alter their pharmacokinetics and, potentially, pharmacodynamics. As shown in Table I, a wide variety of mutations in the CH2-CH3 domain interface around the FcRn binding site of IgG have significant effects on IgG interactions with FcRn and, consequently, on in vivo half-life. Moreover, in at least one case, such mutations have been shown to increase the efficacy of a therapeutic monoclonal antibody [81]. These successes highlight the potential opportunities that exist in modulating FcRn-IgG interactions [81].
Conclusions
FcRn is now recognized to play an important role in a wide variety of physiologic functions, based upon its interactions with its two major ligands, IgG and albumin. In this manner, FcRn is responsible for the bidirectional transport of IgG and IgG antigen/antibody complexes across polarized epithelia. This process plays an important role in mucosal protection from infection, protection of monomeric IgG and albumin from catabolism via FcRn expression in parenchymal and hematopoietic cells—which accounts for the long half-lives of these important macromolecules—and, finally, in both MHC class I cross-presentation and MHC class II presentation by professional APC, leading to adaptive immunity. Such knowledge will play an important role in driving the discovery of novel means to prevent and treat a variety of conditions that are directly and indirectly related to the activities of these interactions and related pathways.
References
Rodewald R, Kraehenbuhl JP. Receptor-mediated transport of IgG. J Cell Biol. 1984;99(1 Pt 2):159s–64s.
Simister NE, Rees AR. Isolation and characterization of an Fc receptor from neonatal rat small intestine. Eur J Immunol. 1985;15(7):733–8. doi:10.1002/eji.1830150718.
Simister NE, Mostov KE. An Fc receptor structurally related to MHC class I antigens. Nature. 1989;337(6203):184–7.
Brambell FW. The transmission of immune globulins from the mother to the foetal and newborn young. Proc Nutr Soc. 1969;28(1):35–41.
Brambell FW, Hemmings WA, Morris IG. A Theoretical Model of Gamma-Globulin Catabolism. Nature. 1964;203:1352–4.
Raghavan M, Gastinel LN, Bjorkman PJ. The class I major histocompatibility complex related Fc receptor shows pH-dependent stability differences correlating with immunoglobulin binding and release. Biochemistry. 1993;32(33):8654–60.
Huber AH, Kelley RF, Gastinel LN, Bjorkman PJ. Crystallization and stoichiometry of binding of a complex between a rat intestinal Fc receptor and Fc. J Mol Biol. 1993;230(3):1077–83.
Roopenian DC, Akilesh S. FcRn: the neonatal Fc receptor comes of age. Nat Rev Immunol. 2007;7(9):715–25.
Burmeister WP, Gastinel LN, Simister NE, Blum ML, Bjorkman PJ. Crystal structure at 2.2 A resolution of the MHC-related neonatal Fc receptor. Nature. 1994;372(6504):336–43.
Chaudhury C, Brooks CL, Carter DC, Robinson JM, Anderson CL. Albumin binding to FcRn: distinct from the FcRn-IgG interaction. Biochemistry. 2006;45(15):4983–90.
Chaudhury C, Mehnaz S, Robinson JM, Hayton WL, Pearl DK, Roopenian DC, et al. The major histocompatibility complex-related Fc receptor for IgG (FcRn) binds albumin and prolongs its lifespan. J Exp Med. 2003;197(3):315–22.
Ahouse JJ, Hagerman CL, Mittal P, Gilbert DJ, Copeland NG, Jenkins NA, et al. Mouse MHC class I-like Fc receptor encoded outside the MHC. J Immunol. 1993;151(11):6076–88.
Story CM, Mikulska JE, Simister NE. A major histocompatibility complex class I-like Fc receptor cloned from human placenta: possible role in transfer of immunoglobulin G from mother to fetus. J Exp Med. 1994;180(6):2377–81.
Ober RJ, Radu CG, Ghetie V, Ward ES. Differences in promiscuity for antibody-FcRn interactions across species: implications for therapeutic antibodies. Int Immunol. 2001;13(12):1551–9.
Firan M, Bawdon R, Radu C, Ober RJ, Eaken D, Antohe F, et al. The MHC class I-related receptor, FcRn, plays an essential role in the maternofetal transfer of gamma-globulin in humans. Int Immunol. 2001;13(8):993–1002.
Kim JK, Firan M, Radu CG, Kim CH, Ghetie V, Ward ES. Mapping the site on human IgG for binding of the MHC class I-related receptor, FcRn. Eur J Immunol. 1999;29(9):2819–25.
Kim JK, Tsen MF, Ghetie V, Ward ES. Localization of the site of the murine IgG1 molecule that is involved in binding to the murine intestinal Fc receptor. Eur J Immunol. 1994;24(10):2429–34.
Martin WL, West Jr AP, Gan L, Bjorkman PJ. Crystal structure at 2.8 A of an FcRn/heterodimeric Fc complex: mechanism of pH-dependent binding. Mol Cell. 2001;7(4):867–77.
Raghavan M, Bonagura VR, Morrison SL, Bjorkman PJ. Analysis of the pH dependence of the neonatal Fc receptor/immunoglobulin G interaction using antibody and receptor variants. Biochemistry. 1995;34(45):14649–57.
Shields RL, Namenuk AK, Hong K, Meng YG, Rae J, Briggs J, et al. High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J Biol Chem. 2001;276(9):6591–604.
Medesan C, Matesoi D, Radu C, Ghetie V, Ward ES. Delineation of the amino acid residues involved in transcytosis and catabolism of mouse IgG1. J Immunol. 1997;158(5):2211–7.
Andersen JT, Dee Qian J, Sandlie I. The conserved histidine 166 residue of the human neonatal Fc receptor heavy chain is critical for the pH-dependent binding to albumin. Eur J Immunol. 2006;36(11):3044–51.
Andersen JT, Dalhus B, Cameron J, Daba MB, Plumridge A, Evans L, et al. Structure-based mutagenesis reveals the albumin-binding site of the neonatal Fc receptor. Nat Commun. 2012;3:610. doi:10.1038/ncomms1607.
Sanchez LM, Penny DM, Bjorkman PJ. Stoichiometry of the interaction between the major histocompatibility complex-related Fc receptor and its Fc ligand. Biochemistry. 1999;38(29):9471–6.
Schuck P, Radu CG, Ward ES. Sedimentation equilibrium analysis of recombinant mouse FcRn with murine IgG1. Mol Immunol. 1999;36(15–16):1117–25.
Junghans RP, Anderson CL. The protection receptor for IgG catabolism is the beta2-microglobulin-containing neonatal intestinal transport receptor. Proc Natl Acad Sci U S A. 1996;93(11):5512–6.
Roopenian DC, Christianson GJ, Sproule TJ, Brown AC, Akilesh S, Jung N, et al. The MHC class I-like IgG receptor controls perinatal IgG transport, IgG homeostasis, and fate of IgG-Fc-coupled drugs. J Immunol. 2003;170(7):3528–33.
Ward ES, Zhou J, Ghetie V, Ober RJ. Evidence to support the cellular mechanism involved in serum IgG homeostasis in humans. Int Immunol. 2003;15(2):187–95.
Kuo TT, Baker K, Yoshida M, Qiao SW, Aveson VG, Lencer WI, et al. Neonatal Fc receptor: from immunity to therapeutics. J Clin Immunol. 2010;30(6):777–89.
Simister NE, Story CM, Chen HL, Hunt JS. An IgG-transporting Fc receptor expressed in the syncytiotrophoblast of human placenta. Eur J Immunol. 1996;26(7):1527–31.
Martin MG, Wu SV, Walsh JH. Ontogenetic development and distribution of antibody transport and Fc receptor mRNA expression in rat intestine. Dig Dis Sci. 1997;42(5):1062–9.
Baker K, Qiao SW, Kuo T, Kobayashi K, Yoshida M, Lencer WI, et al. Immune and non-immune functions of the (not so) neonatal Fc receptor, FcRn. Semin Immunopathol. 2009;31(2):223–36.
Adamski FM, King AT, Demmer J. Expression of the Fc receptor in the mammary gland during lactation in the marsupial Trichosurus vulpecula (brushtail possum). Mol Immunol. 2000;37(8):435–44.
Kacskovics I, Wu Z, Simister NE, Frenyo LV, Hammarstrom L. Cloning and characterization of the bovine MHC class I-like Fc receptor. J Immunol. 2000;164(4):1889–97.
Mayer B, Zolnai A, Frenyo LV, Jancsik V, Szentirmay Z, Hammarstrom L, et al. Redistribution of the sheep neonatal Fc receptor in the mammary gland around the time of parturition in ewes and its localization in the small intestine of neonatal lambs. Immunology. 2002;107(3):288–96.
Schnulle PM, Hurley WL. Sequence and expression of the FcRn in the porcine mammary gland. Vet Immunol Immunopathol. 2003;91(3–4):227–31.
West Jr AP, Herr AB, Bjorkman PJ. The chicken yolk sac IgY receptor, a functional equivalent of the mammalian MHC-related Fc receptor, is a phospholipase A2 receptor homolog. Immunity. 2004;20(5):601–10.
Spiekermann GM, Finn PW, Ward ES, Dumont J, Dickinson BL, Blumberg RS, et al. Receptor-mediated immunoglobulin G transport across mucosal barriers in adult life: functional expression of FcRn in the mammalian lung. J Exp Med. 2002;196(3):303–10.
Antohe F, Radulescu L, Gafencu A, Ghetie V, Simionescu M. Expression of functionally active FcRn and the differentiated bidirectional transport of IgG in human placental endothelial cells. Hum Immunol. 2001;62(2):93–105.
Leach JL, Sedmak DD, Osborne JM, Rahill B, Lairmore MD, Anderson CL. Isolation from human placenta of the IgG transporter, FcRn, and localization to the syncytiotrophoblast: implications for maternal-fetal antibody transport. J Immunol. 1996;157(8):3317–22.
Akilesh S, Huber TB, Wu H, Wang G, Hartleben B, Kopp JB, et al. Podocytes use FcRn to clear IgG from the glomerular basement membrane. Proc Natl Acad Sci U S A. 2008;105(3):967–72.
Bai Y, Ye L, Tesar DB, Song H, Zhao D, Bjorkman PJ, et al. Intracellular neutralization of viral infection in polarized epithelial cells by neonatal Fc receptor (FcRn)-mediated IgG transport. Proc Natl Acad Sci U S A. 2011. doi:10.1073/pnas.1115348108.
Claypool SM, Dickinson BL, Wagner JS, Johansen FE, Venu N, Borawski JA, et al. Bidirectional transepithelial IgG transport by a strongly polarized basolateral membrane Fcgamma-receptor. Mol Biol Cell. 2004;15(4):1746–59.
Claypool SM, Dickinson BL, Yoshida M, Lencer WI, Blumberg RS. Functional reconstitution of human FcRn in Madin-Darby canine kidney cells requires co-expressed human beta 2-microglobulin. J Biol Chem. 2002;277(31):28038–50.
Ellinger I, Rothe A, Grill M, Fuchs R. Apical to basolateral transcytosis and apical recycling of immunoglobulin G in trophoblast-derived BeWo cells: effects of low temperature, nocodazole, and cytochalasin D. Exp Cell Res. 2001;269(2):322–31.
Li Z, Palaniyandi S, Zeng R, Tuo W, Roopenian DC, Zhu X. Transfer of IgG in the female genital tract by MHC class I-related neonatal Fc receptor (FcRn) confers protective immunity to vaginal infection. Proc Natl Acad Sci U S A. 2011;108(11):4388–93. doi:10.1073/pnas.1012861108.
Yoshida M, Kobayashi K, Kuo TT, Bry L, Glickman JN, Claypool SM, et al. Neonatal Fc receptor for IgG regulates mucosal immune responses to luminal bacteria. J Clin Invest. 2006;116(8):2142–51.
Bitonti AJ, Dumont JA, Low SC, Peters RT, Kropp KE, Palombella VJ, et al. Pulmonary delivery of an erythropoietin Fc fusion protein in non-human primates through an immunoglobulin transport pathway. Proc Natl Acad Sci U S A. 2004;101(26):9763–8.
Bitonti AJ, Dumont JA. Pulmonary administration of therapeutic proteins using an immunoglobulin transport pathway. Adv Drug Deliv Rev. 2006;58(9–10):1106–18.
Dumont JA, Bitonti AJ, Clark D, Evans S, Pickford M, Newman SP. Delivery of an Erythropoietin-Fc Fusion Protein by Inhalation in Humans through an Immunoglobulin Transport Pathway. J Aerosol Med. 2005;18(3):294–303.
Low SC, Nunes SL, Bitonti AJ, Dumont JA. Oral and pulmonary delivery of FSH-Fc fusion proteins via neonatal Fc receptor-mediated transcytosis. Hum Reprod. 2005;20(7):1805–13.
Yoshida M, Claypool SM, Wagner JS, Mizoguchi E, Mizoguchi A, Roopenian DC, et al. Human neonatal Fc receptor mediates transport of IgG into luminal secretions for delivery of antigens to mucosal dendritic cells. Immunity. 2004;20(6):769–83.
Ben Suleiman Y, Yoshida M, Nishiumi S, Tanaka H, Mimura T, Nobutani K, et al. Neonatal Fc receptor for IgG (FcRn) expressed in the gastric epithelium regulates bacterial infection in mice. Mucosal Immunol. 2012;5(1):87–98. doi:10.1038/mi.2011.53.
Ye L, Zeng R, Bai Y, Roopenian DC, Zhu X. Efficient mucosal vaccination mediated by the neonatal Fc receptor. Nat Biotechnol. 2011;29(2):158–63.
Akilesh S, Christianson GJ, Roopenian DC, Shaw AS. Neonatal FcR expression in bone marrow-derived cells functions to protect serum IgG from catabolism. J Immunol. 2007;179(7):4580–8.
Ghetie V, Hubbard JG, Kim JK, Tsen MF, Lee Y, Ward ES. Abnormally short serum half-lives of IgG in beta 2-microglobulin-deficient mice. Eur J Immunol. 1996;26(3):690–6.
Israel EJ, Wilsker DF, Hayes KC, Schoenfeld D, Simister NE. Increased clearance of IgG in mice that lack beta 2-microglobulin: possible protective role of FcRn. Immunology. 1996;89(4):573–8.
Ober RJ, Martinez C, Vaccaro C, Zhou J, Ward ES. Visualizing the site and dynamics of IgG salvage by the MHC class I-related receptor, FcRn. J Immunol. 2004;172(4):2021–9.
Qiao SW, Kobayashi K, Johansen FE, Sollid LM, Andersen JT, Milford E, et al. Dependence of antibody-mediated presentation of antigen on FcRn. Proc Natl Acad Sci U S A. 2008;105(27):9337–42.
Tzaban S, Massol RH, Yen E, Hamman W, Frank SR, Lapierre LA, et al. The recycling and transcytotic pathways for IgG transport by FcRn are distinct and display an inherent polarity. J Cell Biol. 2009;185(4):673–84.
Montoyo HP, Vaccaro C, Hafner M, Ober RJ, Mueller W, Ward ES. Conditional deletion of the MHC class I-related receptor FcRn reveals the sites of IgG homeostasis in mice. Proc Natl Acad Sci U S A. 2009;106(8):2788–93.
Kobayashi K, Qiao SW, Yoshida M, Baker K, Lencer WI, Blumberg RS. An FcRn-dependent role for anti-flagellin immunoglobulin G in pathogenesis of colitis in mice. Gastroenterology. 2009;137(5):1746–56e1.
Presta LG. Molecular engineering and design of therapeutic antibodies. Curr Opin Immunol. 2008;20(4):460–70. doi:10.1016/j.coi.2008.06.012.
Kuo TT, Aveson VG. Neonatal Fc receptor and IgG-based therapeutics. MAbs. 2011;3(5):422–30. doi:10.4161/mabs.3.5.16983.
Akilesh S, Petkova S, Sproule TJ, Shaffer DJ, Christianson GJ, Roopenian D. The MHC class I-like Fc receptor promotes humorally mediated autoimmune disease. J Clin Invest. 2004;113(9):1328–33.
Hansen RJ, Balthasar JP. Intravenous immunoglobulin mediates an increase in anti-platelet antibody clearance via the FcRn receptor. Thromb Haemost. 2002;88(6):898–9.
Hansen RJ, Balthasar JP. Effects of intravenous immunoglobulin on platelet count and antiplatelet antibody disposition in a rat model of immune thrombocytopenia. Blood. 2002;100(6):2087–93.
Li N, Zhao M, Hilario-Vargas J, Prisayanh P, Warren S, Diaz LA, et al. Complete FcRn dependence for intravenous Ig therapy in autoimmune skin blistering diseases. J Clin Invest. 2005;115(12):3440–50.
Vaccaro C, Zhou J, Ober RJ, Ward ES. Engineering the Fc region of immunoglobulin G to modulate in vivo antibody levels. Nat Biotechnol. 2005;23(10):1283–8.
Vaccaro C, Bawdon R, Wanjie S, Ober RJ, Ward ES. Divergent activities of an engineered antibody in murine and human systems have implications for therapeutic antibodies. Proc Natl Acad Sci U S A. 2006;103(49):18709–14. doi:10.1073/pnas.0606304103.
Mezo AR, Sridhar V, Badger J, Sakorafas P, Nienaber V. X-ray Crystal Structures of Monomeric and Dimeric Peptide Inhibitors in Complex with the Human Neonatal Fc Receptor, FcRn. J Biol Chem. 2010;285(36):27694–701.
Mezo AR, McDonnell KA, Hehir CA, Low SC, Palombella VJ, Stattel JM, et al. Reduction of IgG in nonhuman primates by a peptide antagonist of the neonatal Fc receptor FcRn. Proc Natl Acad Sci U S A. 2008;105(7):2337–42.
Liu L, Garcia AM, Santoro H, Zhang Y, McDonnell K, Dumont J, et al. Amelioration of experimental autoimmune myasthenia gravis in rats by neonatal FcR blockade. J Immunol. 2007;178(8):5390–8.
Hansen RJ, Balthasar JP. Pharmacokinetic/pharmacodynamic modeling of the effects of intravenous immunoglobulin on the disposition of antiplatelet antibodies in a rat model of immune thrombocytopenia. J Pharm Sci. 2003;92(6):1206–15.
Qiao SW, Sollid LM, Blumberg RS. Antigen presentation in celiac disease. Curr Opin Immunol. 2009;21(1):111–7. doi:10.1016/j.coi.2009.03.004.
Baker K, Qiao SW, Kuo TT, Aveson VG, Platzer B, Andersen JT, et al. Neonatal Fc receptor for IgG (FcRn) regulates cross-presentation of IgG immune complexes by CD8-CD11b+ dendritic cells. Proc Natl Acad Sci U S A. 2011;108(24):9927–32.
Cresswell P, Ackerman AL, Giodini A, Peaper DR, Wearsch PA. Mechanisms of MHC class I-restricted antigen processing and cross-presentation. Immunol Rev. 2005;207:145–57. doi:10.1111/j.0105–2896.2005.00316.x.
Amigorena S, Savina A. Intracellular mechanisms of antigen cross presentation in dendritic cells. Curr Opin Immunol. 2010;22(1):109–17. doi:10.1016/j.coi.2010.01.022.
Hildner K, Edelson BT, Purtha WE, Diamond M, Matsushita H, Kohyama M, et al. Batf3 deficiency reveals a critical role for CD8alpha+ dendritic cells in cytotoxic T cell immunity. Science. 2008;322(5904):1097–100. doi:10.1126/science.1164206.
Jongbloed SL, Kassianos AJ, McDonald KJ, Clark GJ, Ju X, Angel CE, et al. Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J Exp Med. 2010;207(6):1247–60. doi:10.1084/jem.20092140.
Zalevsky J, Chamberlain AK, Horton HM, Karki S, Leung IW, Sproule TJ, et al. Enhanced antibody half-life improves in vivo activity. Nat Biotechnol. 2010;28(2):157–9.
Hinton PR, Xiong JM, Johlfs MG, Tang MT, Keller S, Tsurushita N. An engineered human IgG1 antibody with longer serum half-life. J Immunol. 2006;176(1):346–56.
Petkova SB, Akilesh S, Sproule TJ, Christianson GJ, Al Khabbaz H, Brown AC, et al. Enhanced half-life of genetically engineered human IgG1 antibodies in a humanized FcRn mouse model: potential application in humorally mediated autoimmune disease. Int Immunol. 2006;18(12):1759–69.
Dall’Acqua WF, Kiener PA, Wu H. Properties of human IgG1s engineered for enhanced binding to the neonatal Fc receptor (FcRn). J Biol Chem. 2006;281(33):23514–24.
Acknowledgments
We apologize to all of our colleagues we have not cited due to space limitations. Timo Rath receives support from the German Research Foundation Grant no. RA 2040/1-1. Timothy Kuo was supported by NIH DK071798. Kristi Baker was supported by The Canadian Institutes for Health Research. Masaru Yoshida receives support from the Global COE Program, Global Center of Excellence for Education and Research on Signal Transduction Medicine in the Coming Generation and Grant-in-Aid for Scientific Research (C) from the Japan Society for the Promotion of Science (JSPS). Derry Roopenian is supported by NIH CA034196 and AR054407. Edda Fiebiger is supported by the NIH AI075037. Wayne Lencer is supported by NIH DK48106, DK084424, DK090603 and Harvard Digestive Diseases Center (NIH P30DK034854). Richard Blumberg is supported by NIH DK44319, DK51362, DK53056, DK88199 and Harvard Digestive Diseases Center (NIH P30DK034854).
Conflict of Interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rath, T., Kuo, T.T., Baker, K. et al. The Immunologic Functions of the Neonatal Fc Receptor for IgG. J Clin Immunol 33 (Suppl 1), 9–17 (2013). https://doi.org/10.1007/s10875-012-9768-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10875-012-9768-y