
Abstract
Ofunato Bay was a semi-closed area because of the breakwater effect at the entrance; however, the breakwater was destroyed by a massive tsunami generated by the 2011 off the Pacific coast of Tohoku Earthquake. Consequently, the physical environment of Ofunato Bay has been changed significantly, i.e., the modification of the stratified structure of seawater inside the bay and the intermittent intrusion of seawater outside the bay. These alterations of physical environment are considered to have an influence on the chemical and biological environment in Ofunato Bay. To elucidate the influence of the tsunami on the aquatic environment, we measured dissolved nutrients, chlorophyll a and dissolved oxygen concentrations, and heterotrophic bacteria abundance inside and outside of Ofunato Bay from 2012 to 2014, and compared these data with those obtained before the earthquake. As compared with before the earthquake, significant changes after the earthquake were (1) decrease of ammonium and phosphate concentrations, (2) increase of chlorophyll a concentration, (3) increase of dissolved oxygen concentration in the bottom, and (4) decrease of heterotrophic bacteria abundance. The collapse of the breakwater and consequential enhanced water exchange were considered to have brought the decrease of nutrient concentration inside the bay. Furthermore, washout of shellfish mariculture rafts by the tsunami decreased the shellfish biodeposits along with the elution of nutrients by heterotrophic bacteria. Decrease of cultivated shellfish further caused a decline in feeding pressure on phytoplankton and, subsequently, increased the phytoplankton biomass that contributed to the decrease of nutrients inside the bay.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
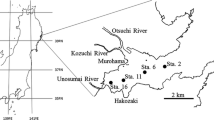
Ofunato Bay is located on the eastern coast of the Tohoku area of Japan and faces the western North Pacific. It stretches from the south to north with a length of 5.2 km and width of 2 km (Fig. 1). Because of its intricate coastline and calm environment, shellfish mariculture (oysters and scallops) have developed inside the bay. Ofunato Bay has been damaged by tsunami many times in the past. In 1960, the Great Chilean Earthquake caused a tsunami that reached the Pacific coast of Japan, killing 53 people in and around Ofunato Bay. Consequently, in 1967, a huge breakwater was constructed at the mouth of the bay. Because of the breakwater, the width of the entrance of Ofunato Bay narrowed from 850 to 200 m, and the depth of the bay entrance decreased from 36 to 16 m.

Sampling sites of Ofunato Bay, Iwate, northern Japan
To determine the effects of this huge artificial structure on the environment of Ofunato Bay, Miyazawa and Hayakawa (1994) compared various environmental factors before and after the construction of the breakwater. They revealed that differences in water temperature between the inside and outside of the bay were >3 °C at the surface and approximately 2 °C at a depth of 20 m. These results suggest that the construction of the breakwater caused stagnation of water inside the bay, thereby enhancing density stratification (Miyazawa and Hayakawa 1994). This stagnation of water may affect the nutrient concentration derived from terrestrial sources. Although there is no detailed information about the nutrient concentration of Ofunato Bay before the construction of the breakwater (before 1967), the concentration of total inorganic nitrogen at the inner reach of the bay increased 5.6 times, from 0.1 mg L−1 in 1979 to 0.56 mg L−1 in 1984 (Ofunato City Health and Environment Division 1986). The concentration of inorganic phosphorus at the same point was 0.05 mg L−1 in 1985, which was 5 times greater than that observed in 1981 (0.01 mg L−1). Higher concentration of nutrients may increase the phytoplankton biomass. Although there is no information about phytoplankton cell density or chlorophyll a concentration before the construction of the breakwater, water transparency inside the bay after the construction of the breakwater (approximately 2–8 m) was lower than that before the construction of the breakwater (approximately 5–12 m, Miyazawa and Hayakawa 1994).
Increase in phytoplankton biomass may induce sedimentation of carcasses at the bottom of the bay; furthermore, a series of feeding and excretion activities of cultivated shellfish promotes sedimentation of organic matter at the bottom of the bay. Terasawa et al. (1995) estimated that cultivated oysters consume organic matter equivalent to 1.24 tons C day−1 that is 30 % of the phytoplankton production in the entire Ofunato Bay. Moreover, they estimated the amount of organic matter flux at the bottom of the bay to be 1.05 tons C day−1. Decomposition of organic matter by bacteria could lead to the depletion of dissolved oxygen at the bottom layer. Kataoka et al. (1998) estimated that dissolved oxygen will decrease approximately 18–20 % in cases where cultivated oysters are present compared to cases where they are absent. The degree of saturation for dissolved oxygen at the bottom layer after the construction of the breakwater (approximately 17–60 % during 1967–1977) was lower than that before the construction of the breakwater (approximately 60–75 % during 1963–1967, Miyazawa and Hayakawa 1994). Depleted dissolved oxygen resulted in a confined and shallower oyster cultivation depth and a sequential decrease in the growth of cultivated oysters. The annual oyster production of Ofunato Bay decreased from 800 to 1000 tons before the construction of the breakwater (the latter 1950s to the former 1960s) to below 400 tons (after 1970s, Miyazawa and Hayakawa 1994). Thus, the construction of the breakwater changed not only the physical but also the biological and chemical environmental characteristics of Ofunato Bay, further leading to the decrease in oyster cultivation.
The breakwater was completely destroyed and shellfish mariculture rafts were wiped away by the huge tsunami triggered by the 2011 off the Pacific coast of Tohoku Earthquake (hereafter, the 2011 earthquake, Mimura et al. 2011, Kemuriyama 2011). To elucidate the influence of the tsunami on the aquatic environment of Ofunato Bay, field observations of tidal current, temperature, salinity, dissolved oxygen, and bottom sediment of the bay were conducted (Murakami et al. 2013, Furudoi et al. 2014). These studies revealed that stratification of seawater inside of the bay had been modified, and the intermittent intrusion of seawater outside of the bay was confirmed. The physical environment of Ofunato Bay has been investigated in detail after the 2011 earthquake, however, research on the impact of the earthquake and tsunami on biological and chemical environmental factors of the bay has been insufficient until now.
We investigated nutrients (nitrate + nitrite, ammonium, phosphate and silicate) and chlorophyll a concentrations, dissolved oxygen and heterotrophic bacteria abundance inside and outside Ofunato Bay from September 2012 to September 2014 after the 2011 earthquake. This study aimed to elucidate alterations in the biological, chemical, and environmental conditions, which were caused by the destruction of artificial construction and facilities because of the tsunami. These results were compared with the data reported by Hayakawa (1990) in 1980s, and the data that we collected from 2009 to 2010. The differences before and after the 2011 tsunami triggered by the earthquake were then discussed.
2 Materials and methods
Monthly sampling was conducted from the inside (St. 1, bottom depth of 32 m) and outside (St. 2, bottom depth of 38 m) Ofunato Bay from September 2012 to September 2014 (Fig. 1). Vertical profiles of temperature, salinity, and dissolved oxygen (DO) were measured using a RINKO Profiler (ASTD102, JFE Advantec Co., Ltd.). A DO sensor was calibrated by 2-point calibration with zero-solution produced by sodium sulfite (Na2SO3). Water samples for nutrients and chlorophyll a measurements were collected from every 5-m layer starting at the surface and continuing to a depth of 30 m using a Niskin water sampler. In the land laboratory, water samples for nutrient measurements were filtered using a DISMIC filter (pore size 0.45 µm, Advantec Co., Ltd.), transferred into 15-ml polypropylene tubes (Fast Gene Co., Ltd.), and frozen at −20 °C until measurement. Nutrient (nitrate + nitrite, ammonium, phosphate, and silicate) concentrations were determined using a QuAAtro2-HR auto analyzer (BLTEC Co., Ltd.) by the azo dye formation method with cadmium reduction for nitrate, indophenol method for ammonium, and the molybdenum blue method for both phosphate and silicate. Detection limits were 0.012 µM for nitrate + nitrite, 0.091 µM for ammonium, 0.028 µM for phosphate and 0.096 µM for silicate. Water samples for chlorophyll a measurement were filtered using a Whatman GF/F filter (pore size 0.7 µm), and chlorophyll pigments on the filter were immediately extracted using N,N-dimethylformamide (DMF) and frozen at −80 °C until measurement. Chlorophyll a concentration was measured using a Trilogy fluorometer (Turner Designs Co., Ltd.) based on the acidified method (Holm-Hansen et al. 1965). Samples for determination of heterotrophic bacteria were collected from the surface and at a depth of 10 m at St. 1 using a Niskin water sampler, transferred into 8-ml plastic vials, fixed with gultaraldehyde (final concentration = 1 %), and refrigerated (below 4 °C) until measurement. Heterotrophic bacteria were stained using 4′,6-diamidino-2-phenylindole (DAPI, final concentration = 0.5 µg ml−1) and filtered onto a black polycarbonate filter (pore size = 0.2 µm, Advantech Co. Ltd.). Ten fields per one filter sample were photographed using a digital camera (DP71, Olympus Co. Ltd.) mounted on an epifluorescence microscope (BX31, Olympus Co. Ltd.) under UV excitation. Heterotrophic bacteria cell numbers of all photographs (10 fields per one sample) were counted using ImageJ software (NIH, USA).
To compare the differences in water quality between before and after the 2011 earthquake, the monthly data average for 10 years (1979–1989) pertaining to the center areas (near St. 1) and the outside (near St. 2 in the present study) of Ofunato Bay reported by Hayakawa (1990) were used. Because these data were collected >25 years ago, we cited the most recent data for four nutrients and chlorophyll a concentration, dissolved oxygen and bacteria abundance collected from 2009 to 2010 by the authors pertaining to the center of the bay (near St. 1) for comparing with the present study data. Water samples for nutrients and chlorophyll a measurements were collected and treated in the same way for the present study. Nutrient concentration was analyzed using a TRAACS2000 auto-analyser (Bran + Luebbe Co. Ltd.). Chlorophyll a concentration was measured using a 10-AU fluorometer (Turner Designs Co., Ltd.) based on the acidified method. DO was measured using a AAQ-RINKO Profiler (AAQ121, JFE Advantec Co., Ltd.). Cell numbers of the heterotrophic bacteria were determined in the same way for the present study.
3 Results
3.1 Temperature and salinity
At both stations, the vertical temperature profiles were homogenous from October to April for two years, and a noticeable thermocline developed from May to August at a depth of 10 m from the surface (Fig. 2a). In September, high water temperatures (>20 °C) were at the surface continuing to a depth of 30 m. The coldest water appeared in spring (March and April). In particular, extremely cold waters (<3 °C) were observed in April 2014. The vertical differences in temperature between the surface to a depth of 10 m was greater at St. 1 (7.3 °C; May 2013) than that at St. 2 (5.0 °C; July 2014). Water temperatures measured at St. 1 and 2 were almost the same from September 2012 to March 2013 for both the surfaces at a depth of 10 m; however, higher temperatures at the surface and lower temperatures at a depth of 20 m at St. 1 than those at St. 2 were observed from May to August 2013. The horizontal difference in water temperature was greater at the surface (2.1–2.9 °C) than that at a depth of 20 m (0.1–1.2 °C). In March 2014, an anomalous higher temperature at a depth of 20 m at St. 1 than that at St. 2 was observed (Fig. 2a).

Monthly variations in vertical profiles of water temperature (a) and salinity (b) at Sts. 1 and 2
At the both sampling sites, salinities were >33.3 below a depth of 10 m, except for the period from March to April 2014 (Fig. 2b). In this period, low salinity (<33.0) was observed extending from the surface to the bottom layer. Extremely low salinity (<30.0) was observed at the surface of St. 1 from June to August (21.9 in July 2013 and 11.5 in June 2014). Similar to the inside of the bay (St. 1), the surface salinity at the outside of the bay (St. 2) also decreased to < 23.0 from June to August; however, a slight decrease in salinity (30–32) was observed from March to May.
3.2 Nutrients
As a general trend, nitrate + nitrite concentrations were high from winter to spring (December–April), and low during early summer (May and June, Fig. 3a). Pertaining to the outside of the bay (St. 2), nitrate + nitrite concentrations increased in the middle layer (at a depth of 10–20 m) during early spring (February and March, Fig. 3a). Pertaining to the inside of the bay (St. 1), extremely high concentrations (10 µM) were observed at the surface from June to July for both years (Fig. 3a).


Monthly variations in vertical profiles of nitrate + nitrite (a), ammonium (b), phosphate (c) and silicate (d) concentrations at Sts. 1 and 2
Vertical profiles of ammonium concentrations were homogenous from winter to spring at both sampling sites (Fig. 3b). Near the surface, seasonal change in ammonium concentration was almost similar between the outside (St. 2) and inside (St. 1) of the bay; they varied from 0.2 to 2.8 µM, except in December 2013 (4.3 µM, Fig. 3b). At the bottom layer, extremely high concentrations were observed from summer to autumn at St. 2 (4.7 µM in September 2012, 6.8 µM in September 2013, and 8.1 µM in July 2014, Fig. 3b). Compared to the inside of the bay, ammonium concentration at the bottom layer at St. 1 was remarkably lower from summer to autumn, ranging from 1.7 to 3.9 µM (Fig. 3b).
Seasonal and vertical variations in phosphate concentrations were similar to those in ammonium concentrations. Seasonal changes were slight at the surface, and remarkably higher values were seen from summer to autumn for the bottom layer at both sampling sites. (Figure 3c). At the outside of the bay (St. 2), high concentrations were observed between depths of 10–30 m during early spring (from March to April, 2013 and 2014). These results were similar to those of nitrate + nitrite concentrations at the same site (Fig. 3a, upper panel). At the inside of the bay (St. 1), remarkably high concentrations in the bottom layer were observed from September 2012 to 2014 (0.67–1.53 µM, Fig. 3c).
At both sampling sites, silicate concentrations tended to be high at the upper layer and low at the bottom layer, except for some sporadically high concentrations observed during September 2013 (Sts. 1 and 2), January 2014 (St. 2), and July 2014 (St. 1, Fig. 3d). During early spring, silicate concentrations near the surface at the outside of the bay (St. 2) were greater (24.9 µM in March 2013 and 11.0 µM in April 2014) than those at the inside of the bay (St. 1, 16.3 µM in the same months, Fig. 3d). During summer, extremely high concentrations were seen at the surface at St. 1 (30.0 µM in July 2012 and 36.6 µM in July 2014).
3.3 Chlorophyll a
Chlorophyll a concentrations showed similar seasonal changes at both sites (low concentrations continued from late autumn to late winter, increased in March, and peaked in April, Fig. 4a). At the outside of the bay (St. 2), chlorophyll a decreased in May and low concentrations (<3.6 µg L−1) continued until early next spring; however, at the inside of the bay (St. 2), chlorophyll a concentrations increased again in summer (from July to August). At both sampling sites, chlorophyll a concentrations in summer 2014 (from May to August) were greater (0.86–5.83 µg L−1 at St. 2 and 0.48–13.37 µg L−1 at St. 1) than those in 2013 (0.12–3.67 at St. 2 and 0.23–11.42 at St. 1).

Monthly variations in vertical profiles of chlorophyll a (a) and dissolved oxygen (b) concentrations at Sts. 1 and 2
3.4 Dissolved oxygen
At both sampling sites from November to March, dissolved oxygen (DO) was vertically homogenous, ranging from 5.5 to 7.5 ml L−1, then peaked (>8 ml L−1) in April and May at the surface layer (Fig. 4b). The DO stratifications were well developed from June to October, and the lowest DO was recorded in both September 2013 and 2014 at the bottom layer. The DO at the bottom layer in September 2012 was higher (4.6–4.8 ml L−1) than that during the same months in 2013 and 2014. The vertical mean DO in September 2012 (5.1–5.3) was also greater than that in 2013 and 2014 (Fig. 4b).
3.5 Heterotrophic bacteria abundance
Seasonal changes in heterotrophic bacteria abundance at depths of 0, 10, and 20 m at the inside of the bay (St. 1) are shown in Fig. 5. As a general trend, bacteria abundance was high at the surface and low at the deeper layers, increasing from spring to summer and decreasing during winter. At the surface, bacteria abundance was <0.6 × 106 cells ml−1 until July 2013, and increased to >1.1 × 106 cells ml−1 in August and September 2013. From spring to summer 2014, bacteria abundance was greater (1.2–1.4 × 106 cells ml−1) than that in the same seasons in 2013 (0.12–0.55 × 106 cells ml−1). A similar tendency was seen at depths of 10 and 20 m.

Monthly variations of the changes in horizontal profiles of heterotrophic bacteria abundance at depths of 0, 10, and 20 m at St. 1
4 Discussion
4.1 Possible influence of breakwater destruction on the circulation of Ofunato Bay
There are several studies that have investigated the impact of artificial structures on the coastal environment; however, there are few studies that have investigated biological and chemical environmental parameters in the event of destruction of artificial structures for any reason. In the present study, we discussed the effects of the destruction of the breakwater on the fisheries and environments of the coastal areas of Ofunato Bay.
Miyazawa and Hayakawa (1994) investigated the influence of the construction of the breakwater on the environment inside Ofunato Bay. They suggested that water exchange between inside and outside the bay subsided because of differences in water temperature between inside and outside the bay being greater in summer. After the breakwater destruction as a result of the tsunami, the differences in water temperature between the inside (St. 1) and outside (St. 2) of the bay in summer (June–September) were within 2 °C at the surface and 1.5 °C at a depth of 20 m (Fig. 6). These differences at the surface were less than those during the period of breakwater existence (1987–1992, >3 °C) and nearly the same as those before breakwater construction (1953–1964, within 2 °C, Miyazawa and Hayakawa 1994). Murakami et al. (2013) reported that a clear thermocline was observed as a boundary at the mouth of the breakwater (depth of 16 m) before the 2011 earthquake; however, the thermocline at the same depth after the 2011 earthquake had decreased (Murakami et al. 2013, cited therein). Based on these phenomena, it is suggested that exchange of seawater between inside and outside the bay became enhanced as a result of the destruction of the breakwater at the mouth of the bay.

Horizontal differences in water temperature between Sts. 1 and 2 during September 2012–September 2014. If the value is positive, the water temperature inside the bay (St. 1) was higher than that outside (St. 2)
4.2 Possible tsunami-induced changes in nutrient status of Ofunato Bay
In a closed water system, great fluctuations in environmental factors are often observed because of changes in weather, rainfall, and sequential inflow of river water. In the case of Ofunato Bay, inflow of river water from the Sakari River (Fig. 1) was estimated to be 3.3 m3 s−1 as an annual mean (Hayakawa 1986) with unusual fluctuations caused by occasional differences in weather conditions. According to the data before the 2011 earthquake, surface salinity fluctuated from 10.7 to 31, silicate varied from 8.1 to 106 µM, and nitrate + nitrite varied from 0.43 to 24.9 µM at the surface layer of the center of the bay (near St. 1) within one month (June 2009 at St. 1). In the present study, significant negative correlations were found between salinity and nitrate + nitrite and silicate concentrations at the surface of both sampling sites (St. 1 and 2, p ≤ 0.01), suggesting that nitrate + nitrite and silicate concentrations were strongly affected by the inflow of river water. To represent mean seasonal changes of the coastal environmental characteristics, it is necessary to calculate the average data obtained over multiple years to compensate for occasional anomalous events. Hayakawa (1990) calculated the mean seasonal changes of nutrients on the basis of monthly measurements for 10 years (1979–1989). These data appropriately represent the seasonal environmental changes of the bay before the 2011 earthquake, whereas the data exhibited in the present study (after the 2011 earthquake) were obtained over a period of only 2 years (from 2012 to 2014). Thus, a direct comparison between the data by Hayakawa (1990) and the present study is inappropriate when considering the impact of the tsunami on the bay environment. In the present study, the vertical profiles of salinity at a depth of <10 m were homogenous throughout the year, suggesting that there was less impact of river water inflow below a depth of 10 m. Therefore, we compared data obtained below a depth of 10 m before and after the 2011 earthquake to discuss the influence of the tsunami on the environment inside Ofunato Bay.
According to Hayakawa (1990), nitrate + nitrite concentrations were >5 µM from December to April, and then decreased to <1 µM from May to August at a depth of 10–15 m. Nitrate + nitrite concentrations then increased 2–5 µM at the center of the bay from October to December. Extremely high concentrations (>8 µM) were observed in the bottom layer in August. Similar seasonal changes of nitrate + nitrite concentrations were observed in the outside of the bay, however, high concentrations at the bottom layer during summer were not observed (Hayakawa 1990). Nitrate + nitrite concentration fluctuations before the 2011 earthquake were similar to those after the 2011 earthquake (Fig. 3a). There were no significant differences in nitrate + nitrite concentrations at depths of 10 and 20 m between before (2009–2010) and after (2012–2014) the 2011 earthquake (p > 0.1, paired t test, Fig. 7a; Table 1). This pattern was also observed in seasonal changes in the silicate concentration; it became low (<5 µM) at the middle layer (near a depth of 10 m) from spring to summer and high (>10 µM) from autumn to winter both before and after the 2011 earthquake (Hayakawa 1990, Fig. 7d). Significant differences between the periods before and after the 2011 earthquake were also not observed at depths of 10 and 20 m (both p > 0.7, paired t-test, Table 1; Fig. 7d). These results suggest that there were no significant effects on nitrate + nitrite and silicate concentrations inside Ofunato Bay as a result of the tsunami.


Comparisons of monthly variations in horizontal profiles of nitrate +nitrite (a), ammonium (b), phosphate (c) and silicate (d) concentrations at depth of 10 and 20 m in St. 1 between before (2009–2010) and after (2012–2014) the 2011 earthquake
Before the 2011 earthquake, ammonium concentrations at the center of the bay ranged from 3 to 5 µM below a depth of 10 m, except for the low concentration (<1 µM) that was found around a depth of 10 m from May to August (Hayakawa 1990). At the bottom layer, ammonium concentrations increased to >5 µM during July–October. The maximum value (15 µM) was observed in September. These values were much greater than those after the 2011 earthquake (Fig. 3b). There were high concentrations at the bottom layer in September 2013 and July 2014, however, the concentrations rose to 8.5 µM (Fig. 3b). Compared to those during 2009–2010, ammonium concentrations measured during 2012–2014 were significantly lower (p < 0.05, paired t-test, Table 1), particularly during summer to autumn at depths of 10 and 20 m (Fig. 7b). Regarding phosphate concentrations, large differences before and after the 2011 earthquake were observed. Before the 2011 earthquake, phosphate concentrations were >0.5 µM, except the concentrations <0.5 µM at the center of the bay during early summer to autumn below a depth of 10 m (Hayakawa 1990), and extremely high concentrations (>3 µM) were observed at the bottom layer in September. On the other hand, the peak value after the 2011 earthquake was only 1.5 µM (Fig. 3c), and throughout the survey period, significantly lower concentrations were seen when compared to those from 2009 to 2010 at depths of 10 and 20 m (p < 0.01, paired t-test, Table 1; Fig. 7c). In contrast to nitrate + nitrite and silica, ammonium and phosphate concentrations after the 2011 earthquake appeared to be affected. These features of nutrient status before the earthquake are considered to be related to the collapse of breakwater by a massive tsunami and consequential enhanced water exchange between inside and outside the bay. Yamamoto et al. (2015) investigated the concentration of nutrients in Kamaishi Bay, close-in Ofunato Bay, after the 2011 earthquake, and revealed that total inorganic nitrogen, phosphate, and silicate showed altered patterns of seasonal variation compared with before the earthquake. Kamaishi Bay, as well as Ofunato Bay, was also a semi-closed area because of the human-made breakwater effect at the entrance; however, the breakwater was destroyed by a massive tsunami. The alteration of nutrient features in Kamaishi Bay after the earthquake is attributed to the collapse of the breakwater (Yamamoto et al. 2015).
Changes of nutrient features in Tohoku coastal areas between before and after the 2011 earthquake were also investigated in Otsuchi Bay, north of Kamaishi Bay, by Fukuda et al. (2015). They revealed that higher concentrations of nitrite and silicate until September 2011, and higher phosphate concentration during the mixing period between November 2011 and January 2012 than those before the earthquake. However, continuous low concentrations of ammonium and phosphate, as observed in Ofunato Bay (Fig. 7b, c), were not recognized. In Otsuchi Bay, the width of the entrance of Otsuchi Bay is approximately 2 km, and the water residence time within the bay is less than a month (Fukuda et al. 2015). On the other hand, the width of the entrance of Ofunato Bay was 200 m before the earthquake. Hibino et al. (2004) simulated that 50 % of seawater at the bottom layer in Ofunato Bay was replaced within 50 days, suggested that water residence time was more than 3 months before the earthquake. Because of its open geographical structure and enhanced water exchange between inside and outside the bay, environmental modifications of the water column of Otsuchi Bay after the earthquake are considered to be moderate compared to those of Ofunato Bay.
Because the changes in concentrations before and after the 2011 earthquake differed depending on the type of nutrients, it is inappropriate to attribute the cause of the change in nutrients only to the destruction of the breakwater and consequential enhanced exchange of seawater between the inside and outside of the bay. In addition to the destruction of the breakwater, another important change in the ‘artificial’ environment of Ofunato Bay after the 2011 earthquake is the elimination of shellfish mariculture rafts. Before the construction of the breakwater (1955–1964), the number of mariculture rafts inside the bay ranged from 2200 to 2800, and the annual catch of cultivated oysters was 800–1100 tons (Miyazawa and Hayakawa 1994). After the construction of the breakwater (1970s), the number of mariculture rafts was nearly the same (2500–2800), however, the annual catch of oysters decreased to 400–500 tons. Subsequently, the oyster catch decreased below 300 tons during the 1980s. Since the 1990s, the annual oyster catch has remained at 300–400 tons until the early 2000s (Ofunato City Agriculture and Fisheries Division 2004). During the most recent period before the 2011 earthquake (2010), the number of mariculture rafts was 1572. Cultivated oysters consume a significant amount of phytoplankton and excrete biodeposits into the surrounding water. Terasawa et al. (1995) simulated that the cultivated oysters in Ofunato Bay grazed approximately 30 % of phytoplankton production (1.24 tonC d−1), and deposited about the same amount of organic matter (1.05 tonC d−1) into the seafloor using an ecosystem model. Hayakawa et al. (2001) measured the seasonal patterns of chlorophyll a concentration and sedimentation rates in Ofunato Bay, and revealed that high sedimentation rates were observed in March and September, corresponding to elevated chlorophyll a concentration. Thus, a grazing and excretion process of cultivated oysters effectively precipitated the suspended organic matter to the seabed. Hayakawa et al. (2001) also measured the seasonal changes of the fluxes of total nitrogen (TN) and phosphorus (TP) in the center of Ofunato Bay and revealed that these values peaked in September (290 mgN m−2 d−1 and 28 mgP m−2 d−1, respectively). However, there were no seasonal trends in elemental composition within the sediment, suggests that biodeposits from cultivated oysters are degraded rapidly at the sediment–water interface (Hayakawa et al. 2001). There were no detailed data about an elution amount of nutrient from biodeposits in Ofunato Bay, although, a release rate of nitrogen and phosphorus from the sediment was estimated to be 75.6 mgN m−2 d−1 and 24.8 mgP m−2 d−1, respectively, in Etajima Bay, and 53.7 mgN m−2 d−1 and 17.4 mgP m−2 d−1, respectively, in Hiroshima Bay (Yamamoto et al. 2010). Considering dense distribution of mariculture rafts and semi-closed environment of Ofunato Bay before the 2011 earthquake, substantial amounts of ammonium and phosphate could have been eluted from biodeposits.
Shellfish mariculture rafts in Ofunato Bay were completely eliminated by the tsunami generated by the 2011 earthquake; however, 1172 new rafts were reconstituted by the end of 2012 or 2013 (Ofunato Fisheries Cooperatives, personal communication). As a simple comparison, the oyster mariculture scale after the 2011 earthquake was 74.5 % of that observed just before the earthquake (2010), and approximately 50 % of that observed in the 1980s (as compared by the number of the mariculture rafts). Reduction of shellfish mariculture scale is considered to have reduced the sedimentation of biodeposits and consequential elution of ammonium and phosphate. Quantitative data about biodeposits from cultured shellfish and consequential elution of nutrients after the 2011 earthquake are not available, however, the density or abundance of heterotrophic bacteria was significantly reduced (p < 0.01, paired t-test, Table 1; Fig. 8c), and dissolved oxygen was significantly increased (p < 0.01, paired t-test, Table 1, Fig. 8b) as compared to that before the 2011 earthquake (2009–2010). Increased dissolved oxygen and decreased heterotrophic bacteria abundance and ammonium and phosphate concentrations could be induced by the possible reduction in biodeposits from cultured shellfish.

Comparisons of monthly variations in horizontal profiles of chlorophyll a (a) and dissolved oxygen (b) concentrations, and heterotrophic bacteria abundance (c) at depth of 10 and 20 m in St. 1 between before (2009–2010) and after (2012–2014) the 2011 earthquake
Although possible remineralization of nutrients by bacteria reduced after the 2011 earthquake, the chlorophyll a concentration at the center of the bay at depths of 10 and 20 m increased significantly (p < 0.01, paired t test, Table 1; Fig. 8a) as compared to that before the earthquake (2009–2010). Chlorophyll a concentrations are affected by filter feeding of shellfish. Kataoka et al. (1998) estimated that the filter feeding of cultivated oysters resulted in a 20–30 % decrease in the phytoplankton biomass of Ofunato Bay. Thus, it is suggested that the decrease in cultivated oyster biomass after the 2011 earthquake reduced feeding pressure on phytoplankton inside the bay, resulting in an increase in phytoplankton biomass. Abundant phytoplankton consumes substantial nutrients, which may have contributed to the decreased nutrient concentrations after the 2011 earthquake, together with the reduced remineralization.
In summary, the destruction of the breakwater at the entrance of Ofunato Bay promoted enhanced circulation of seawater between inside and outside the bay, decreasing the nutrient concentrations inside the bay. The reduction of the oyster mariculture scales diminished the sedimentation of organic matter, resulting in reduction of nutrients (particularly ammonium and phosphate) released from organic sediments. Furthermore, the decrease of feeding pressure of cultivated oysters on phytoplankton cause an increase in the phytoplankton biomass, resulting in greater nutrient consumption. The degree of oyster growth is largely related to the chlorophyll a concentration (the phytoplankton biomass, Miyazawa and Hayakawa 1994), thus, compared to that before the 2011 earthquake, the environmental condition after the earthquake was considered to be favorable for oyster mariculture.
The breakwater reconstruction began in 2012 and will be completed in 2016. The new breakwater has a seawater exchange function between inside and outside the bay to keep the water quality of the bay in good condition (Okawa and Higashiyama 2013, Iwate Prefectural Land Development Section 2015), however, an effect of the new breakwater on environmental quality is not clear enough. Therefore, continuous environmental monitoring and detailed field studies are required to evaluate the effects of the reconstruction of the breakwater on the coastal environment of Ofunato Bay.
References
Fukuda H, Katayama R, Yang Y, Takasu H, Nishibe Y, Tsuda A, Nagata T (2015) Nutrient status of Otsuchi Bay (northeastern Japan) following the 2011 off the Pacific coast fo Tohoku Earthquake. J Oceanogr. doi:10.1007/s10872-015-0296-2
Furudoi K, Sakai S, Murakami K, Nakamura Y, Abe I, Okada T, Shibaki H, Takao T (2014) Analysis of influence factors to the water environment of Ofunato Bay based on the observed data. J Japan Soc Civil Engineers, Ser B2 70:426–430. doi: 10.2208/kaigan.70.I_426 (in Japanese with English abstract)
Hayakawa Y (1990) Mean seasonal changes of dissolved inorganic nutrients in the Ofunato estuary. Nippon Suisan Gakkaishi 56:1717–1729
Hayakawa Y, Kobayashi M, Izawa M (2001) Sedimentation flux from mariculture of oyster (Crassostrea gigas) in Ofunato estuary, Japan. ICES J Mar Sci 58:435–444
Hibino T, Tsuruya H, Itabashi N (2004) Seasonal variation characteristics of water structure and quantitative analysis of seawater exchange in Ofunato Bay. J Japan Soc Civil Engineers, Ser 761:69–80. doi: 10.2208/jscej.2004.761_69 (in Japanese with English abstract)
Holm-Hansen O, Lorenzen C, Holmes R, Strickland JDH (1965) Fluorometric determination of chlorophyll. J du Cons 30:3–15
Iwate Prefectural Land Development Section (2015) Overview of prefectural land development administration, 2015 version. Iwate prefecture Home page. Download from http://www.pref.iwate.jp/dbps_data/_material_/_files/000/000/015/349/h27kendogyousei.pdf (in Japanese)
Kataoka S (1998) About the current status and reproducibility of flow regime and water quality, and the role of aquaculture oyster. J Ocean Eng 45:481–485 (in Japanese)
Kemuriyama A (2011) The damage done to fisheries in Iwate Prefecture by the Great East Japan Earthquake. Nippon Suisan Gakkaishi 77:712–714. doi:10.2331/suisan.77.712 (in Japanese)
Mimura N, Yasuhara K, Kawagoe S, Yokoi H, Kazama S (2011) Damage from the Great East Japan Earthquake and Tsunami—a quick report. Mitig Adapt Strateg Glob Chang 16:803–818. doi:10.1007/s11027-011-9297-7
Miyazawa K, Hayakawa Y (1994) A large-scale structure and the environmental capacity for aquaculture—effects of and artificial structure (breakwater at the entrance of the Ofunato estuary) on aquaculture and water qualities in the estuary. Bull Coast Oceanogr 32:29–38 (in Japanese with English abstract)
Murakami A, Sakai S, Murakami K, Nakamura Y, Okada T, Takao T, Shibaki H (2013) Change in water and sediment quality in Ohfunato Bay induced by the 2011 Great East Japan Tsunami. J Japan Soc Civil Engineers, Ser B2 69:496–500. doi:10.2208/kaigan.69.I_496 (in Japanese with English abstract)
Ofunato City Agriculture and Fisheries Division (2004) Fisheries promotion plan in Ofunato City. Ofunato City Home page. Download from http://www.city.ofunato.iwate.jp/www/contents/1342083442949/files/suisan_keikaku.pdf (in Japanese)
Ofunato City Health and Environment Division (1986) The white book on environmental pollution of Ofunato City, 1986 version. p 84 (in Japanese)
Okawa M, Higashiyama K (2013) An examination of water passing hole in the underwater bank of opening part of the breakwater at the entrance of the Ofunato Bay. Rep Coast Dev Inst Tech 13:5–8 (in Japanese)
Terasawa T, Nakata K, Taguchi K (1995) Numerical modeling on the Ofunato Bay ecosystem including the oyster farming. In: Proceedings of 4th International Conference on Estuarine and costal modeling, ASCE, pp 530–541
Yamamoto T, Date E, Oda S (2010) Characteristics of sediment in Etajima Bay and influences of oyster cultivation on sediment. Bull Hiroshima Pref Tech Res Inst Public Health Environ Center 18:7–13 (in Japanese with English abstract)
Yamamoto M, Kato T, Tabetata S, Kitazawa D, Fujino M, Shozugawa K, Matsuo M, Tanaka K, Michida Y (2015) Changes in the coastal environment in Kamaishhi Bay after the Great East Japan Earthquake. Nippon Suisan Gakkaishi 81:243–255. doi:10.2331/suisan.81.243 (in Japanese with English abstract)
Acknowledgments
We are grateful to Y. Azuma and Ofunato Fisheries Cooperatives for their help in field sampling. This study was supported by the research program “Tohoku Ecosystem-Associated Marine Science” from the Ministry of Education, Culture, Sports, Science and Technology in Japan.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Yamada, Y., Kaga, S., Kaga, Y. et al. Changes of seawater quality in Ofunto Bay, Iwate, after the 2011 off the Pacific coast of Tohoku Earthquake. J Oceanogr 73, 11–24 (2017). https://doi.org/10.1007/s10872-015-0336-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10872-015-0336-y