
Abstract
The Eastern North American monarch butterfly population has severely declined over the past decade. The decreasing availability of larval host plants (milkweeds) due to the use of herbicide-tolerant crops has been implicated in this decline. Roadsides could provide additional habitat for monarchs. In this study we document the occurrence of milkweed and monarchs on roadsides, and discuss whether roadsides are appropriate targets for monarch habitat restoration. We sampled roadside rights-of-way in the Upper Midwestern U.S. during the summer of 2015 to estimate the abundance, distribution, and diversity of milkweeds and the extent to which monarchs use these milkweeds. We then compared monarch densities in roadsides to other habitat types and modelled immature monarch densities based on several site characteristics. Our findings suggest that roadsides have conservation potential for monarchs, especially when other habitat is scarce and if wildlife-friendly management practices are enacted. Milkweeds were found on ~60% of roadside transects. Asclepias syriaca was the most common of the seven species encountered, occurring on 97% of transects with milkweed. Immature monarchs were observed in roadsides, but in lower densities than other habitats during the same time period. At lower milkweed densities, immature monarch density per unit area is positively correlated with milkweed density. However, milkweed density weakens as a predictor of immature monarch density over ~0.6 plants per m2, possibly indicating a saturation effect.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The recent decline of the eastern migratory North American monarch butterfly (Danaus plexippus) population has been well documented. Annual population estimates in the overwintering sites in Mexico reveal a decline in the area occupied by monarchs over the past decade (Brower et al. 2012; Vidal and Rendón-Salinas 2014), albeit with a promising uptick in the winter of 2015–2016 (World Wildlife Fund-Mexico 2016). Additionally, measurements during the breeding season in the Northern U.S. indicate that the density of monarch eggs per milkweed stem has declined since 2006 (Stenoien et al. 2015). Overlaid on this decline is a great deal of year to year variation that appears to be weather-driven (Zipkin et al. 2012; Saunders et al. 2016). While many factors could be contributing to declining monarch numbers, there is strong evidence that the loss of the host plant resource in the Upper Midwestern U.S. (Midwest) has been especially detrimental (Brower et al. 2012; Pleasants and Oberhauser 2013; Flockhart et al. 2015; Stenoien et al. 2016).
Monarch larvae consume plants in the genus Asclepias (and a few closely related genera), commonly known as milkweeds. For much of the twentieth century, the vast majority of monarchs produced in the eastern U.S. likely originated from milkweed growing in agricultural fields, primarily corn (Zea mays) and soy (Glycine max) (Oberhauser et al. 2001). However, since the introduction of genetically modified herbicide-tolerant row crops (corn and soybeans), milkweed has virtually disappeared from within these fields (Hartzler and Buhler 2000; Hartzler 2010; Pleasants and Oberhauser 2013; Pleasants 2016). Pleasants (2016) estimates a 40% decline in milkweed present to the Midwest landscape from 1999 to 2014. The majority of these milkweeds were lost from agricultural fields, which average 3.9 times more monarch eggs than milkweeds in non-agricultural habitats, thus the potential monarch support capacity loss has been 71% (Pleasants and Oberhauser 2013; Pleasants 2016). Projection models estimate that milkweed abundance will continue to decrease by another 14% within a century, a slower decline than has been experienced to date, but worrisome enough to predict a monarch quasi-extinction (<1000 individuals) probability greater than 5% within the same timeframe (Flockhart et al. 2015). A more recent population viability analysis (Semmens et al. 2016) suggests that the current risk of reaching a population size that would occupy 0.15 ha at the Mexican wintering sites, a value less than a quarter of the previous low, in the next 10 years is 30%.
Due to their collective acreage, roadsides could provide an important source of monarch habitat if managed properly (Ries et al. 2001; Mueller and Baum 2014; Hopwood et al. 2015). They comprise over 10 million acres of land in the U.S. (Forman et al. 2003) and in many states they are the largest holdings of public land (Hopwood et al. 2015). Past studies highlight benefits that roadsides can have on survival, reproduction, and migration for various butterfly species in Iowa (Ries et al. 2001; Hopwood et al. 2015), especially when planted with appropriate host plants and nectar plants. In particular, studies that have looked specifically at monarchs suggest that their populations would benefit from increased abundance and connectivity of milkweed habitat (Zalucki and Kitching 1982; Zalucki and Lammers 2010; Zalucki et al. 2015). In highly developed areas such as agricultural or urban settings, roadsides may provide the only semi-natural habitat available for milkweeds. In fact, Flockhart et al. (2015) estimated that roadsides account for 10% of the remaining milkweed in the central U.S. region. Hartzler (2010) documented a 15% increase in milkweed abundance in Iowa roadsides between 1999 and 2009, perhaps due to a decreased herbicide use in roadside rights-of-way as part of an Integrated Roadside Vegetation Management (IRVM) program started in 1988.
Here, we assess the abundance of milkweed plants along roadside rights-of-way in the Midwest, a region that appears to contain the most important natal grounds for eastern migratory North American monarchs (Malcolm et al. 1993; Wassenaar and Hobson 1998), although these estimates were made before the advent of herbicide-tolerant row crops, and thus the relative importance of the area to monarchs could have declined as milkweed has disappeared from agricultural fields. Despite research on declining milkweed in agricultural habitats (Pleasants and Oberhauser 2013), there is a lack of comprehensive data on the abundance, diversity, and density of milkweed in roadside rights-of-way. If roadsides represent a significant source of habitat, these data could inform ongoing conservation priorities and practices. We also assess the extent to which monarchs use roadside habitat for reproduction and larval development by comparing the productivity of roadsides to that of other habitat types using citizen science data from the Monarch Larva Monitoring Project (2016).
Methods
Site selection
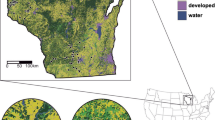
We randomly selected 212 roadside sites within a 250 mile radius of Minneapolis, Minnesota, an area that included portions of Minnesota, Wisconsin, South Dakota, and Iowa (Fig. 1). Large urban and heavily forested locations (including most of northeastern Minnesota and northern Wisconsin) were excluded from the sampling area because few milkweeds grow in these ecosystems. Each day, we selected groups of sites to sample that minimized distance traveled and maximized the number of sites visited. Driving direction also changed each week in order to sample all areas using a temporally balanced method (this was important for studying monarch phenology). Surveys were conducted from July 13th to October 15th, 2015. Within each site, we sampled for milkweed species richness, milkweed density, monarch egg and larval density, and adult monarch presence.

Locations of 1045 sampled transects. Each cluster of points represents five transects at a randomly-selected site
Sampling technique
We used a modified interrupted belt transect sampling method. Throughout this paper, “sites” refer to the initial randomly generated points and associated five-mile stretches of road, and “transects” refer to each of the five associated surveys at 1 mile intervals from the initial point. In a few cases, the 5 mile-long sites overlapped; when this occurred, both were sampled since the 50 m transects (spaced 1 mile apart) never overlapped. Upon arrival at each site, we first flipped a coin to choose a side of the road to sample, then ran the transect line from the front of the car in the direction it was facing. We measured five transects at 1 mile intervals (stop, survey, drive 1 mile, stop, survey, etc.); each transect was 50 m long parallel to the road, and the width extended from the road edge (where vegetation began growing) to an obvious outer edge barrier (fence, agricultural field, wooded area, lawn edge, etc.). For transects adjacent to “grassland” (see Table 1), there was often no clear delineation of where the roadside ended. In such cases, we measured roadside width to telephone or electrical poles adjacent to the road. At roadsides adjacent to “forest,” we measured width to where mature trees were growing. Thus, the sampling width included the non-forest roadside patch as well as 1–3 m into the wooded area. Plants in these margins were treated equally in our analysis. Roadside width was used to calculate area sampled and milkweed density (plants/m2), and was also used in models predicting densities of monarch eggs and larvae. GPS coordinates were recorded at a single point on the transect, designated at the north, east, northeast, or southeast end of each transect.
Roadside description
We recorded the land use/cover of the land adjacent to each roadside transect (Table 1). Adjacent landcover was not recorded for 13 sites in the beginning of our study. These sites were revisited using Google Maps (Google Maps 2016), and all were corn, soy, or forest. Corn and soy were combined into one category because these crops are often grown on a rotational basis. Adjacent land types found on fewer than ten transects (e.g. tomato field, tree farm, lettuce field) were combined into an “Other” category. If a transect spanned two different roadside types, its label reflected the land use that covered most of the transect. Some of the non-“Town or city” or non-“Lawn” sites included an area that was mowed, such as crop margins. To account for this, we recorded if roadsides had been recently mowed (within 2–3 weeks) at the time of observation. Transect locations that were not safely accessible (e.g. a cliff, area under a bridge, or on an interstate), were not sampled.
Milkweed sampling
We recorded the total number of milkweed stems in each transect, referred to from here on as “plants.” A milkweed stem was counted as a plant if soil separated stems of the same species. While multiple stems of several species, such as A. syriaca and A. verticillata may be ramets from a single plant (genet), this was impossible to distinguish using our sampling method. The exception was A. tuberosa, for which we counted stem clusters that were clearly part of a single plant, and separately recorded the number of stems observed. A. verticillata occurred in high densities that made individual stem counting difficult. Thus, A. verticillata counts were estimated by counting individual stems in two randomly selected 1 m2 quadrats within the larger patch, and multiplying the mean count for these quadrats by the area of the patch. Our sampling timeframe captured milkweed once it was fully vegetated, and ended when A. syriaca was mostly senesced throughout the Midwest.
Monarch presence
We monitored the first 15 plants encountered, then every additional fifth plant, for the presence of monarch eggs and larvae (identified to stadium). Transects wider than 10 m were sampled by walking from road to boundary at 5 m intervals. This allowed us to monitor a smaller swath of vegetation at a time, and still cover the full transect. The number of adult monarchs observed during each transect sampling period was also recorded.
To assess whether roadside milkweeds were used by monarchs at the same rate as plants in other habitats, we compared egg and larval densities recorded in this study to densities measured by Monarch Larva Monitoring Project (MLMP) volunteers within the same spatial and temporal boundaries. For more detail on these citizen science monitoring procedures, see Prysby and Oberhauser (2004). Briefly, MLMP volunteers monitor sites with milkweed on a weekly basis, counting the number of eggs and larvae (identified to stadium) observed on a known number of milkweed plants. Their sites include backyard gardens, natural areas (such as state parks and other nature preserves), pastures, old fields, and other areas that contain milkweed. Thus, we compared our roadside data to all of the data collected by MLMP volunteers, with exceptions described below.
All MLMP data were taken from within the same latitudinal and longitudinal bounds as the area we sampled. We then systematically excluded MLMP data that were likely to be inaccurate or unrepresentative of the landscape. MLMP data were excluded if: (1) they were from garden sites, because gardens tend to have higher per plant monarch densities than other site types (Stenoien et al. 2015) (30.5% of sites were excluded by this criterion); (2) they were from roadside sites, because we wanted to compare our randomly selected roadsides to other types of habitats (6% of sites were excluded by this criterion); (3) volunteers observed unrealistic ratios of eggs to larvae throughout the entire season, indicating that they may not have been able to accurately discern monarch eggs or that they selectively monitored plants on which they saw late instar larvae, which are often more visible from a distance (4.2% of sites were excluded by these criteria). Sites monitored fewer than four times were not excluded based on criterion three because seasonal fluctuations in immature monarch density mean that it is common to find more late-instar larvae than early instar larvae or eggs during some parts of the summer with infrequent monitoring. However, given that relatively few sites were excluded on the basis of ratios of eggs to larvae in sites monitored more frequently, we assumed these volunteers monitored accurately. A total of 239 MLMP sites (37.8% of a possible 633) were excluded because of one or a combination of the above criteria. The final MLMP dataset included 394 sites.
We used t tests to compare egg and larval per plant densities (combining all larval instars into a single category) between roadside transects and MLMP sites in Minnesota, Iowa, Wisconsin, and North Dakota during the same week. For this analysis, each MLMP site visit or roadside transect was treated as a single monitoring event.
To assess the effect of adjacent habitat type on immature monarch presence we used linear models and linear mixed effects models implemented via the lme4 package (Bates et al. 2015a) in R 3.2.2 (R Core Team 2014). Two models were created to assess the effects of several site characteristics on the observed densities of immature monarchs. These models differed in that one predicted monarchs per m2 and the other predicted monarchs per plant. Monarchs per m2 was calculated as follows:
where M = the number of immature monarchs found on the transect, p = the number of plants checked for monarchs, P = the total plants on the transect, and A = the area of the transect. Monarchs per plant was calculated as \(\frac{M}{p}\), the same metric used by the MLMP to measure immature monarch density (Oberhauser and Prysby 2008; Monarch Larva Monitoring Project 2016). We used backwards model selection based on Akaike Information Criterion (AIC) values (Akaike 2011) to select the best fitting model that still included adjacent habitat as a predictor for immature monarch presence. Our initial model included adjacent habitat type, day of year, roadside width, and milkweed density as linear fixed effects; and site ID as a random effect. The lme4 package was initially used, but if site ID was excluded, we used the lm function (base R) for the remainder of the model selection. Milkweed density was treated as a second order polynomial variable because when plotted on a scatterplot against immature monarch density, the relationship appeared to be curvilinear (non-linear, but displaying a smooth data trend). We used ggplot2 (Wickham 2009) in R3.2.2 (R Core Team 2014) to construct this plot. These data were fitted with a loess line because this fit our data best. The line was used to visualize what seemed to be a nonlinear relationship between milkweed and egg density without any a priori hypotheses, and was not used to extrapolate any further conclusions. The span parameter in this figure determines the curve’s sensitivity to variation in the data. The span we used gave the smoothest line while still fitting the data. While monarch densities in this region tend to have two peaks across the summer (Stenoien et al. 2015), our sampling period started at approximately the time of the second peak, after which monarch egg and larvae densities begin to decline as the population switches from breeding to migratory behavior. We only included sites with either A. syriaca or A. incarnata in our models; other milkweed species were rarely observed (Table 2), and more difficult to monitor accurately. Roadsides that had been recently hayed, or where vegetation was taller/thicker made species like A. verticillata more difficult to detect given its smaller structure. Additionally, the difference in biomass is a confounding variable for modelling when density is measured in plants/m2. To account for this, we did not include A. verticillata density in our linear models in which milkweed density was included as a predictor of monarch density.
We used Chi square tests to determine if the presence of adult monarchs on a transect was associated with milkweed presence. We did this for (1) all transects up to August 20 (when breeding monarchs are likely to be present, based on egg presence), and (2) all transects observed from August 20 to September 21 (when primarily non-breeding, migratory monarchs are expected to be present) (Stenoien et al. 2015).
Results
Sites sampled
We sampled 1045 roadside transects. Corn/soy was the most common roadside type observed (Table 1), with 250 transects adjacent to corn fields (23.5% of total), 158 adjacent to soybean fields (14.9%), and 41 adjacent to a harvested field that had been corn or soy (3.86%). Roadside width ranged from 2 to 37.2 m, with 68% of roadsides between 5 and 15 m wide, and an overall mean of 12.47 m (SE = ± 0.204). Roadside width varied among the categories for adjacent land type (F10, 1051 = 114.8, P < 0.0001), but the means of all categories were within 5 m of each other (Table 1).
Milkweed species
We found at least one milkweed plant on 622 of the 1045 transects surveyed (59.5%). We observed seven different milkweed species, but the majority of the plants were A. syriaca, which occurred in 57.8% of surveyed transects and 97.1% of transects with milkweed (Table 2). When milkweed was present on a transect, density appeared to vary between species, with A. verticillata and A. ovalifolia growing at higher mean densities. We did not test the significance of these density differences due to small sample sizes; only one transect contained A. ovalifolia and 28 had A. verticillata. There was no difference in milkweed density between different roadside types (F7, 800 = 0.112, P = 0.998 [excluding sites with A. verticillata]).
Because A. syriaca was the dominant milkweed in roadsides surveyed, we present summary statistics on its density. When present on a transect, A. syriaca density ranged from 0.000809 to 5.235 plants/m2 (Fig. 2, n = 595, mean = 0.0884, SE = ± 0.0118, median = 0.027). The median A. syriaca density for all transects, including those without milkweed, was 0.0036 plants/m2 (n = 1,062, mean = 0.0508, SE = ± 0.00681), and 0.011 plants/m2 (n = 838, mean = 0.0650, SE = ± 0.00887) for all transects with potential habitat (excluding lawn and town/city).

Frequency distribution of A. syriaca densities, when present on a transect
We surveyed three transects with A. syriaca milkweed densities greater than 5 SDs from the mean for all transects. These were considered outliers, and were removed from further analysis. These densities were: 5.24 plants/m2 (23.8 SDs from mean), 3.56 plants/m2 (15.8 SDs from mean), 1.37 plants/m2 (5.92 SDs from mean). All other densities observed were within 4 SDs of the mean.
Monarch density and presence
Monarch eggs and larvae were observed during every week from July 13th to September 9th (Fig. 3). We continued sampling for monarchs past this date, in case fresh milkweed growth promoted by mowing resulted in late-season laying, but no more eggs were found. Using September 9th as a cut-off, monarch eggs and larvae were found on 23.5% of our surveys on transects with milkweed plants (91 of 387 sites). Combining all observations from the weeks of July 13th through September 7th, roadside sites had significantly lower mean egg and larval per plant densities than MLMP sites monitored in the same area over the same time (eggs/plant: MLMP mean = 0.144, n = 222, Roadside mean = 0.0391, n = 399, t447 = −6.15, P < 0.00001, larvae/plant: MLMP mean = 0.0596, n = 222, Roadside mean = 0.0199, n = 399, t391 = −4.81, P < 0.0001) (Fig. 3). Comparisons between individual weeks when monarchs were observed revealed that roadside egg densities were significantly lower than MLMP eggs densities for 5 of the 9 weeks, and roadside larval densities were significantly lower than MLMP larval densities for 4 of the 9 weeks (Fig. 3).

Immature monarch density summed for all roadside and all MLMP sites in the sampling area that were observed during that week (comparisons made using each site as an individual observation). Roadside data are the darker shade on the left, MLMP data are the lighter shade on the right. Hash symbol indicates that both eggs and larvae were significantly different between MLMP and roadside data. Asterisk indicates that only eggs were significantly different between MLMP and roadside data
Our backwards model selection started using the lme4 package (Bates et al. 2015b), where site ID was treated as a random variable. Site ID was eliminated based on AIC value and then we switched to lm for the remainder of the model selection process. Following the full backwards selection, roadside width and site ID were excluded from the final model testing factors that affect immature monarch density on a per area basis (Table 3a). Monarchs per m2 decreased with day of year as the population shifted from breeding to migratory behaviors. Sites adjacent to forest had significantly higher and sites adjacent to corn/soy had marginally higher densities of immature monarchs than sites adjacent to grassland. Monarchs per m2 was positively affected by milkweed density at low and moderate milkweed densities, but at higher densities, the overall effect weakened or became negative, as evidenced by the negative coefficient for squared milkweed density. These coefficients were used to generate a point estimate of milkweed density at which the density of immature monarchs is maximized. This point estimate was determined to be 1.40 milkweeds/m2. Furthermore, when considered in isolation of other predictors, milkweed density appears to have a non-linear relationship with immature monarch density per unit area (Fig. 4). At low and medium milkweed densities (0 to ~0.6 milkweed/m2) immature monarch density increased linearly with milkweed density, but as milkweed densities increased above ~0.6 milkweed/m2, immature monarch density remained constant or decreased (although we observed few sites with milkweed densities at the high end of the range we modeled).

Plot of immature monarch density per m2.vs. milkweed density per m2. Each dot represents a single observation, with darker dots representing multiple observations in that data range. The grey area outlines the 95% confidence interval for the trend line (shown in black) (Span parameter = 0.75)
Backward selection of a model predicting immature monarchs per plant (Table 3b) excluded roadside width, site ID, and milkweed density as predictors (we used the same combination of lme4 and lm as described above). This model showed that sites adjacent to forests had more immature monarchs per plant, and that monarch per plant density decreased with day of year, but no other predictors were statistically significant.
The presence of adult monarchs on roadsides was associated with the presence of milkweed during the breeding period, but not during the migratory period. From July 20 to August 20, adult monarchs were more likely to be observed in roadside transects with milkweed. Adults were seen in 49 of 240 sites with milkweed, and 6 of 147 sites without milkweed (\(\chi _{1}^{2}=19.95\), d.f. = 1, n = 387, P < 0.0001) This was not the case from August 20 to Sept 21, when adults were seen in 16 of 204 sites with milkweed, and 6 of 131 sites without milkweed; (\(\chi _{1}^{2}=1.38\), n = 335, P = 0.239).
Discussion
Monarchs use milkweed growing along roadsides as breeding habitat. The distribution and abundance of milkweeds in our randomly surveyed sites suggests that these habitats are important for monarchs, but that roadside milkweeds have lower per plant use than milkweeds in other habitats. Further augmentation and management practices that favor the continued presence of milkweeds and other desirable habitat features such as nectar plants are likely to benefit monarchs.
Milkweed occurred on 59.5% of roadsides within our sampling area. We found seven species, but only A. syriaca was frequently observed. Three species (A. exaltata, A. ovalifolia, A. sullivantii) were found at only one site each, and three (A. verticillata, A. incarnata, A. tuberosa) at fewer than 30 sites each. It should be noted that A. sullivantii is considered threatened in Minnesota (USDA 2016). The success of A. syriaca in roadside sites relative to other species suggests that it may be best suited to survival in roadsides, and thus a good candidate for inclusion in restoration efforts aimed at adding milkweed plants into roadside vegetation. However, the fact that it is already present in most roadside transects, at least in the Midwest, suggests that consideration be given to effective management of existing resources rather than planting new roadside habitat. For example, roadsides already containing milkweed could be mowed at times that limit monarch egg and larval mortality, and the addition of nectar sources should be considered. A study in Germany and Switzerland concluded that there was a positive correlation between nectar plant abundance and increased fecundity in the map butterfly (Araschnia levana) (Mevi-Schutz and Erhardt 2005). Further research is needed to determine how monarchs respond to increased nectar plant abundance in restored habitats.
Although we did find comparatively high overall densities of A. verticillata, these were skewed by very high densities in a few scattered patches. This finding suggests that A. verticillata can do well on roadsides, and may be an appropriate species to consider in new plantings. Asclepias ovalifolia grew at a very high density in the one site in which we found it, so it may also be a candidate worthy of further consideration for roadside restorations. However, these species could have been found at few sites because they have highly specific habitat requirements.
The density of milkweed plants varied substantially across the sampled transects, both within and between species. For example, at one transect we found 1231 A. syriaca plants, and on the very next transect in the site (one mile down the road), we found zero plants in what appeared to be equally suitable habitat. The high variance in milkweed density, illustrated in Table 3, reflects its patchy distribution across the landscape.
The variability of milkweed presence and density calls for a multi-faceted approach to improving roadside habitat for monarchs. Roadsides that contain milkweed should be managed to benefit monarch presence (mowing schedule, inclusion of nectar plants, minimized risk of pesticide exposure). In areas devoid of milkweed, milkweed should be included in seed mixes or plug planting, and then managed accordingly.
We recorded if sites we surveyed had been recently mowed, but because we only visited sites once, these data do not include information on roadside mowing frequency or succession after mowing events. As such, we do not report on the frequency with which our sites had been recently mowed. Studies on the effect of early-season mowing in New York, and the Midwest have found that mowing conducted around July 1st or July 24th actually sustained suitable egg-laying habitat for longer than naturally grown habitat (Baum and Mueller 2015; Fischer et al. 2015). Female monarchs preferentially oviposit on fresh milkweed growth, which occurs after mowing, but it is not yet clear whether the number of juveniles lost to mowing may be offset by increased egg densities due to enhanced egg-laying (Fischer et al. 2015). Further research is needed to determine how effective Integrated Roadside Vegetation Management (IRVM) programs can be towards improving roadside habitat quality for monarchs.
Sites monitored by MLMP volunteers had significantly higher per plant monarch densities than roadside habitats throughout the summer. However, this does not mean that roadsides are not an important source of habitat for monarchs. The presence of 5th instar larvae on roadside milkweed plants indicates that monarch caterpillars are able to develop within the confines of roadsides. In fact, immature monarchs were significantly more abundant on roadsides adjacent to forest, with a trend toward higher monarch abundances adjacent to corn/soy sites. These two land cover categories are likely to have less monarch habitat available than many of the other categories we measured, which suggests that monarchs use roadsides more when other habitat is not available. Testing this possibility will require more detailed landscape-level analyses and a site selection process designed to include more equal representation of adjacent land categories.
Although our research was not focused on adult monarch behavior, during the time that egg laying took place, there was a significant positive correlation between adult occurrence and milkweed presence. This correlation disappeared in the late summer/early fall; in fact, we saw few monarchs along roadsides while monitoring during the fall migration. The fact that adult monarchs are more likely to be found in habitats containing milkweeds than other habitats during the breeding period, suggests that breeding monarchs are able to locate isolated milkweed patches in roadsides. It should be noted that we spent more time in transects with milkweed, and did not control for unequal sampling times in this analysis; thus, the value of roadsides to adult monarchs needs further study. Furthermore, a fruitful avenue for further research will be to document the current value of roadsides with respect to adult monarch food sources and the potential for increasing this value.
Monarch egg and larval density had a complex relationship with milkweed density. We analyzed egg and larval density in two ways: monarchs per plant and monarch per m2. The number of monarchs per plant was not affected by plant density. However, monarchs per m2 had an asymptotic or parabolic relationship with milkweed density that suggests an upper threshold or decline in egg and larval density when milkweed density was high (~0.6 milkweeds/m2 (Fig. 4), although this relationship is dependent on other features of the habitat). Thus, the addition of milkweed to roadsides (and potentially other habitat) appears to have an upper limit, beyond which more milkweed does not lead to more monarchs per unit area. There are several possible explanations for this, including potential density dependent effects on immature survival, and female preference for laying eggs over a wider area rather than in a single high-density milkweed patch. For management, this could mean that low density, high frequency milkweed patches are more beneficial than high density, low frequency patches. It should be noted that these patterns may not apply to milkweed densities beyond those used to generate our model, and that more complicated non-linear relationships may underlie these patterns.
We suggest optimism for the conservation potential of roadsides, especially when other habitat is scarce and wildlife-friendly roadside management practices are in place. Conservation efforts on roadside habitat require special consideration, because of the proximity to dangers from the road and surrounding landscape. Pesticide contamination from nearby agricultural fields (Krischik et al. 2007; Hopwood et al. 2015; Mogren and Lundgren 2016; Xu et al. 2016), mortality due to traffic (Mckenna et al. 2001; Ries et al. 2001), effects of runoff from road salt application (Snell-Rood et al. 2014), heavy metal build-up from brake pads (Lagerwerff and Specht 1970; Jaradat and Momani 1999), and vegetation management practices have all been examined as threats to pollinators in roadside habitats (Ries et al. 2001; Hopwood et al. 2015). Because we visited each site only once, we cannot estimate monarch survival, and thus cannot provide insights into the magnitude of these risks. Future studies that include survival data could offer a better picture of how certain roadside types affect monarch production, particularly in terms of risks specific to adjacent land types such as pesticide drift or excessive mowing. Nonetheless, our results demonstrate that roadsides can support both milkweed and immature monarchs while also attracting breeding monarchs when milkweeds are present. Even if productivity is lower in roadsides than other types of sites, the overall contribution is large now and has even more potential with wise management of this high-acreage habitat resource.
References
Akaike H (2011) Akaike’s information criterion. International Encyclopedia of Statistical Science. Springer, Berlin, pp 25–26
Bates D, Maechler M, Bolker B, Walker S (2015a) lme4: Linear mixed-effects models using Eigen and S4. In: R Packag. version 1
Bates D, Maechler M, Bolker B, Walker S (2015b) lme4: Linear mixed-effects models using Eigen and S4. In: R Packag. version 1. http://cran.r-project.org/web/packages/lme4/index.html. Accessed 19 Jun 2015
Baum KA, Mueller EK (2015) Grassland and roadside management practices affect milkweed abundance and opportunities for monarch recruitment. In: Oberhauser KS, Nail KR, Altizer SM (eds) Monarchs in a changing world: biology and conservation of an iconic insect. Cornell University Press, Ithica, pp 197–201
Brower LP, Taylor OR, Williams EH et al (2012) Decline of monarch butterflies overwintering in Mexico: is the migratory phenomenon at risk? Insect Conserv Divers 5:95–100. doi:10.1111/j.1752-4598.2011.00142.x
Fischer SJ, Williams EH, Brower LP, Palmiotto PA (2015) Enhancing monarch butterfly reproduction by mowing fields of common milkweed. Am Midl Nat 173:229–240. doi:10.1674/amid-173-02-229-240.1
Flockhart DTT, Pichancourt JB, Norris DR, Martin TG (2015) Unravelling the annual cycle in a migratory animal: breeding-season habitat loss drives population declines of monarch butterflies. J Anim Ecol 84:155–165. doi:10.1111/1365-2656.12253
Forman RTT, Sperling D, Bissonette JA et al (2003) Road ecology: science and solutions. Rev Lit Arts Am 481
Google Maps (2016) Google Maps. https://www.google.com/maps.
Hartzler RG (2010) Reduction in common milkweed (Asclepias syriaca) occurrence in Iowa cropland from 1999 to 2009. Crop Prot 29:1542–1544. doi:10.1016/j.cropro.2010.07.018
Hartzler RG, Buhler DD (2000) Occurrence of common milkweed (Asclepias syriaca) in cropland and adjacent areas. Crop Prot 19:363–366. doi:10.1016/S0261-2194(00)00024-7
Hopwood J, Black SH, Lee-MÓ“der E et al (2015) Literature review: pollinator habitat enhancement and best management practices in highway rights-of-way 68
Jaradat QM, Momani KA (1999) Contamination of roadside soil, plants, and air with heavy metals in Jordan, a comparative study. Turkish J Chem 23:209–220
Krischik VA, Landmark AL, Heimpel GE (2007) Soil-applied imidacloprid is translocated to nectar and kills nectar-feeding Anagyrus pseudococci (Girault) (Hymenoptera: Encyrtidae). Environ Entomol 36:1238–1245. doi:10.1603/0046-225X(2007)36
Lagerwerff JV, Specht AW (1970) Contamination of roadside soil and vegetation with cadmium, nickel, lead, and zinc. Environ Sci Technol 4:583–586. doi:10.1021/es60042a001
Malcolm SB, Cockrell BJ, Brower LP (1993) Spring recolonization of eastern North America by the monarch butterfly: successive brood or single sweep migration? In: Malcolm SB, Zalucki MP (eds) Biology and conservation of the monarch butterfly. Natural History Museam of Los Angeles County, Los Angeles, pp 253–268
Mckenna DD, Mckenna KM, Malcom SB (2001) Mortality of lepidoptera along roadways in Central Illinois. J Lepid Soc 55:63–68
Mevi-Schutz J, Erhardt A (2005) Amino acids in nectar enhance butterfly fecundity: a long-awaited link. Am Nat 165:411–419. doi:10.1086/429150
Mogren CL, Lundgren JG (2016) Neonicotinoid-contaminated pollinator strips adjacent to cropland reduce honey bee nutritional status. Nat Publ Gr. doi:10.1038/srep29608
Monarch Larva Monitoring Project (2016) Homepage. http://www.mlmp.org/. Accessed 19 Jan 2016
Mueller EK, Baum KA (2014) Monarch—parasite interactions in managed and roadside prairies. J Insect Conserv 18:847–853. doi:10.1007/s10841-014-9691-z
Oberhauser KS, Prysby MD (2008) Citizen science: creating a research army for conservation. Am Entomol 54:103–105.
Oberhauser KS, Prysby MD, Mattila HR et al (2001) Temporal and spatial overlap between monarch larvae and corn pollen. Proc Natl Acad Sci USA 98:11913–11918. doi:10.1073/pnas.211234298
Pleasants J (2016) Milkweed restoration in the Midwest for monarch butterfly recovery: estimates of milkweeds lost, milkweeds remaining and milkweeds that must be added to increase the monarch population. Insect Conserv Divers. doi:10.1111/icad.12198
Pleasants JM, Oberhauser KS (2013) Milkweed loss in agricultural fields because of herbicide use: effect on the monarch butterfly population. Insect Conserv Divers 6:135–144. doi:10.1111/j.1752-4598.2012.00196.x
Prysby MD, Oberhauser KS (2004) Temporal and geographic variation in Monarch densities: citizen scientists document monarch population patterns. In: The monarch butterfly: biology and conservation. Cornell University Press, Ithaca, pp 9–20
R Core Team (2014) R foundation for statistical computing, Vienna, Austria. http://www.r-project.org/. Accessed 16 Feb 2016
Ries L, Debinski DM, Wieland ML (2001) Conservation value of roadside prairie restoration to butterfly communities\rValor de la Conservación de la Restauración de Praderas Ruderales para Comunidades de Mariposas. Conserv Biol 15:401–411. doi:10.1046/j.1523-1739.2001.015002401.x
Saunders SP, Ries L, Oberhauser KS, Zipkin EF (2016) Evaluating confidence in climate-based predictions of population change in a migratory species. Global Ecol Biogeogr 25:1000–1012. doi:10.1111/geb.12461
Semmens BX, Semmens DJ, Thogmartin WE et al (2016) Quasi-extinction risk and population targets for the Eastern, migratory population of monarch butterflies (Danaus plexippus). Nat Sci Rep 6:1–7. doi:10.1038/srep23265
Snell-Rood EC, Espeset A et al (2014) Anthropogenic changes in sodium affect neural and muscle development in butterflies. Proc Natl Acad Sci USA 111:10221–10226. doi:10.1073/pnas.1323607111
Stenoien C, Nail KR, Oberhauser KS (2015) Habitat productivity and temporal patterns of monarch butterfly egg densities in the Eastern United States. Ann Entomol Soc Am 108:670–679. doi:10.1093/aesa/sav054
Stenoien C, Nail KR, Zalucki JM et al (2016) Monarchs in decline: a collateral landscape level effect of modern agriculture. Insect Sci. doi:10.1111/1744-7917.12404
USDA (2016) No Title. http://plants.usda.gov/core/profile?symbol=ASSU3.
Vidal O, Rendón-salinas E (2014) Dynamics and trends of overwintering colonies of the monarch butterfly in Mexico. Biol Conserv 180:165–175. doi:10.1016/j.biocon.2014.09.041
Wassenaar LI, Hobson KA (1998) Natal origins of migratory monarch butterflies at wintering colonies in Mexico: New isotopic evidence. Proc Natl Acad Sci USA 95:15436–15439. doi:10.1073/pnas.95.26.15436
Wickham H (2009) ggplot2: elegant graphics for data analysis. Springer, New York
World Wildlife Fund-Mexico (2016) Aumenta la superficie ocupada por la mariposa monarca en los santuarios mexicanos. http://www.wwf.org.mx/?262370/Aumenta-superficie-ocupada-por-mariposa-monarca-en-santuarios-mexicanos. Accessed 20 Aug 2016
Xu T, Dyer DG, Mcconnell LL et al (2016) Clothianidin in agricultural soils and uptake into corn pollen and canola nectar after multiyear seed treatment applications. Environ Toxicol Chem 35:311–321. doi:10.1002/etc.3281
Zalucki MP, Kitching RL (1982) Temporal and spatial variation of mortality in field population of Danaus plexipus L. and D. chrysippus L. larvae (Lepidoptera: Nymphalidae). Oecologia 53:201–207
Zalucki MP, Lammers JH (2010) Dispersal and egg shortfall in monarch butterflies: what happens when the matrix is cleaned up?. Ecol Entomol 35:84–91. doi:10.1111/j.1365-2311.2009.01160.x
Zalucki MP, Parry HR, Zalucki JM (2015) Movement and egg laying in monarchs: to move or not to move, that is the equation. Austral Ecol. doi:10.1111/aec.12285
Zipkin EF, Ries L, Reeves R et al (2012) Tracking climate impacts on the migratory monarch butterfly. Glob Chang Biol 18:3039–3049. doi:10.1111/J.1365-2486.2012.02751.X
Acknowledgements
We thank Wayne Thogmartin and the United States Geological Survey for providing GIS support and selecting sampling points to initiate our study. A special recognition goes to University of Minnesota Monarch Lab members Dane Elmquist, Cora Lund Preston, and Joe Miller for their support and company in the field. We thank all of the MLMP volunteers as well as coordinators from the U of M Monarch Lab, who go beyond their normal duties to ensure the continuance of this project. We are grateful for staff from Monarch Joint Venture and Monarch Net for providing valuable expertise and resources. Funding was provided by Environmental Stewards in conjunction with the USGS and AmeriCorps to KK and a National Science Foundation Graduate Research Fellowship (00039202) to CS.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kasten, K., Stenoien, C., Caldwell, W. et al. Can roadside habitat lead monarchs on a route to recovery?. J Insect Conserv 20, 1047–1057 (2016). https://doi.org/10.1007/s10841-016-9938-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-016-9938-y