
Abstract
Agricultural intensification reduces the biodiversity of European farmlands. Hay meadows represent an important farmland habitat, traditionally used to produce hay. With decreased demand for hay, the continuation of hay harvest is supported by Agri-environmental schemes across European Union. Modern hay harvest techniques differ from traditional manual harvest by removing the grass instantaneously over large land areas. To minimize adverse effects on meadow invertebrates, diversifying harvest operations is time and space is often recommended, but effects of such diversification are little studied. We compared the impact of uniform hay harvests with harvests executed in patchy manners, using four arthropod groups (butterflies, ground beetles, orthopterans and spiders) at productive, species-poor meadows in the Czech Republic. Butterflies, observed along transects, avoided uniformly cut units, preferring those cut as strips or blocks. In the three remaining groups, recorded using pitfall traps, a majority of species prevailed in traps located in uncut conditions. Synchronous mowing of large areas suppresses population sizes and diminishes the diversity of common arthropods. Besides of direct mortality and depletion of such resources as nectar or shelter, it synchronises sward regrowth, threatening also species requiring short-sward patches. Uniformly executed mowing contradicts the biodiversity conservation goal of Agri-environmental schemes. Diversifying the mowing operations via temporary fallows, or sequential mowing of land units, will improve the situation for common cultural meadows.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Agricultural intensification and abandonment of less productive lands has critically impoverished the biodiversity of temperate farmlands, particularly in Europe (e.g., Stoate et al. 2009; Ekroos et al. 2010). Both the EEC/EU Common Agricultural Policy and the former eastern block farmland collectivisation brought about increased use of fertilisers, pesticides, and modern machinery, as well as land consolidation, irrigation and amelioration schemes, all resulting in increased production, but causing alarming declines of species from many groups of organisms (Donald et al. 2001; Kleijn et al. 2009), including insects (Conrad et al. 2006; Fitzpatrick et al. 2007; Van Dyck et al. 2009). These losses may eventually lead to erosion of crucial ecological services, such as pollination, decomposition, and soil fertility (e.g., Ghazoul 2005; Potts et al. 2009).
Seminatural grasslands, traditionally used for grazing and hay making, suffered at a particularly alarming rate. The magnitude of the losses is difficult to estimate, because statistics in many countries do not distinguish seminatural grasslands from grasslands in various stages of agricultural improvement. The extent of the losses is well known, e.g., for Britain, where about 97% of seminatural grasslands were lost following World War II. (van Dijk 1991). For the Czech Republic, where this study was carried out, relevant statistics do not exist, but local and regional comparisons (e.g., Skaloš 2006) point to losses over 50%, the rest being merged into land units that are, in average, 5–10 times larger than 50 years ago due to loss of hedgerows and field embankments. Invertebrates associated with seminatural grasslands are among the most threatened invertebrates across Europe (e.g., Thomas et al. 1994; Duelli and Obrist 2003). In addition to sensitive specialists, common generalists are declining as well (e.g., Conrad et al. 2006; Gaston and Fuller 2007; Van Dyck et al. 2009).
In Europe, efforts to halt the grassland biodiversity losses rely on two complementary approaches. The first is establishing reserves managed to mimic traditional use, with specific objection to promote grassland biota (Morris 2000). The second, administered as EU agri-environmental schemes [AES] (Pellet 2009), motivates farmers for environmentally benign practices. AES represent the largest monetary transfer to the European biodiversity preservation, many authors view them as a great hope for European flora and fauna (e.g., Kleijn et al. 2001; Marshall et al. 2006) and it is believed that they will eventually represent the main conservation tool for non-reserve lands (Fox et al. 2006). Although schemes for arable land exist, pastures or hay meadows cover by far the largest proportions of thus subsidised lands across Europe (Kleijn and Sutherland 2003; Stoate et al. 2009), including the Czech Republic.
Available studies of AES conservation efficiency, despite some encouraging reports (e.g., Aviron et al. 2009; Merckx et al. 2009), admit that the biodiversity effects are “mixed” at best (Kleijn et al. 2006; Roth et al. 2008). Whereas schemes targeted for specific habitat type or location tend to be successful (Brereton et al. 2008; Wrbka et al. 2008; Merckx et al. 2009), generic prescriptions applied synchronously across large regions often fail to produce diversity benefits (e.g., Kleijn et al. 2001; Blomqvist et al. 2009), and may deplete populations of endangered species (Konvicka et al. 2008). Such schemes do not promote land use heterogeneity, which is crucial for insects (Hendrickx et al. 2007), because heterogeneous environments provide multiple vital resources (e.g., Dennis et al. 2003; Pywell et al. 2005; Geiger et al. 2009).
Mowing for hay production is an important management practice in areas where winter grazing is impossible. Despite its wide use in continental Europe, a majority of studies of mowing impacts on insect diversity were carried out in England (e.g., Southwood and van Emden 1967; Morris 1979; Purvis and Curry 1978; Morris and Lakhani 1979; Morris and Plant 1983; Morris and Rispin 1988). This is somehow surprising, both if compared with much wider geographic scope of grazing effects studies (e.g., Purvis and Curry 1981; Poyry et al. 2006; Sjodin et al. 2008; Dover et al. 2011a), and if considered that mowing has historically represented a major use of more productive grasslands. In contrast to grazing, mowing exports plant biomass more rapidly, temporarily depleting resources such as herbivore food, and causing direct animal mortality (Morris 2000; Humbert et al. 2009, 2010; Dover et al. 2010). Many species find their optima under less intensive management, or under different time harvest time, than is optimal for farmers (e.g., Morris 1981a, b; Ockinger et al. 2006; Poyry et al. 2006), and such species likely persisted via tracking of temporarily existing unmown patches on traditionally mowed lands (Konvicka et al. 2008; Baguette et al. 2011). All these concerns are rising in importance, because mowing is currently practised even in some formerly grazed reserves as alternative management (Balmer and Erhardt 2000; Morris 2000).
Based on the impacts of mowing, it is often recommended to proceed in a mosaic-like manners, creating checkerboards or strips of cut and temporarily uncut sward (Humbert et al. 2009; Schmidt et al. 2008). These prescriptions are being increasingly implemented at localities of endangered species (e.g., Johst et al. 2006; Schmidt et al. 2008). There remains, however, only weak empirical evidence advocating such practices. On the other hand, the calls for mowing diversification generate resistance from the AES funding agencies, who argue that mosaic approaches are more costly than simple edge-to-edge cuts repeated several times a year, and also impair landscape aesthetics due to existence of “weedy” patches within meadows.
Here, we assess the impacts of diversification of mowing regimes on four arthropod groups with diverse life strategies (butterflies, ground beetles, orthopterans and spiders), inhabiting alluvial hay meadows within an intensively farmed Czech Republic landscape. The study was launched during preparation of a management plan for a reserve which, contrary to the most reserves in the region, was established to protect a historical scenic landscape. As a result of intensive meadow management in the past and isolation from other natural habitats, the studied meadows are species-poor, thus representing common productive grasslands of Central Europe.
Materials and methods
Study area
Babiččino údolí National Nature Reserve (50°24′52″N, 16°3′16″E, alt. 277–350 m) encompasses a 3.5 km long section of the Úpa River valley, situated amidst the intensively-farmed landscape of Eastern Bohemia, Czech Republic. The climate is moderately warm and slightly humid. The reserve is the historical heritage site of the Ratibořice mansion, famous from the 19th century patriotic novel ‘Babička’ (The Grandmother). The valley slopes are covered by deciduous forests, whereas the bottom, with average width 250 m, contains 68 ha of improved alluvial meadows. Being situated on poor acidic soils, they were mainly grazed until the 1840s, when drainage ditches were excavated, allowing using the meadows for hay making. This system persisted until the intensification period in 1970s, when use of fertilisers increased and the meadows were improved by reseeding. In the late 1980s, in order to restore a more natural grasslands character, fertilising was diminished and the meadows were sown with local mixtures.
Since this period until 2004, all the meadows have been managed by machine mowing twice per year. Because demand for hay decreased over this period, the primary reason for mowing of the reserve is to preserve its scenic character. Despite this, the mowing is commissioned to local farming cooperative, which is using the more efficient and destructive rotary mowers, in some years all meadows were even mulched. The cutting method thus represents the highly efficient technique widely across Central Europe.
Mowing diversification experiment
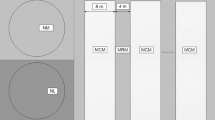
Hedges, ditches, roads and the Úpa River itself divide the Babiččino údolí meadows into distinct units (Fig. 1). In 2005 and 2006, three distinct mowing regimes were experimentally applied: Complete units were mown uniformly edge-to-edge, in June and again in August; this regime is applied to a vast majority of AES meadows the Czech Republic. Strip units were mown so that during the June cut, 5–10 m wide unmown strips alternated with mown strips of a similar width, and the mown and unmown sections were switched in August. Finally, Block units were also cut only partly, but the width of unmown sections was ca 50 m.

Map of the Babiččino údolí reserve, showing the positions of differently managed hay meadow units, as applied during the study, and the positions of the butterfly transects and the pitfall-trapping sites
Studied groups
We targeted four arthropod groups with diverse life histories, all often used as models in conservation studies. Butterflies are phytophagous as larvae but depend on nectar as adults, and encompass both highly mobile and sedentary species. Ground beetles are mostly ground-dwelling predators or omnivores, little dependent on the species composition of vegetation, and relatively immobile. Orthopterans are generalist phytophages or omnivores representing an important constituent of grassland insect biomass. Spiders are predators, hunting both at the ground and in vegetation, more mobile than ground beetles.
Species names and life history information follow Benes et al. (2002) for butterflies, Kočárek et al. (2005) for orthopterans, Hůrka (1996) for ground beetles, and Buchar and Růžička (2002) for spiders.
Data collecting
Butterfly transects
We established 11 fixed linear transects (mean length 218, 21 m SE), crossing centres of 11 units (Fig. 1) but terminating 30 m from unit edges to minimise species leakage from neighbouring habitats. We walked the transects in 2005 and 2006, approximately fortnightly with 11 walks each year (from May, with the start of the first mowing, to September), always at a slow pace (ca 2 km per h) between 10.00 and 16.00 CEST and in suitable weather (>15°C, none to mild wind).
Butterflies were counted during slow walks, in a 5 m radius in front of the recorder. Transect Length was treated as covariable in all models. For each walk, we recorded the following Weather factors: ambient temperature (Temp), wind speed (Wind) and cloudiness (Cloud), the latter two on 1–5 ordinal scales. Covariables describing sward conditions were Height (averaged from 5 measurements along each transect and walk), nectar abundance (Nabd: ordinal 1–5 with 1 being the lowest), and nectar diversity (Ndiv: number of actually flowering species along the transect). The dependent variable, coded as three factors, was mowing Regime. Although the regime changed between the two studied years at some units (Fig. 1), the transect positions remained the same.
Pitfall traps
In 2006, we carried out pitfall trapping at seven units (Fig. 1), four managed by total cut and three as blocks or strips (herein partial). The traps (plastic cups: diameter 9 cm, depth 15 cm, containing 5% formaldehyde) formed lines of eight traps each, set in 8 m intervals, always in units’ centres. At partial units, the traps were set to cut and temporally uncut parts proportionally to the representation of cut and uncut sward at the unit. The trapping, carried out from May 8 to September 17, proceeded so that the traps were always exposed for 3 days and then inactivated for 10 days. There were 10 trapping periods in total. Following each period, we sorted and identified all ground beetles, orthopterans and spiders captured.
To describe Weather during these 3-days trapping periods, we obtained hourly weather data from the closest meteorological station (Velichovky, 50°21′20.027″N, 15°50′31.349″E). Sums of these hourly values were used to describe wind intensity (Wind S ), humidity (Hum S ), solar radiation (Rad S ), precipitation (Prec S ) and temperature (Temp S ). Sward variables were Nabd, Ndiv and Height as for butterfly transects, but recorded within 5 m diameter circles around each trap.
Analyses
We used the redundancy analysis (RDA), a constrained linear ordination method, to relate the species composition to meadow management. We analysed separately the transect recorded butterflies and the trapped ground beetles, orthopterans and spiders, in CANOCO for Windows 4.5 (ter Braak and Smilauer 2002). Statistical significance of the ordinations was assessed via the Monte Carlo tests (999 permutations, full model). Individual transect walks and pitfall traps are herein considered as samples; species scores were divided by standard deviation, and species were centered (Lepš and Šmilauer 2003).
Butterfly transects
The explanatory variable was Regime, coded as a categorical variable and constant for entire unit and year. The permutation tests reflected spatial and temporal distribution of records: the two subsequent seasons were treated as blocks to filter out yearly differences; the 22 repeated walks to each transect formed whole plots, permuted as time series, whereas individual transects were permuted randomly.
Transect Length was treated as a nuisance covariable in all models. We tested for independent effect of Regime, and for its effect after filtering out Weather effects (an adequate weather model was obtained via CANOCO forward selection procedure from all weather covariables). In parallel, we forward-selected and tested the effects of Sward variables, both without and with Weather effects in the model. Finally, we tested if Regime had some residual effect on the model after Sward covariables, and both Weather and Sward covariables, entered the model.
Ground dwelling arthropods
The local situation around each trap and visit (i.e., if located in cut or uncut part of a unit) was explanatory variable. As cut, we considered sward <20 cm, which corresponds to ca. 2 weeks after mowing, and the state of individual traps changed among visits. For permutation design, entire units formed blocks, filtering out the effects of different locations on trap samples. The ten trapping periods were whole plots permuted as time series, and the lines of eight traps were split plots permuted as line transects.
As in case of butterflies, we first tested for the separate effect of Regime, and the effect of Regime after considering Weather variables. We also defined best-fitting Sward models, again without and with Weather, using CANOCO forward selection. Finally, we entered the forward-selected Sward variables onto the Regime models.
Results
Arthropod assemblage composition
The recorded assemblages were species-poor. Numbers of species/individuals were 32/2,197 for butterflies, 34/4,220 for ground beetles, 6/116 for orthopterans, and 30/8,318 for spiders. Rank-abundance plots (Fig. 2) reveal that all four assemblages were dominated by a few extremely abundant species. Over 50% of all records were represented by three butterflies (Pieris rapae, Pieris napi, Maniola jurtina), two ground beetles (Poecilus cupreus, P. versicolor), two orthopterans (Chorthippus paralellus, Tetrix subulata) and just a single spider (Pardosa palustris).

The butterfly assemblage consisted of common grasslands generalists (Maniola jurtina, Coenonympha pamphilus, Thymelicus lineola) and ubiquitous highly mobile species (Pieris napi, P. rapae, P. brassicae, Aglais urticae), supplemented by a high abundance of the grassland specialist Aricia agestis. Only two species, Phengaris nausithous and P. teleius, are endangered in the Czech Republic, but these two species were recorded in very low abundance. In the case of ground beetles, most of recorded species were widespread generalists of open habitats (Carabus violaceus, Poecilus cupreus, Amara aenea), supplemented by common species of more xeric (Amara equestris) and wet (Agonum sexpunctatum, Clivina collaris) grasslands, and a few individuals of woodland species (Carabus coriaceus, Cychrus caraboides). The most common orthopterans were two species of herbivorous Chorthippus grasshoppers (C. parallelus, C. biguttulus), and detritovorous Tetrix subulata. All spiders were also widely distributed species of open habitats (Xysticus kochi, Pardosa palustris, Pachygnatha degeeri), some of them displaying an affinity to wet conditions (Pardosa amentata, Antistea elegans, Pirata latitans).
Mowing and butterfly activity
Mowing regime had a significant effect on butterflies (Table 1). Temperature was the only significant Weather covariable. Filtering out the temperature effect produced an ordination, in which a majority of species avoided Complete cut (Fig. 3). The three species with some affinity towards Complete (Araschnia levana, Argynnis paphia, Celastrina argiolus) were woodland butterflies, probably not developing at the meadows and visiting them only temporarily. No straightforward differences in life history traits between species with affinity towards Blocs and Strips were apparent, as both regimes attracted both widespread and mobile generalists (blocks: Aglais urticae, Issoria lathonia, Pieris rapae; strips: Colias hyale, Inachis io, Pieris napis), and relatively sensitive specialists (blocks: Aricia agestis, Phengaris nausithous; strips: Boloria dia, Phengaris teleius).

Ordination diagram (RDA) showing association of butterflies with the three hay mowing regimes. 1, Aglais urticae; 2, Aphanthopus hyperanthus; 3, Araschnia levana; 4, Argynnis paphia; 5, Aricia agestis; 6, Boloria dia; 7, Celastrina argiolus; 8, Coenonymha pamphilus; 9, Colias hyale; 10, Erynnis tages; 11, Gonepteryx rhamni; 12, Inachis io; 13, Issoria lathonia; 14, Lasiommata megera; 15, Leptidea reali; 16, Lycaena phlaeas; 17, Lycaena tityrus; 18, Phengaris nausithous; 19, Phengaris teleius; 20, Maniola jurtina; 21, Melanargia galathea; 22, Ochlodes sylvanus; 23, Papilio machaon; 24, Pieris brassicae; 25, Pieris napi; 26, Pieris rapae; 27, Polygonia c-album; 28, Polyomatus icarus; 29, Pontia daplidice; 30, Thymelicus lineola; 31, Vanessa atalanta; 32, Vanessa cardui
Nectar supply, expressed as either Nabd or Ndiv, represented the only sward factors structuring the butterfly records (Table 1). All butterflies inclined towards high nectar supply (figure not shown). Regime model with Nabd as a covariate remained significant, however, suggesting some influence of other factors than nectar. A likely candidate was Height, because model with both Height and Nabd consumed the entire explicable variation.
Mowing and ground dwelling arthropods
For all three groups, Regime affected the composition of pitfall trap samples, even after considering Weather effects (Table 2). The majority of ground beetles (Fig. 4) inclined towards Uncut conditions. The few species inclining towards Cut conditions were mostly species tolerating frequently disturbed, even ploughed, environments (e.g., Amara aenea, Calathus fuscipes, Poecilus cupreus), whereas more specialised but still common species (such as xerophilous Amara equestris, hygrophilous Epaphius secalis, or edge specialist Carabus coriaceus) inclined towards Cut conditions. In orthopterans (Fig. 5), the polarity between Cut and Uncut conditions reflected a gradient of humidity requirements in the three most abundant species: the relatively hygrophilous Tetrix subulata inclined towards unmown section, generalist Chorthippus parallelus was intermediate, and C. biguttulus, a species preferring drier grasslands, inclined towards mown ones. Finally, spiders (Fig. 6) with affinity to Uncut conditions were mostly widespread generalists (e.g., Erigone dentipalpis, Pardosa lugubris, Pardosa palustris), whereas hygrophilous species (e.g., Antistea elegans, Centromerus sylvaticus, Pardosa amentata) inclined towards Cut conditions.

Ordination diagrams (RDA) showing affinities of ground beetles to cut and uncut parts of the studied units. 1, Agonum sexpunctatum; 2, Amara aenea; 3, Amara aulica; 4, Amara equestris; 5, Amara montivaga; 6, Anchomenus dorsalis; 7, Anisodactylus signatus; 8, Badister sodalis; 9, Bembidion properans; 10, Bembidion quadrimaculatum; 11, Calathus fuscipes; 12, Calathus melanocephalus; 13, Carabus coriaceus; 14, Carabus granulatus; 15, Carabus scheidleri; 16, Carabus ulrichi; 17, Carabus violaceus; 18, Cicindela campestris; 19, Clivina collaris; 20, Cychrus scarabeoides; 21, Epaphius secalis; 22, Harpalus luteicornis; 23, Loricera pilicornis; 24, Ophonus nitidulus; 25, Ophonus schaubergerianus; 26, Panagaeus cruxmajor; 27, Platynus assimilis; 28, Poecilus cupreus; 29, Poecilus versicolor; 30, Pseudoophonus rufipes; 31, Pterostichus nigrita; 32, Pterostichus ovoideus; 33, Pterostichus vernalis; 34, Pterostichus vulgaris

Ordination diagrams (RDA) showing affinities of orthopterans to cut and uncut parts of the studied units. 1, Chorthippus biguttulus; 2, Chorthippus dorsatus; 3, Chorthippus paralleulus; 4, Tetrix subulata; 5, Tetrix tenuicornis; 6, Tetrix undulata

Ordination diagrams (RDA) showing affinities of spiders to cut and uncut parts of the studied units. 1, Alopecosa cuneata; 2, Alopecosa pulverulenta; 3, Antistea elegans; 4, Bathyphantes gracilis; 5, Centromerus sylvaticus; 6, Diplostyla concolor; 7, Drassyllus lutetianus; 8, Drassyllus praeficus; 9, Drassyllus pusillus; 10, Erigone atra; 11, Erigone dentipalpis; 12, Hahnia nava; 13, Hahnia pusilla; 14, Micaria pulicaria; 15, Micrargus subaequalis; 16, Ozyptila trux; 17, Pachygnatha degeeri; 18, Pardosa amentata; 19, Pardosa lugubris; 20, Pardosa palustris; 21, Pardosa prativaga; 22, Pardosa pullata; 23, Pelecopsis radicicola; 24, Pirata latitans; 25, Pisaura mirabilis; 26, Trochosa ruricola; 27, Xysticus audax; 28, Xysticus cristatus; 29, Xysticus kochi; 30, Zora spinimana
Testing for Sward conditions revealed significant effects on ground beetles and spiders. Ground beetles differed from spiders, however, so that after inclusion of Sward variables, Regime retained some significant effect on the former but not on the latter. The effects of mowing on spiders thus can be attributed solely to vegetation height and diversity, which is not the case for ground beetles. This analysis was not carried out for orthopterans, because the low total number of species did not leave any unexplained variation (Table 2).
Discussion
Hay harvest restructures arthropod assemblages in intensively managed alluvial meadows, and diversifying of mowing operations promotes biodiversity at these species-poor habitats. Transect recording of butterflies revealed that the overwhelming majority of species preferred units mown heterogeneously, with parts left temporarily unmown. In ground beetles, orthopterans and spiders, pitfall traps surrounded by recently cut sward differed in species composition from those surrounded by uncut sward. The effects of mowing methods were visible on a scale of hectares for butterflies, and on a scale of metres for ground beetles, orthopterans and spiders.
At the small scale of the study relative to the mobility of the invertebrates, our approach tracked invertebrate activity, rather than habitat preferences. We do not view it as a major problem. Animal activity reflects the location and distribution of their resources. From this viewpoint, mowing has two likely effects. It directly kills some individuals, and depletes resources for others, forcing them to disperse. These effects both change the composition of local assemblages (directly assessed by our sampling methods) and influence individual survival and population persistence (a main focus of conservation management).
Comparison among taxa
An overwhelming majority of butterflies inclined towards heterogeneously managed units. The impact of mowing on butterflies is easily understood, but surprisingly little appreciated. It causes direct mortality, because mowing is normally performed in early mornings, when the butterflies are still inactive (Dover et al. 2010). It also depletes nectar, forcing surviving individuals to leave or to face starvation, and destroys shelters, exposing them to adverse weather or predators. If performed over large land units, or, due to synchronisation of operations, over entire landscapes (Konvicka et al. 2008), dispersing individuals may perish before they locate new resources (cf. Dover and Settele 2009).
The analyses for butterflies reflected whole-season management, not momentary sward conditions. The results allow concluding that butterflies avoided units cut twice per year, despite the periods between cuts being sufficiently long for plants to sprout new flowers. It is presumable that besides of nectar depletion, too frequent mowing affects the suitability of grasslands for butterflies in more subtle ways, such as by enhancing larval mortality. Clearly, less intensive and/or more diversified mowing will support more butterflies.
In the remaining three taxa, we worked on too small a scale to discuss landscape-level effects. Part of the patterns detected could be attributed to direct mortality caused by mechanised mowing, as in butterflies (e.g., Gardiner and Hill 2006). Still, the response variable reflected momentary sward height, and it is more legitimate to interpret the results in terms of species’ behaviour.
With activity largely restricted to soil surfaces, ground beetles unlikely suffer too much mortality from mowing. Instead, they require various sheltering structures, often located in plant litter (Batary et al. 2007; Kagawa and Maeto 2009), which is more abundant in less frequently mown sward. Consequently, more species, including the most abundant ones, headed towards traps located in uncut sward. In contrast, the few species preferring drier conditions (e.g., Amara equestris) inclined towards shortly cut patches.
Unlike ground beetles and similarly to butterflies, orthopterans utilise grassland environments in a three-dimensional manner. Short homogeneous sward may be unsuitable due to the lack of tussocks that provide shelter from weather (Gardiner and Hill 2004; Gardiner and Hassall 2009) and predation.
The background of spiders’ responses is likely intermediate between butterflies and ground beetles. As predators, spiders do not use nectar, but unlike ground beetles, they utilise entire three-dimensional structure of grassland environments for prey hunting, sheltering, and other activities. Indirect effects are also possible, as spiders follow the abundance of their insect prey. It was repeatedly shown that highly structured vegetation increases the diversity of spiders, the type of mowing machine plays also an important role (e.g., Bell et al. 2001; Schmidt and Tscharntke 2005).
Homogeneous versus heterogeneous mowing
Although mowing always increases the mortality and depletes resources for grassland invertebrates, the very existence of the hay meadow habitat depends on it. As illustrated by our results, mowing mode and intensity crucially affect the composition of local invertebrate assemblages. In the past, with less efficient mechanisation and more scattered land holding, mowing proceeded patchily, reflecting the needs of individual farmers. The slow pace of manual mowing allowed the invertebrates to escape, and as it took days and weeks to mow larger units, the temporarily depleted resources had enough time to recover (Morris 2000; Humbert et al. 2009). With rapid and synchronous machinery mowing, the associated mortality likely exceeds the replacement ability of local invertebrate populations (Gardiner and Hill 2006; Schmidt et al. 2008; Dover et al. 2010). The mowing technique used also plays also a role. Although both are destructive to invertebrates, bar mowers are safer than more widely used rotary mowers (Humbert et al. 2010). Totally devastating must be mulching, which shreds biomass to small pieces.
Traditional heterogeneous mowing had yet another, rarely appreciated effect. The preference of some ground beetles, spiders and orthopterans for freshly cut surfaces indicates that short-sward patches also constitute a resource for some animal activities (e.g., butterflies bask on cut patches in cool weather: Ouin et al. 2004). Frequently mown patches certainly existed in traditional landscapes, owing to such farmers’ needs as supply of green fodder during summer, and species requiring such patches likely tracked them in a similar manner as species requiring taller vegetation tracked temporarily unmown swards. Because synchronous mowing synchronises the subsequent regrowth, species requiring short sward may be impaired as seriously, as the rapid removal of biomass by modern machinery impairs tall-sward species (Berga and Gustafsonb 2007; Šálek and Schröpfer 2008).
We documented that the diversification of mowing diversifies invertebrate assemblages on very small scales (among meadows units for butterflies, and within the units for ground beetles, spiders and orthopterans), on meadows that were initially species poor. Therefore, any diversification of mowing operations will contribute to the diversity of animal resources, and hence species diversity. The finding that even common species inhabiting biologically poor grasslands responded to management diversification is important for managing grasslands with no special biological richness. Earlier studies documented that more diversified management is indispensable for sensitive grassland specialists (Johst et al. 2006; Spitzer et al. 2009). We now showed that it would also benefit common and widespread species.
Conclusion
Traditional agricultural landscapes were much more diverse, in terms of both land holding and land use, than modern intensified farmland. Everything else being equal, decreasing size of management units, increasing temporal asynchronicity of operations, or use of more diverse technologies, all benefit farmland biodiversity (e.g., Rundlof et al. 2008; Butler et al. 2009). Sadly, the homogenisation trends typical for productive farming continue even on lands subsidised for environmental goods, because funding agencies favour simple prescriptions enforceable over entire regions (Beckmann et al. 2009). This is no longer tolerable, if the massive monetary transfers of AES are to meet their goals (Dover et al. 2011b).
Even with modern technology, hay harvest can be easily diversified either in space (temporarily uncut strips or blocks) or time (sequential mowing within an area). Despite being increasingly used in grassland reserves (e.g., Morris 2000; WallisDeVries et al. 2002), these practices do not even bear an established name and are variously labelled as “rotational fallows” (Schmidt et al. 2008) or “mosaic mowing” (Konvicka et al. 2008). It is clear that insular reserves, no matter how well managed, cannot maintain regional species pools indefinitely, due to the effects of stochastic catastrophes on isolated populations (Hanski 1999; Rosenzweig 2003). Management diversification needs to expand beyond reserves, to lands farmed under environmental subsidies. The diversification measures applied in this study will unlikely restore the entire biotic richness of historical grasslands, but will certainly slow down the biodiversity loss.
References
Aviron S, Nitsch H, Jeanneret P, Buholzer S, Luka H, Pfiffner L, Pozzi S, Schupbach B, Walter T, Herzog F (2009) Ecological cross compliance promotes farmland biodiversity in Switzerland. Front Ecol Environ 7:247–252
Baguette M, Clobert J, Schtickzelle N (2011) Metapopulation dynamics of the bog fritillary butterfly: experimental changes in habitat quality induced negative density-dependent dispersal. Ecography 34:170–176
Balmer O, Erhardt A (2000) Consequences of succession on extensively grazed grasslands for central European butterfly communities: rethinking conservation practices. Conserv Biol 14:746–757
Batary P, Baldi A, Szel G, Podlussany A, Rozner I, Erdos S (2007) Responses of grassland specialist and generalist beetles to management and landscape complexity. Divers Distrib 13:196–202
Beckmann V, Eggers J, Mettepenningen E (2009) Deciding how to decide on agri-environmental schemes: the political economy of subsidiarity, decentralisation and participation in the European Union. J Environ Plan Manag 52:689–716
Bell JR, Wheater CP, Cullen WR (2001) The implications of grassland and heathland management for the conservation of spider communities: a review. J Zool 255:377–387
Beneš J, Konvička M, Dvořák J, Fric Z, Havelda Z, Pavlíčko A, Vrabec V, Weidenhoffer Z (2002) Butterflies of the Czech Republic: distribution and conservation, I, II. SOM, Prague
Berga Å, Gustafsonb T (2007) Meadow management and occurrence of corncrake Crex crex. Agric Ecosyst Environ 120:139–144
Blomqvist MM, Tamis WLM, de Snoo GR (2009) No improvement of plant biodiversity in ditch banks after a decade of agri-environment schemes. Basic Appl Ecol 10:368–378
Brereton TM, Warren MS, Roy DB, Stewart K (2008) The changing status of the Chalkhill Blue butterfly Polyommatus coridon in the UK: the impacts of conservation policies and environmental factors. J Insect Conserv 12:629–638
Buchar J, Růžička V (2002) Catalogue of spiders of the Czech Republic. Peres, Prague
Butler SJ, Brooks D, Feber RE, Storkey J, Vickery JA, Norris K (2009) A cross-taxonomic index for quantifying the health of farmland biodiversity. J Appl Ecol 46:1154–1162
Conrad KF, Warren MS, Fox R, Parsons MS, Woiwod IP (2006) Rapid declines of common, widespread British moths provide evidence of an insect biodiversity crisis. Biol Conserv 132:279–291
Dennis RLH, Shreeve TG, van Dyck H (2003) Towards a functional resource-based concept for habitat: a butterfly biology viewpoint. Oikos 102:417–426
Donald PF, Green RE, Heath MF (2001) Agricultural intensification and the collapse of Europe’s farmland bird populations. P Roy Soc Lond B Biol Sci 268:25–29
Dover J, Settele J (2009) The influences of landscape structure on butterfly distribution and movement: a review. J Insect Conserv 13:3–27
Dover JW, Rescia A, Fungariño S, Fairburn J, Carey P, Lunt P, Dennis RLH, Dover CJ (2010) Can hay harvesting detrimentally affect adult butterfly abundance? J Insect Conserv 14:413–418
Dover JW, Rescia A, Fungariño S, Fairburn J, Carey P, Lunt P, Arnot C, Dennis RLH, Dover CJ (2011a) Land-use, environment, and their impact on butterfly populations in a mountainous pastoral landscape: species richness and family-level abundance. J Insect Conserv 15:207–220
Dover JW, Spencer S, Collins S, Hadjigeorgiou I, Rescia A (2011b) Grassland butterflies and low intensity farming in Europe. J Insect Conserv 15:129–137
Duelli P, Obrist MK (2003) Regional biodiversity in an agricultural landscape: the contribution of seminatural habitat islands. Basic Appl Ecol 4:129–138
Ekroos J, Heliola J, Kuussaari M (2010) Homogenization of lepidopteran communities in intensively cultivated agricultural landscapes. J Appl Ecol 4:459–467
Fitzpatrick U, Murray TE, Paxton RJ, Breen J, Cotton D, Santorum V, Brown MJF (2007) Rarity and decline in bumblebees—a test of causes and correlates in the Irish fauna. Biol Conserv 136:185–194
Fox R, Asher J, Brereton T, Roy D, Warren M (2006) The state of butterflies in Britain and Ireland. Nature Bureau, Newbury
Gardiner T, Hassall M (2009) Does microclimate affect grasshopper populations after cutting of hay in improved grassland? J Insect Conserv 13:97–102
Gardiner T, Hill J (2004) Directional dispersal patterns of Chorthippus parallelus (Orthoptera: Acrididae) in patches of grazed pastures. J Orthoptera Res 13:135–141
Gardiner T, Hill J (2006) Mortality of Orthoptera caused by mechanised mowing of grassland. Br J Entomol Nat Hist 19:38–40
Gaston KJ, Fuller RA (2007) Biodiversity and extinction: losing the common and the widespread. Prog Phys Geog 31:213–225
Geiger F, Wackers F, Bianchi FJJA (2009) Hibernation of predatory arthropods in semi-natural habitats. Biocontrol 54:529–535
Ghazoul J (2005) Buzziness as usual? Questioning the global pollination crisis. Trends Ecol Evol 20:367–373
Hanski I (1999) Metapopulation ecology. Oxford University Press, UK
Hendrickx F, Maelfait JP, Van Wingerden W, Schweiger O, Speelmans M, Aviron S, Augenstein I, Billeter R, Bailey D, Bukacek R, Burel F, Diekotter T, Dirksen J, Herzog F, Liira J, Roubalova M, Vandomme V, Bugter R (2007) How landscape structure, land-use intensity and habitat diversity affect components of total arthropod diversity in agricultural landscapes. J Appl Ecol 44:340–351
Humbert JY, Ghazoul J, Walter T (2009) Meadow harvesting techniques and their impacts on field fauna. Agric Ecosyst Environ 130:1–8
Humbert JY, Ghazoul J, Sauter GJ, Walter T (2010) Impact of different meadow mowing techniques on field invertebrates. J Appl Entomol 134:592–599
Hůrka K (1996) Carabidae of the Czech and Slovak Republics. Kabourek, Zlin
Johst K, Drechsler M, Thomas J, Settele J (2006) Influence of mowing on the persistence of two endangered large blue butterfly species. J Appl Ecol 43:333–342
Kagawa Y, Maeto K (2009) Spatial population structure of the predatory ground beetle Carabus yaconicus (Coleoptera: Carabidae) in the mixed farmland-woodland satoyama landscape of Japan. Eur J Entomol 106:385–391
Kleijn D, Sutherland WJ (2003) How effective are European agri-environment schemes in conserving and promoting biodiversity? J Appl Ecol 40:947–969
Kleijn D, Berendse F, Smit R, Gilissen N (2001) Agri-environment schemes do not effectively protect biodiversity in Dutch agricultural landscapes. Nature 413:723–725
Kleijn D, Baquero RA, Clough Y, Diaz M, De Esteban J, Fernandez F, Gabriel D, Herzog F, Holzschuh A, Johl R, Knop E, Kruess A, Marshall EJP, Steffan-Dewenter I, Tscharntke T, Verhulst J, West TM, Yela JL (2006) Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecol Lett 9:243–254
Kleijn D, Kohler F, Baldi A, Batary P, Concepcion ED, Clough Y, Diaz M, Gabriel D, Holzschuh A, Knop E, Kovacs A, Marshall EJP, Tscharntke T, Verhulst J (2009) On the relationship between farmland biodiversity and land-use intensity in Europe. P Roy Soc B Biol Sci 276:903–909
Kočárek P, Holuša J, Vidlička L (2005) Blattaria, Mantodea, Orthoptera and Dermaptera of the Czech and Slovac Republics. Kabourek, Zlin
Konvicka M, Benes J, Cizek O, Kopecek F, Konvicka O, Vitaz L (2008) How too much care kills species: grassland reserves, agri-environmental schemes and extinction of the Colias myrmidone butterfly from its former stronghold. J Insect Conserv 12:519–525
Lepš J, Šmilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Marshall EJP, West TM, Kleijn D (2006) Impacts of an agri-environment field margin prescription on the flora and fauna of arable farmland in different landscapes. Agric Ecosyst Environ 113:1–4
Merckx T, Feber RE, Riordan P, Townsend MC, Bourn NAD, Parsons MS, Macdonald DW (2009) Optimizing the biodiversity gain from agri-environment schemes. Agric Ecosyst Environ 130:177–182
Morris MG (1979) Responses of grassland invertebrates to management by cutting. II. Heteroptera. J Appl Ecol 16:417–432
Morris MG (1981a) Responses of grassland invertebrates to management by cutting. III. Adverse effects on Auchenorhyncha. J Appl Ecol 18:107–123
Morris MG (1981b) Responses of grassland invertebrates to management by cutting. IV. Positive responses of Auchenorhyncha. J Appl Ecol 18:763–771
Morris MG (2000) The effects of structure and its dynamics on the ecology and conservation of arthropods in British grasslands. Biol Conserv 95:129–142
Morris MG, Lakhani KL (1979) Responses of grassland invertebrates to management by cutting. I. Species diversity of Hemiptera. J Appl Ecol 16:77–98
Morris MG, Plant R (1983) Responses of grassland invertebrates to management by cutting. V. Changes following the cessation of management. J Appl Ecol 20:157–177
Morris MG, Rispin WE (1988) A beetle fauna of oolitic limestone grassland, and the response of species to conservation management by different cutting regimes. Biol Conserv 43:87–105
Ockinger E, Eriksson AK, Smith HG (2006) Effects of grassland abandonment, restoration and management on butterflies and vascular plants. Biol Conserv 133:291–300
Ouin A, Aviron S, Dover J, Burel F (2004) Complementation/supplementation of resources for butterflies in agricultural landscapes. Agric Ecosyst Environ 103:473–479
Pellet J (2009) Does delaying the first mowing date increase biodiversity in European farmland meadows? http://www.environmentalevidence.org/Documents/Protocol72.pdf. Accessed 21 Sep 2010
Potts SG, Woodcock BA, Roberts SPM, Tscheulin T, Pilgrim ES, Brown VK, Tallowin JR (2009) Enhancing pollinator biodiversity in intensive grasslands. J Appl Ecol 46:369–379
Poyry J, Luoto M, Paukkunen J, Pykala J, Raatikainen K, Kuussaari M (2006) Different responses of plants and herbivore insects to a gradient of vegetation height: an indicator of the vertebrate grazing intensity and successional age. Oikos 115:401–412
Purvis G, Curry JP (1978) The effects of grazing and silage production systems on the epigeal fauna of grassland. Sci Proc R Dublin Soc A 6:317–325
Purvis G, Curry JP (1981) The influence of sward management on foliage arthropod communities in a ley grassland. J Appl Ecol 18:711–725
Pywell RF, James KL, Herbert I, Meek WR, Carvell C, Bell D, Sparks TH (2005) Determinants of overwintering habitat quality for beetles and spiders on arable farmland. Biol Conserv 123:79–90
Rosenzweig ML (2003) Win-win ecology: how the earth’s species can survive in the midst of human enterprise. Oxford University Press, New York
Roth T, Amrhein V, Peter B, Weber D (2008) A swiss agri-environment scheme effectively enhances species richness for some taxa over time. Agric Ecosyst Environ 125:167–172
Rundlof M, Bengtsson J, Smith HG (2008) Local and landscape effects of organic farming on butterfly species richness and abundance. J Appl Ecol 45:813–820
Šálek M, Schröpfer L (2008) Population decline of the little owl in the Czech Republic. Pol J Ecol 56:527–534
Schmidt MH, Tscharntke T (2005) The role of perennial habitats for Central European farmland spiders. Agric Ecosyst Environ 105:235–242
Schmidt MH, Rocker S, Hanafi J, Gigon A (2008) Rotational fallows as overwintering habitat for grassland arthropods: the case of spiders in fen meadows. Biodivers Conserv 17:3003–3012
Sjodin NE, Bengtsson J, Ekbom B (2008) The influence of grazing intensity and landscape composition on the diversity and abundance of flower-visiting insects. J Appl Ecol 45:763–772
Skaloš J (2006) Patterns and changes of intensively utilised agricultural landscape in the Czech Republic between 1937 and 2002, aerial photography analysis. Ekologia 21:232–248
Southwood TRE, van Emden HF (1967) A comparison of the fauna of cut and uncut grasslands. Z Angew Entomol 60:188–198
Spitzer L, Benes J, Dandova J, Jaskova V, Konvicka M (2009) The large blue butterfly, Phengaris [Phengaris] arion, as a conservation umbrella on a landscape scale: the case of the Czech Carpathians. Ecol Indicators 9:1056–1063
Stoate C, Baldi A, Beja P, Boatman ND, Herzon I, van Doorn A, de Snoo GR, Rakosy L, Ramwell C (2009) Ecological impacts of early 21st century agricultural change in Europe—a review. J Environ Manag 91:22–46
ter Braak CJF, Smilauer P (2002) CANOCO reference manual and CanoDraw for Windows user’s guide: software for canonical community ordination (version 4.5). Microcomputer Power, Ithaca
Thomas JA, Morris MG, Hambler C (1994) Patterns, mechanisms and rates of extinction among invertebrates in the United-Kingdom. Philos Trans R Soc London Biol 344:47–54
van Dijk G (1991) The status of semi-natural grasslands in Europe. In: Goriup PD, Batten LA, Norton JA (eds) The conservation of lowland dry grassland birds in Europe. Joint Nature Conservation Committee, Peterborough, pp 15–36
Van Dyck H, Van Strien AJ, Maes D, Van Swaay CAM (2009) Declines in common, widespread butterflies in a landscape under intense human use. Conserv Biol 23:957–965
WallisDeVries MF, Poschlod P, Willems JH (2002) Challenges for the conservation of calcareous grasslands in northwestern Europe: integrating the requirements of flora and fauna. Biol Conserv 104:265–273
Wrbka T, Schindler S, Pollheimer M, Schmitzberger I, Peterseil J (2008) Impact of the Austrian agri-environmental scheme on diversity of landscapes, plants and birds. Community Ecol 9:217–227
Acknowledgments
This study originated during the preparation of the management plan for the Babiččino údolí reserve. We would like to thank T. Bury, S. Korinkova and M. Kubiczkova for help with data collection; A. Kodadkova, T. Lamosova, J. J. Michalek and K. Novakova for material sorting; J. Benes and J. Blizek for ground beetles determination; P. Smilauer for help with analyses design; M. Sweney for English corrections; and J. Benes and P. Vrba for valuable comments; and two anonymous referees for useful suggestions. Funding was provided by the Czech Agency for Nature Conservation (PPK-35a/62/06), the Czech Ministry of Education (LC 06073, MSM 6007665801) and Environment (SP/2D3/62/08) and the Czech Science Foundation (208/08/H044).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cizek, O., Zamecnik, J., Tropek, R. et al. Diversification of mowing regime increases arthropods diversity in species-poor cultural hay meadows. J Insect Conserv 16, 215–226 (2012). https://doi.org/10.1007/s10841-011-9407-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-011-9407-6