
Abstract
Grasshoppers could be considered as appropriate ecological indicators for grasslands owing to their sensitive response to environmental features. However, if grasshoppers are a good ecological indicator, they must (i) also represent other taxa, and (ii) provide additional information over straight measurement of environmental variables. To assess this, we compared the congruence of species richness patterns of grasshoppers with butterflies and environmental variables in two areas with extensive ecological networks (ENs). ENs are landscape-scale remnants of corridors and nodes of natural habitat running throughout a transformed, usually agricultural, landscape. Species richness of grasshoppers and butterflies did not differ among reference and EN sites, but guild composition differed significantly. While ENs adequately conserved overall diversity of these two groups, they were utilized preferentially by small-sized grasshoppers and shrub and tree-feeding butterflies. Reference sites had significantly more graminivorous and intermediate-mobility grasshopper species, as well as more butterfly species with widespread distribution, herbaceous dicot feeders and those with no recorded association to forest edges. Nevertheless, grasshopper and butterfly species richness’ were highly correlated. These results were similar across geographic areas, despite the fact that the areas differed significantly in their overall richness and species composition. Although there were some specific significant correlations between environmental variables and diversity, none of the variables could adequately replace use of the insect assemblage for bioindication. We conclude that grasshopper species and guild richness are representative of the butterfly assemblage, and provide information which is not sufficiently clear when utilizing only environmental variables.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Terrestrial invertebrates have been suggested as potential bioindicators for many scenarios and regions of the world (Duelli et al. 1999; Kremen et al. 1993; McGeoch 2007) However, for an organism to be considered a bioindicator, several steps must be followed and certain critical criteria met (Duelli and Obrist 2003; Fleishman and Murphy 2009; McGeoch 2007). Firstly, clarification is required within the specific context, of both the term ‘indicator’ and the specific goals of the proposed indicator. Here, we adopt McGeoch’s (2007) definition of an ecological indicator as “a species or group of species that demonstrates the effect of environmental change (such as habitat alteration, fragmentation and climate change) on biota or biotic systems”. To increase the ‘predictability’ of the indicator, McGeoch (2007) then suggests several additional steps including establishment of the relationship between the indicator and object to be indicated, robustness of this relationship and representativeness of the indicator for other taxa.
Another important factor to take into account during insect bioindicator designation is feasibility, as the process of adequately sampling insects in an ecosystem can be time-consuming and costly (Fleishman and Murphy 2009). In a commercial context, these considerations may determine whether a bioindication system, once developed, is actually applied. McGeoch et al. (2002) address this when they identify an indicator assemblage of dung beetles in South Africa and then validate it on a smaller dataset representative of a collection for monitoring purposes. They also identify ‘detector’ species—indicators of the turnover of habitat along a gradient of disturbance—from mixed woodland to sand forest. However, if this turnover were easily observed in the vegetation composition or structure, then this may preclude the need for a time-consuming survey of the dung beetle assemblage.
Among the arthropods, grasshoppers (Insecta: Orthoptera: Acridoidea) have been suggested as appropriate bioindicators for several regions and biomes (Báldi and Kisbenedek 1997; Kati et al. 2004; Sauberer et al. 2004; Steck et al. 2007). Within South Africa’s grasslands, they have been shown to fulfill the first criteria for an ecological indicator (McGeoch 2007), strong and sensitive response to their habitat (Kinvig 2006), particularly to management practices such as burning (Chambers and Samways 1998), grazing (Gebeyehu and Samways 2003; Rivers-Moore and Samways 1996), and mowing (Chambers and Samways 1998). In this study, we build upon the strength of grasshoppers’ relationships with their habitat, and address two further considerations for their application as bioindicators: whether species diversity and composition is congruent with that of butterflies, and whether their response to habitat can be adequately indicated by measurement of environmental variables.
Butterflies were chosen for this study because in South Africa, they respond to landscape design by flying faster through narrow than wide corridors (Pryke and Samways 2001), and have been shown to have a clear preference for habitat of high quality (Pryke and Samways 2003). A similar study in Greece showed that grasshoppers and butterflies had strongly congruent species richness patterns and that both groups were influenced by common ecological factors (Zografou et al. 2009). To increase the robustness of our findings, we compiled information from two geographic regions with distinct insect assemblages (Fleishman and Murphy 2009). We also included guild-level assemblage characteristics since information may be lost when species identity is not taken into account (Magurran 2004; McGeoch et al. 2002; Pearman and Weber 2007). Certain species are known to benefit from disturbance (Grez et al. 2004; Schulze et al. 2004), and high species diversity does not always correlate with the presence of high conservation value species (Schulze et al. 2004). Application of a species-level bioindicator also presents a problem, as assemblages have been shown to turn over, sometimes as often as every 11 years (Margules et al. 1994). Dividing species into guilds provides an intermediate alternative to these levels (Barbaro and van Halder 2009). Grasshoppers and butterflies were collected and compared within a complex system of remnant grassland corridors and nodes (ecological networks, ENs) within an exotic timber plantation matrix in South Africa.
Ecological networks are landscape-scale systems of remnant or restored natural habitat which are established and managed within transformed, usually agricultural, landscapes (Jongman and Pungetti 2004; Samways 2007). They are intended to provide additional habitat and movement corridors for organisms to navigate altered landscapes, thereby mitigating the effects of this transformation. However, although some landscapes which unintentionally meet these criteria have been in place for centuries, the concept of managing and designing ENs for optimal conservation function is a new one, requiring much more research (Boitani et al. 2007; Rouget et al. 2006).
Commercial, exotic, timber plantations cover 1.1% of South Africa’s land surface area (500,000 ha) (DWAF 2009), mostly within the highly threatened grassland biome (Neke and du Plessis 2004; Reyers et al. 2001). In order to increase the market value of their goods, South African timber companies leave, on average, one-third of all of their holdings permanently unplanted, and these areas form the ENs of grassland running throughout the exotic timber matrix (Jackleman et al. 2006).
Conditions of many aspects of the ENs can be monitored directly (e.g. soil moisture content, mean vegetation height) or by using GIS (e.g. size and isolation of ENs). Since both grasshoppers and butterflies respond sensitively to their environments, then it is conceivable that they could correlate strongly with one or several environmental variables, which could then be monitored instead of the assemblage, if the grasshoppers are found not to provide sufficient unique information to warrant such costly monitoring techniques. Here, we compare the congruence of grasshopper and butterfly species richness patterns to assess representativeness, and then correlate the species richness’ of the two groups with various environmental variables.
We ask the questions: (1) To what extent are ENs effectively conserving diversity of grasshoppers and butterflies? (2) To what extent are grasshopper and butterfly species diversity and composition congruent? (3) Can any environmental variables be identified which correlate with diversity of either taxon or their individual guilds and that could replace the necessity for a bioindicator?
Methods
Study area
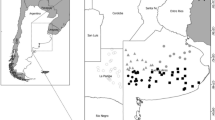
The study region consisted of four exotic timber management units (MUs) in KwaZulu-Natal Province, South Africa, in two geographically distinct regions of the province, Zululand (3 MUs, 28°15′S 32°20′E) and the Midlands (1 MU, 29°25′S 30°30′E), about 200 km apart (Fig. 1, Table 1). MUs were established by Mondi, the commercial paper producer, for logistical and business reasons. Each MU is under the control of a different manager, which may affect the frequency and extent of management actions, as these are heavily reliant on logistical and commercial considerations and do not take top priority (e.g. if weather is unseasonably hot and windy, then prescribed burnings may be delayed, or if there is a pest outbreak damaging commercial trees, then all necessary resources would be diverted to this). The two regions differed in vegetation type: Zululand consisted mostly of Maputaland Wooded Grassland and the Midlands of Drakensberg Foothills Moist Grassland (Mucina and Rutherford 2006), in elevation—Zululand was 10–100 m above sea level and the Midlands was 1,400–1,800 m above sea level—and in topography—Zululand was flat with an elevational range of 90 m and the Midlands was more hilly with an elevation range of 400 m. Additionally, the Zululand MUs consisted of mostly Eucalyptus spp., while the Midlands MU consisted of predominantly Pinus spp. Both types of trees have previously been shown to be detrimental to insect assemblages (Armstrong and van Hensbergen 1996; Ratsirarson et al. 2002; Samways and Moore 1991) and both act as barriers to free insect movement and ensure independence of EN sites at the time scale of the study.

Map of study area in KwaZulu-Natal Province, South Africa, with inlay of two geographical regions (Midlands and Zululand) and four sampled management units (numbered 1–4). Enlarged image of management unit 1 provided for greater detail
The MUs were 7,826, 6,653, 8,595, and 5,241 ha, respectively with 17–49% (mean value 27%) of the MU maintained as conservation areas which made up the ENs. All four MUs were originally planted between 50 and 100 years prior to sampling. Natural grassland remnants varied in age, with some having never been planted and others having been restored to natural conditions within the last 5 years. ENs in the MUs consisted of conservation areas deliberately left unplanted in accordance with regulations instated by South Africa’s Department of Water Affairs and Forestry (DWAF), as well as utility areas such as power line servitudes, fire breaks, dwellings and office complexes.
Three large, unfragmented, natural grassland areas adjacent to the MUs were utilized as reference sites for the purpose of comparing assemblages within ENs to the most likely assemblage outside of the EN (Colwell and Coddington 1994). These were the only available reference sites in the region or more would have been included. At the Midlands MU (MU 4, see Table 1), the reference site was a large grassland on the outskirts of the forests (308 ha) adjacent to the Karkloof Nature Reserve. In Zululand, the two reference sites were the Langepan Vlei (40 ha) home to the only remaining population of Kniphofia leucocephala (Baijnath 1992), a critically endangered plant (for MU 1), and the entrance way to the iSimangaliso Wetland Park (70 ha), a World Heritage Site (for MU 2 and 3).
Grasshopper and butterfly sampling
Thirty-one sites of remnant grassland along with three references sites were chosen for sampling. These sites ranged from 1 to 22 ha and the minimum distance between any two sites was 500 m, separated by exotic timber which was inhospitable to the study organisms and ensured independence of sites. One large quadrat of 50 m × 50 m was delineated in the center of each grassland site, 10 m internally to the edge of the site to avoid the influence of edge effects (Chambers and Samways 1998). This was the maximum quadrat size possible to sample in all sites. Where sites were within power line servitudes, the quadrat dimensions were altered to 125 m × 20 m because of the long, narrow shape of these sites. Three types of sites were identified and are referred to as “site types”: reference sites, continuous sites (i.e. power line servitudes or fire breaks), and isolated patches.
Grasshoppers and butterflies were sampled by CS Bazelet and BN Gcumisa, four times within each site between February and April 2008, between 09:00 and 17:00 on sunny days with low cloud cover, twice in the morning and twice in the afternoon on alternating days to avoid sampling bias. There were no significant differences in species richness among the times of day (AM vs. PM) (Kruskal–Wallis test in SAS Enterprise Guide 4.1: grasshoppers: χ2 = 1.77, P > 0.05; butterflies: χ2 = 1.05, P > 0.05). Grasshopper sampling was completed four times for 30 min each time, by walking through the quadrat and flushing individuals, then actively capturing them with an insect net. Of the four total hours spent sampling, sweep netting was conducted for half an hour to account for the sedentary minority of species. All adult grasshoppers were retained and identified by CS Bazelet to the most recent accepted taxonomic classifications using Dirsh (1965), Eades and Otte (2009), and Johnsen (1984, 1991). Although individuals were counted, for the purpose of this study, values have been reduced to incidence data. Timed counts of large quadrats is an acceptable method for sampling grasshoppers in relatively short grass swards (<50 cm) when they occur in low densities (Gardiner et al. 2005).
Butterflies were sampled in the same quadrats as the grasshoppers to standardize methods. Butterflies were also sampled four times, for 20 min each time by two people. All new species were captured and identified in the field using Woodhall (2005). Individuals that could not be identified in the field and one individual of each new species encountered were stored in a voucher collection. All other individuals were released. Only incidence data were recorded per species per site to avoid pseudoreplication.
Guilds
All species were assigned to guilds based on known life history traits (“Appendix”). The life-history traits chosen were expected to be good predictors of species response to fragmentation (Barbaro and van Halder 2009; Henle et al. 2004). Four life history traits were selected for grasshoppers, and evaluated from a number of sources (Dirsh 1965; Gandar 1983; Johnsen 1984, 1991). Where specific information could not be found (for South African grasshoppers ecological information is particularly scant), we extrapolated from closely-related species. Five butterfly traits were selected and evaluated using a number of sources (Henning et al. 1997; Van Son 1949, 1955, 1963, 1979; Woodhall 2005).
Grasshopper and butterfly distributions were categorized as localized, regional or widespread on the basis of their extent of occurrence in South Africa. Grasshopper trophic guild was designated as graminivorous, mixed-feeder or forb-feeder only (Kinvig 2006). Butterfly feeding guild was evaluated based on the larval food plant as graminivorous, herbaceous dicot-feeders, shrub and tree-feeders, and a single insectivorous species (Barbaro and van Halder 2009). Body size was categorized as small, medium or large. For grasshoppers, we used the classification assigned by Dirsh (1965) for each grasshopper genus, although he did not specify the size range of each of the three categories. For butterflies, we categorized size as: <25 mm (small), 25–50 mm (medium), or >50 mm (large). Mobility was low, medium or high for both groups based on their flight capabilities and migratory patterns. Grasshoppers of the Oedipodinae subfamily are known to be strong fliers (Ritchie 1981) so were considered of high mobility. Apterous lentulids, brachypterous and known sedentary grasshopper species were considered to be of low mobility. For butterflies, migratory and strong-flying species were classified as high mobility species. Species which were described in the literature as poor fliers, were classified as low mobility species. All grasshopper and butterfly species which did not fall into any of these categories were classified as having intermediate mobility. Butterfly habitat affinity was categorized into three groups: species with a known association with forest edges, no recorded association with forest edges, and wetland species. Association with forest edges could play an important role in these ENs. The same was not done for grasshoppers as these data have never been recorded, and because, although there are species with an affinity for edge habitat, we do not expect any grasshopper species to respond positively to forest as they are grassland specialists (Armstrong and van Hensbergen 1996).
Environmental variables
Three groups of environmental variables were quantified (Table 2). Vegetation variables were related to the internal characteristics of the sites. Grasshopper species composition responds more strongly to vegetation structure than to grass species composition (Gandar 1982; Hochkirch and Adorf 2007; Joern 2005), and therefore only structural diversity of vegetation was surveyed. Vegetation type was measured along ten, 5 m transects, placed at random throughout each site, internally to the 30 m edge (Bullock and Samways 2006). Ground cover was grouped into four categories: tall grasses (>30 cm), short grasses (<30 cm), non-grass vegetation (e.g. shrubs or herbaceous vegetation), or bare ground. Proportion of each vegetation category for all ten transects per site was calculated. Vegetation height was measured at thirty random points throughout the site.
Landscape variables related to the structure of the EN were calculated using ArcGIS 9, digital maps and aerial photographs. Management variables were calculated from surveys completed by managers of each of the MUs. Data were verified with other forest personnel and personal observation to standardize answers given by different managers. Prescribed burning is a necessary management practice within these grasslands, as they naturally succeed to indigenous forest. Most EN sites are slotted for annual or biennial burning, with biennial burns considered optimal from a conservation perspective (Lipsey and Hockey 2010), but managers did not always strictly adhere to this schedule for a variety of logistical reasons.
Statistical analysis
In order to standardize between different organisms, sampling methods and sampling efforts, we compared total species richness per site. Species accumulation curves were plotted for grasshoppers and butterflies using EstimateS and 50 repetitions (Colwell 2009). Typically for many field studies (Schulze et al. 2004), these curves did not reach an asymptote, and therefore we employed the Chao2 species richness estimator (Chao 2005; Colwell and Coddington 1994). This estimator derives the minimum estimate of richness, is appropriate for incidence data, and performs well in many situations, including when the estimator itself has not reached an asymptote, as was the case for butterflies in this study (Colwell and Coddington 1994; Longino et al. 2002; Magurran 2004).
Differences in grasshopper and butterfly assemblages among regions, MUs, site types and sites were assessed as follows: when species richness’ of both taxa were taken together, data were normally-distributed and factorial ANOVA in Statistica 9 determined significant differences in species richness among taxa and levels. When species richness’ of the two taxa were compared separately, data were non-normally distributed and Kruskal–Wallis tests were used to compare differences within taxa. Whittaker’s measure (βW) was used to assess β diversity of raw incidence data. This measure was chosen because it consistently performed better than other β diversity measures (Magurran 2004; Wilson and Shmida 1984). Its values range from 0 (no turnover) to 1 (each sample has a unique set of species) (Magurran 2004) and it was calculated by hand and plotted in Excel 2007.
We compared two square matrices using a Mantel’s test in the ade4 package in R (Dray and Dufour 2007): (1) pairwise β diversity among each pair of sites, with (2) geographical (Euclidean) distance between each pair of sites. Raw species richness of each reference site was also compared with that of its associated sites only using a Mann–Whitney U test in Statistica 9. Box and whisker plots were prepared using values from Statistica 9 imported into Excel 2007. Species richness of individual guilds in reference sites versus non-reference sites was compared using a Kruskal–Wallis test in Statistica 9, as well.
Estimated species richness and observed species richness were found to be highly, significantly correlated. So grasshopper and butterfly raw and guild-level species richness’ and environmental variables were correlated using Spearman’s correlation for non-parametric data in Statistica 9. Finally, linear regressions were plotted in SAS Enterprise Guide 4 to determine whether guilds with higher species richness were more likely to produce statistically significant Spearman’s correlations.
Results
Descriptive data
Overall, 61 species of grasshoppers and 55 species of butterflies were collected (Table 1). Neither group reached an asymptote overall or within any individual MU. Grasshopper sampling was more complete than that for butterflies (78% completeness vs. 49% completeness overall). Within the four MUs, grasshopper completeness ranged from 80 to 96%, while butterfly completeness ranged from 36 to 78%. The Chao2 estimate was significantly, positively correlated with raw species richness for both groups (grasshoppers: R 2 = 0.53; P < 0.0001; butterflies: R 2 = 0.58; P < 0.0001).
The Midlands region had significantly higher species richness of grasshoppers and butterflies than Zululand (grasshoppers: H = 10.81, P = 0.001; butterflies: H = 6.26, P = 0.012). The four MUs were also significantly different from each other with the highest species richness in the Midlands MU (MU-4) (grasshoppers: H = 12.17, P = 0.007; butterflies: H = 14.12, P = 0.003). The site types did not have significantly different species richness’ (grasshoppers: H = 2.37, P = 0.307; butterflies: H = 2.14, P = 0.343) (Fig. 2). This was also true when species richness of each reference site was compared with its associated sites only (grasshoppers: UMU1 = −0.15, P = 0.880; UMU2,3 = 0.76, P = 0.450; UMU4 = 1.22, P = 0.224; butterflies: UMU1 = 1.20, P = 0.229; UMU2,3 = −0.47, P = 0.640; UMU4 = 0.17, P = 0.860).

Results of factorial ANOVA of differences among regions (a), among site types (b), and among management units (c). Letters indicate significant differences and bars indicate 95% confidence intervals
However, there were significant differences between reference sites and all other sites, when comparing individual guilds. Graminivorous grasshoppers and those with intermediate mobility were more speciose in reference sites (H = 4.55, P = 0.033; H = 4.76, P = 0.029, respectively), while small grasshoppers were more speciose in non-reference sites (H = 4.11, P = 0.043). Butterflies with widespread distribution, herbaceous dicot feeders, and those with no recorded association to forest edges were on average more speciose in reference sites (H = 3.96, P = 0.047; H = 4.31, P = 0.038, H = 9.60, P = 0.002, respectively), while shrub and tree-feeders were more speciose in non-reference sites (H = 4.83, P = 0.028).
Similarly, β diversity was greatest between regions (grasshoppers: βW = 0.74; butterflies: βW = 0.75) (Fig. 3). Median turnover decreased from regions to MUs to site types, with grasshoppers and butterflies sharing similar values. Among sites, median turnover was large (grasshoppers: βW = 0.67; butterflies: βW = 0.80) with a large variance among values. Mantel’s tests showed that the matrix of β diversity values for each pair of sites was significantly correlated with the distance between the sites for both grasshoppers and butterflies (grasshoppers: Mantel’s r = 0.67, P = 0.001; butterflies: Mantel’s r = 0.38, P = 0.001).

Comparison of β diversity among regions, plantations, site types and sites. Horizontal bar indicates median value, top and bottom of box indicate 1st and 3rd quartiles respectively, and vertical bars indicate minimum and maximum values. Grey shaded boxes, and horizontal bar with “x” indicate grasshopper scores. White boxes and horizontal bar with no “x” indicate butterfly scores
Correlations
Grasshopper Chao2 estimated species richness was significantly correlated with butterfly Chao2 estimated species richness (r = 0.37, P = 0.035) (Table 3). Conservation area within 500 m surrounding the site was the only environmental variable significantly correlated with grasshopper and with butterfly total species richness (r = 0.36, P = 0.036; r = 0.37, P = 0.035, respectively).
Of the grasshopper guilds, those with localized distribution were significantly correlated with the greatest number of butterfly guilds (n = 6), and also with total butterfly estimated species richness (Table 3). Medium-sized grasshoppers were correlated with richness of five butterfly guilds, while small-sized grasshoppers were significantly correlated with six of the environmental variables. Wetland-associated and intermediate mobility butterfly guilds were each significantly positively correlated with six grasshopper guilds as well as with total grasshopper species richness.
The proportion of short grasses at a site was correlated with the greatest number of insect guilds: three butterfly and four grasshopper guilds (Table 3, 4). Conservation area within 500 m surrounding a site followed closely with six insect guilds, three butterfly and three grasshopper guilds. Overall, there were nine significant correlations between grasshopper guilds and vegetation variables (n = 5), five landscape variables (n = 4), and five management variables (n = 3). There were seven significant correlations between butterfly guilds and vegetation variables, seven with landscape variables and three with management variables. All correlations between grasshoppers and butterflies were positive, while several environmental variables were negatively correlated with guilds. Speciose guilds were not more likely to correlate significantly (grasshoppers: R 2 = 0.03, P > 0.05; butterflies: R 2 = 0.09, P > 0.05).
Discussion
Response to ecological networks
The similarity in species richness between the extensive, natural reference sites and the EN sites indicated how effective ENs were for maintaining the local species richness. This was true when classifying sites by their structural type (reference, continuous or patchy) and when the richness of each reference site was contrasted with its associated sites only.
However, arguably more important than this are the species identities in the ENs. Natural reference sites had only a few unique species (Table 1). Of the grasshoppers, only three species of 61 were unique to reference sites, and butterflies had only one unique species to the reference sites. In terms of compositional biodiversity and the finer guild scale, there were some small differences between reference and EN sites. Three grasshopper guilds and four butterfly guilds had significantly different richness in reference vs. EN sites. Of these, all but two were more speciose in the reference sites. ENs may be beneficial to small grasshoppers because they are sufficiently expansive for these small insects to meet their home range requirements, while providing shelter from larger, predatory organisms which would utilize the small grasshoppers as a food source but whose home ranges necessitate larger areas than those provided by the ENs. Additionally, shrub and tree-feeding butterflies would naturally prefer ENs which contain a greater diversity of their larval food plants than the reference sites which are managed optimally for grassland and are therefore devoid of shrubs and trees.
Nevertheless, overall ENs appeared to adequately conserve grasshoppers and butterflies. These findings corroborate those of Bullock and Samways (2006) who found that insects are capable of utilizing fragmented grassland as long as their host plant subsists within the fragments. Field (2002) found a slight decrease in some native pollinators within grassland fragments, but the remnant assemblage was probably sufficient to ensure the long-term survival of native wildflowers, whose assemblage persisted on a smaller scale. Butterflies have previously been found to utilize grassland fragments preferentially depending on the level of disturbance within the fragment, the size of the fragment (or width if it is a corridor) and characteristics of the butterfly species (e.g. mobility and distribution) (Pryke and Samways 2001, 2003). Our findings do not corroborate these as we would expect to see marked differences between reference and EN sites were this true. Perhaps, species level analysis is necessary in order to detect these differences. Other organisms may not utilize the ENs as successfully as grasshoppers and butterflies, for example grassland birds have reduced assemblages in ENs (Lipsey and Hockey 2010) with certain species requiring large, unfragmented grasslands similar to the reference sites, to thrive.
Our results also lend support to previous studies which have shown that species richness or other diversity—index based measures which do not take into account species composition, are insufficient for assessing habitat quality (e.g. McGeoch et al. 2002). When we looked at species assemblage at a slightly finer scale of guild level, already differences became apparent between sites which were masked when assessing species richness alone.
Congruence
There was a strong relationship between grasshopper and butterfly species richness in ENs in total, and also at guild level. At guild level, there were numerous positive correlations, although similar guilds did not correlate with each other (for instance graminivores of grasshoppers and butterflies did not correlate), with the exception of intermediate and high mobility grasshoppers and butterflies, which did correlate. Similarly to Zografou et al. (2009), our study found no negative correlations between the two taxa.
Representative value of environmental variables as measured in this study was minimal in comparison to that of the taxa themselves. In most cases, those guilds with a large number of correlations to environmental variables were not the same guilds with significant correlations to guilds of the other taxon (with the exception of wetland-associated butterflies which correlated with numerous grasshopper guilds and with environmental variables). This illustrates that, as expected, species richness of grasshoppers and butterflies reflects a complicated response to numerous variables, not all of which can be detected or measured with confidence.
Environmental variables can be quantified by managers and forest personnel as they were in this study, but here we see that no single variable can provide adequate information regarding the status of the resident species. Species composition of insects in ENs reflects a complex response to variables beyond our immediate comprehension. The positive correlation of overall species richness for grasshoppers and butterflies with area of natural habitat in the vicinity, indicates the importance of connectivity of EN sites for insects. However, beyond this, in order to assess the state of biotic assemblages within ENs, no environmental measure gives as complete a picture as collections of the insects themselves, as different species and groups with different traits respond differently to ongoing processes within ENs.
In conclusion, grasshopper bioindicators provide information on the status of biotic communities within ENs. Their response to ENs is representative of that of butterflies. No environmental variable of the ENs can adequately replace information obtained by monitoring of grasshopper bioindicators. Environmental variables should be measured complementarily to the grasshopper bioindicator as the two provide different important information for management of ENs.
References
Armstrong AJ, van Hensbergen HJ (1996) Impacts of afforestation with pines on assemblages of native biota in South Africa. S Afr Forest J 175:35–42
Baijnath H (1992) Kniphofia leucocephala (Asphodelaceae): a new white-flowered red-hot poker from South Africa. S Afr J Bot 58:482–485
Báldi A, Kisbenedek T (1997) Orthopteran assemblages as indicators of grassland naturalness in Hungary. Agr Ecosyst Environ 66:121–129
Barbaro L, van Halder I (2009) Linking bird, carabid beetle and butterfly life-history traits to habitat fragmentation in mosaic landscapes. Ecography 32:321–333
Boitani L, Falcucci A, Maiorano L, Rondinini C (2007) Ecological networks as conceptual frameworks or operational tools in conservation. Conserv Biol 21:1414–1422
Bullock WL, Samways MJ (2006) Conservation of flower-arthropod interactions in remnant grassland linkages among pine afforestation. Biodivers Conserv 14:3093–3103
Chambers BQ, Samways MJ (1998) Grasshopper response to a 40-year experimental burning and mowing regime, with recommendations for invertebrate conservation management. Biodivers Conserv 7:985–1012
Chao A (2005) Species richness estimation. In: Encyclopedia of statistical sciences. Wiley, New York, pp 7909–7916
Colwell RK (2009) Estimates: statistical estimation of species richness and shared species from samples. Version 8.2. user’s guide and application. http://purl.oclc.org/estimates. Accessed 9 March 2010
Colwell RK, Coddington JA (1994) Estimating terrestrial biodiversity through extrapolation. Philos Trans Biol Sci 345:101–118
Dirsh VM (1965) The African genera of Acridoidea. Anti-locust research centre at the University Press, Cambridge, UK
Dray S, Dufour AB (2007) The ade4 package: implementing the duality diagram for ecologists. J Stat Softw 22:1–20
Duelli P, Obrist MK (2003) Biodiversity indicators: the choice of values and measures. Agr Ecosyst Environ 98:87–98
Duelli P, Obrist MK, Schmatz DR (1999) Biodiversity evaluation in agricultural landscapes: above-ground insects. Agr Ecosyst Environ 74:33–64
DWAF (Department of Water Affairs and Forestry) (2009) Abstract of South African forestry facts for the year 2007/2008. http://www2.dwaf.gov.za/webapp/Documents/FSA-Abstracts2009.pdf. Accessed 3 Sept 2010
Eades DC, Otte D (2009) Orthoptera species file Online. Version 2.0/3.5. http://Orthoptera.SpeciesFile.org. Accessed 3 Sept 2010
Field LF (2002) Consequences of habitat fragmentation for the pollination of wildflowers in moist upland grasslands of KwaZulu-Natal. M.Sc. Thesis. University of KwaZulu-Natal, Pietermaritzburg
Fleishman E, Murphy DD (2009) A realistic assessment of the indicator potential of butterflies and other charismatic taxonomic groups. Conserv Biol 23:1109–1116
Gandar MV (1982) The dynamics and trophic ecology of grasshoppers (Acridoidea) in a South African savanna. Oecologia 54:370–378
Gandar MV (1983) Ecological notes and annotated checklist of the grasshoppers (Orthoptera: Acridoidea) of the Savanna Ecosystem Project Study Area, Nylsvley
Gardiner T, Hill JK, Chesmore D (2005) Review of the methods frequently used to estimate the abundance of Orthoptera in grassland ecosystems. J Insect Conserv 9:151–173
Gebeyehu S, Samways MJ (2003) Responses of grasshopper assemblages to long-term grazing management in semi-arid African savanna. Agr Ecosyst Environ 95:613–622
Grez A, Zaviezo T, Tischendorf L, Fahrig L (2004) A transient, positive effect of habitat fragmentation on insect population densities. Oecologia 141:444–451
Henle K, Davies KF, Kleyer M, Margules C, Settele J (2004) Predictors of species sensitivity to fragmentation. Biodivers Conserv 13:207–251
Henning GA, Henning SF, Joannou JG, Woodhall SE (1997) Living butterflies of southern Africa: biology, ecology and conservation. Volume 1: Hesperidae, Papilionidae and Pieridae of South Africa. Umdaus Press, Hatfield
Hochkirch A, Adorf F (2007) Effects of prescribed burning and wildfires on Orthoptera in Central European peat bogs. Environ Conserv 34:225–235
Jackleman J, Wistebaar N, Rouget M, Germishuizen S, Summers R (2006) An assessment of the unplanted forestry land holdings in the grasslands biome of Mpumalanga, KwaZulu-Natal and the Eastern Cape. Grasslands: living in a working landscape. Report No. 8. South African National Biodiversity Institute - Grasslands Programme, Pretoria
Joern A (2005) Disturbance by fire frequency and bison grazing modulate grasshopper assemblages in tallgrass prairie. Ecology 86:861–873
Johnsen P (1984) Acridoidea of Zambia. Aarhus University Zoological Laboratory, Aarhus
Johnsen P (1991) Acridoidea of Botswana. Aarhus University Zoological Laboratory, Aarhus
Jongman RHG, Pungetti G (2004) Ecological networks and greenways—concept, design and implementation. Cambridge University Press, Cambridge
Kati V, Devillers P, Dufrêne M, Legakis A, Vokou D, Lebrun P (2004) Testing the values of six taxonomic groups as biodiversity indicators at a local scale. Conserv Biol 18:667–675
Kinvig R (2006) Biotic indicators of grassland condition in KwaZulu Natal with management recommendations. Ph.D. Dissertation. University of KwaZulu-Natal, Pietermaritzburg
Kremen C, Colwell RK, Erwin TL, Murphy DD, Noss RF, Sanjayan MA (1993) Terrestrial arthropod assemblages: their use in conservation planning. Conserv Biol 7:796–808
Laurance WF, Yensen E (1991) Predicting the impacts of edge effects in fragmented habitats. Biol Conserv 55:77–92
Lipsey MK, Hockey PAR (2010) Do ecological networks in South African commercial forests benefit grassland birds? Agr Ecosyst Environ 137:133–142
Longino JT, Coddington J, Colwell RK (2002) The ant fauna of a tropical rain forest: estimating species richness three different ways. Ecology 83:689–702
Magurran AE (2004) Measuring biological diversity. Blackwell, Malden
Margules CR, Nicholls AO, Usher MB (1994) Apparent species turnover, probability of extinction and the selection of nature reserves—a case study of the Ingleborough limestone pavements. Conserv Biol 8:398–409
McGeoch MA (2007) Insects and bioindication: theory and progress. In: Insect conservation biology. CABI, Wallingford, pp 144–174
McGeoch MA, van Rensburg BJ, Botes A (2002) The verification and application of bioindicators: a case study of dung beetles in a savanna ecosystem. J Appl Ecol 39:661–672
Mucina L, Rutherford MC (2006) The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute, Pretoria
Neke KS, du Plessis MA (2004) The threat of transformation: quantifying the vulnerability of grasslands in South Africa. Conserv Biol 18:466–477
Pearman PB, Weber D (2007) Common species determine richness patterns in biodiversity indicator taxa. Biol Conserv 138:109–119
Pryke SR, Samways MJ (2001) Width of grassland linkages for the conservation of butterflies in South African afforested areas. Biol Conserv 101:85–96
Pryke SR, Samways MJ (2003) Quality of remnant indigenous grassland linkages for adult butterflies (Lepidoptera) in an afforested African landscape. Biodivers Conserv 12:1985–2004
Ratsirarson H, Robertson HG, Picker MD, van Noort S (2002) Indigenous forests versus exotic eucalypt and pine plantations: a comparison of leaf-litter invertebrate communities. Afr Entomolo 10:93–99
Reyers B, Fairbanks DHK, van Jaarsveld AS, Thompson M (2001) Priority areas for the conservation of South African vegetation: a coarse-filter approach. Divers Distrib 7:79–95
Ritchie JM (1981) A taxonomic revision of the genus Oedaleus Fieber (Orthoptera: Acrididae). Bull Br Mus (Nat Hist) Entomo 42:83–183
Rivers-Moore NA, Samways MJ (1996) Game and cattle trampling, and impacts of human dwellings on arthropods at a game park boundary. Biodivers Conserv 5:1545–1556
Rouget M, Cowling RM, Lombard AT, Knight AT, Kerley GIH (2006) Designing large-scale conservation corridors for pattern and process. Conserv Biol 20:549–561
Samways MJ (2007) Implementing ecological networks for conserving insect and other biodiversity. In: Insect conservation biology. CABI, Wallingford, pp 127–143
Samways MJ, Moore SD (1991) Influence of exotic conifer patches on grasshopper (Orthoptera) assemblages in a grassland matrix at a recreational resort, Natal, South Africa. Biol Conserv 57:117–137
Sauberer N, Zulka KP, Abensperg-Traun M, Berg H-M, Bieringer G, Milasowszky N, Moser D, Plutzar C, Pollheimer M, Storch C, Trostl R, Zechmeister H, Grabherr G (2004) Surrogate taxa for biodiversity in agricultural landscapes of eastern Austria. Biol Conserv 117:181–190
Schulze CH, Waltert M, Kessler PJA, Pitopang R, Shahabuddin, Veddeler D, Mühlenberg M, Gradstein R, Leuschner C, Steffan-Dewenter I, Tscharntke T (2004) Biodiversity indicator groups of tropical land-use systems: comparing plants, birds, and insects. Ecol Appl 14:1321–1333
Steck CE, Burgi M, Coch T, Duelli P (2007) Hotspots and richness pattern of grasshopper species in cultural landscapes. Biodivers Conserv 16:2075–2086
Van Son G (1949) The butterflies of southern Africa. Part I: Papilionidae and Pieridae. Transvaal Museum Memoir No. 3, Pretoria
Van Son G (1955) The butterflies of southern Africa. Part II: Nymphalidae: Danainae and Satyrinae. Transvaal Museum Memoir No. 8, Pretoria
Van Son G (1963) The butterflies of southern Africa. Part III: Nymphalidae: Acraeinae. Transvaal Museum Memoirs No. 14, Pretoria
Van Son G (1979) The butterflies of southern Africa. Part IV: Nymphalidae: Nymphalinae. Transvaal Museum Memoirs No. 22, Pretoria
Wilson MV, Shmida A (1984) Measuring beta diversity with presence absence data. J Ecol 72:1055–1064
Woodhall S (2005) Field guide to butterflies of South Africa. Struik, Durban
Zografou K, Sfenthourakis S, Pullin A, Kati V (2009) On the surrogate value of red-listed butterflies for butterflies and grasshoppers: a case study in Grammos site of Natura 2000, Greece. J Insect Conserv 13:505–514
Acknowledgments
We thank the Mauerberger Foundation Fund and RUBICODE for funding. Mondi, Mondi Shanduka, SiyaQhubeka Forestry, iSimangaliso Wetland Park and EKZN Wildlife provided accommodation in the field, as well as insect collection permits and permission to sample on their holdings. B. Gcumisa provided field assistance. GIS maps were supplied by D. van Zyl. M. Masango, C. Burchmore, P. Gardner, N. Mkhize, L. Nel, R. Muller, G. Kruger and A. Madikane provided technical assistance for this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bazelet, C.S., Samways, M.J. Grasshopper and butterfly local congruency in grassland remnants. J Insect Conserv 16, 71–85 (2012). https://doi.org/10.1007/s10841-011-9394-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-011-9394-7