
Abstract
The migration of monarch butterflies (Danaus plexippus) from Canada and the United States to overwintering sites in Mexico is one of the world’s most amazing biological phenomena, although recent threats make it imperative that the resources needed by migrating monarchs be conserved. The most important first step in preserving migration resources—determining the migration flyways—is also the most challenging because of the large-scale nature of the migration. Prior attempts to determine the flyways using mark-recapture techniques with wing tags gave some clues, but this important information has never been fully obtainable until now. In 2005 the citizen-science program, Journey North, initiated a project that asked participants to record sightings of overnight roosts of monarchs during their fall migration, and this project now provides an ideal way to illustrate the flyways used by monarchs on their way to Mexico, with the assumption that roost locations indicate migration routes. We used 3 years of this data to elucidate the flyways on a continent-wide scale, that revealed two distinct flyways, but only one appears to lead directly to Mexico. This main, ‘central’ flyway begins in the American Midwest states and southern Ontario, then continues south-southwest through the states of Kansas, Missouri, Oklahoma and Arkansas, and finally passes through Texas and northern Mexico. These data also highlighted a separate, smaller flyway along the eastern and coastal states, but there was a noticeable lack of roost sightings in this flyway at lower latitudes. Since there are few recoveries of marked monarchs in Mexico originating from coastal areas, we compared the timing of roost formation in this ‘eastern’ flyway with the main, central flyway. Roosts in the eastern flyway lagged behind the central roosts in timing, suggesting that monarchs traveling in this flyway have a reduced chance of making it to the Mexico wintering site. Combined, our evidence indicates that locations in the central flyway should be considered priority areas for conserving migration resources.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Monarch butterflies (Danaus plexippus) in eastern North America undertake the longest insect migration in the world, traveling each fall as much as 3000 km from breeding areas in Canada, to a few select overwintering sites in the high-altitude mountains of Mexico (Brower 1995). These wintering colonies were first discovered in 1975 (Urquhart 1976) and since then, this amazing journey has been the focus of considerable scientific research (reviewed in Brower 1995, 1996). Today, monarchs in eastern North America face a multitude of threats including loss of habitat in their wintering grounds to illegal logging and changing climate (Oberhauser and Peterson 2003), loss of breeding habitat to urbanization and agriculture, especially genetically modified crops (Oberhauser et al. 2001; Oberhauser and Rivers 2003), and chemical spraying for pest insects (Oberhauser et al. 2006). Importantly, adult monarch butterflies also require resources during their fall migrations, for which little is known scientifically. Each fall, their journey begins in August, and monarchs can be seen flying southward throughout much of Canada and the northern United States, often in large numbers (Moskowitz et al. 2001) during this period. However, since they do not fly at night, monarchs must stop along the way throughout this journey to rest or nectar, and these stopover sites are an important resource for successful monarch migration (Davis and Garland 2004; Brower et al. 2006). At these sites, monarchs settle on trees or other vegetation for the night, sometimes in tight clusters, and these overnight clusters are generally considered ‘roosts’ (Fig. 1). Ironically, despite recent progress in understanding aspects of this unique insect migration (Meitner et al. 2004; Stalleicken et al. 2005; Walton et al. 2005; Gibbs et al. 2006), the places where monarchs choose to stop and roost as well as the locations of major flyways remain unknown, except for a select few sites where fall accumulations are common (Garland and Davis 2002). It is therefore of paramount importance to the conservation of this unique butterfly that the locations of important stopover areas be determined, as well as the flyways that they use most heavily.

Photographs submitted to Journey North of monarch butterflies in overnight roosts in Enid, OK (a), Watertown, SD (b), Texas City, TX (c) and Bolivar, TX (d)
Determining the locations of the migration flyways is not an easy task. Prior attempts involving tag recoveries (whereby monarchs are ‘tagged’ with adhesive, numbered stickers by volunteers) revealed some information regarding possible routes of travel (Urquhart and Urquhart 1977, 1978, 1979b), although the recaptures in these studies provided only straight lines on a map from point of release to point of recovery. With this caveat in mind, these studies appeared to show that monarchs travel from areas such as southern Ontario and Minnesota in a general south-southwest direction. Other important information can also be drawn from current volunteer tagging programs, such as MonarchWatch (www.monarchwatch.org), and the recovery rates of tagged monarchs in the Mexican overwintering sites. For example, Garland and Davis (2002) examined Mexican recovery rates of monarchs tagged at a site in coastal Virginia and found they were much lower than the normal recovery rate of approximately 1%. Moreover, recent work examining recovery rates of other east coast tagging sites has also found extremely low Mexico recovery rates (Brindza et al. in review). Thus, evidence to date indicates that for some reason, monarchs that migrate along the east coast of North America are less successful at reaching the Mexico wintering colonies.
Citizen science programs allow using large numbers of observers to address large scale issues over a broad geographic area. They also offer scientists a way to gather data not obtainable by normal scientific methods. In particular, Journey North is an online citizen-science program (http://www.learner.org/jnorth/) whereby volunteers record sightings of natural history events such as the first adult in their location each spring, and this information is plotted online to show the spring migration of monarchs (for animated maps drawn from sightings see http://www.learner.org/jnorth/monarch/index.html). The majority of participants in the program are students from primary through secondary schools (Howard and Davis 2004). While the goal of these projects is public education, the spring monarch migration data has proven an ideal way to document the spatial and temporal occurrence of monarchs each spring on a continent-wide scale (Howard and Davis 2004; Davis and Howard 2005).
While the prior focus of the monarch program was the spring migration, in 2005, a new Journey North program was established that asked participants to record sightings of overnight roosts of monarchs in the fall. In this program, when participants observe monarchs roosting in trees, bushes etc., they enter the date and location of the sighting into the online database, and these sightings are then plotted online so that the temporal and spatial progression of the fall migration can be viewed, similar to that of the spring migration progression. This program offers a unique opportunity to document on a large scale, the fall migration flyway(s) taken by monarchs in North America, since places where roosts form undoubtedly represent locations where large numbers of monarchs stopover. This data set also allows for interesting comparisons of certain aspects of the flyways such as the pace of migration in each. We used three years of fall roost data from this program in the current paper to address these issues.
Materials and methods
Journey North participants
The Journey North program has been described previously (Howard and Davis 2004). Briefly, the roost sightings project asks volunteers to record when they observe an overnight roost formation each fall, as well as the roost location (the address of the participant, if the roost occurred on their property, or the nearest town). For the purposes of the Journey North program, a general definition of a roost is outlined on the website; i.e. the gathering must contain at least 12 butterflies to be considered a roost (Fig. 1). It is also made clear that a large number of butterflies that have gathered at a nectar source during the day is not considered a roost. Observers are not required to watch the butterflies to establish if they spend the night (it is assumed the butterflies are roosting if they are seen late in the day or early in the morning, and/or they are inactive). More information about typical roosting behavior is available on the website. The participants are not required to report the size of the roost, although there is an optional space online for recording notes on the roosts. Finally, the roost sightings project begins in the third week of August and continues through to the end of October each year.
Data handling
The Journey North staff previews all observations prior to inclusion into the database each year, to minimize the possibility of reports of non-monarchs. However, since most participants in this project also participate in the spring monarch project, and since it is very difficult to mistake a large number of bright orange butterflies with any other species, we are confident that the Journey North sightings are valid occurrences of monarch roosts. In addition, when sightings are verified the staff records the latitude and longitude of the sighting in the database.
Flyway depiction and data analysis
We used the roost sightings data from the first 3 years of this project (2005–2007) for the current study. Our first goal was to graphically present the roost observations so that the migration flyway(s) would be depicted. For this, we pooled the roost data from all 3 years and plotted these points on a map of North America using the latitude and longitude associated with the sightings (Fig. 2). Animated, color maps of these sightings can be viewed online at the Journey North website (http://www.learner.org/jnorth/), and these online maps allow for the temporal occurrence of roosts to be seen, but only for one year at a time. Here, when we initially plotted the roost sightings in Fig. 2 we noticed two distinct groupings of roosts, which we consider to represent separate flyways. The largest by far appears as the group that traverses the center of the breeding range, and we term this the ‘central’ flyway. A second, smaller flyway is visible along the Northeast, mid-Atlantic and coastal plain states, and appears to stop near the Carolinas (or at least no more roosts form), and we term this the ‘eastern’ flyway.

Map of Journey North roost sightings from all years combined (2005–2007). Dashed line indicates division of central and eastern flyways in analysis. Roosts in Florida were not included in the analyses. Inset map shows the locations of all Journey North participants from 1997 to 2007. Star indicates location of Mexico overwintering sites
While it was not the primary goal of this study, these two apparently distinct flyways offered us an opportunity to explore possible differences in the characteristics of both, specifically the timing of roost formation, which we can consider a proxy for the southward migration ‘wave front’ in much the same as the northward wave front in the spring migration can be inferred from adult sightings (Davis and Howard 2005). Thus, we separated the combined roost data by location, so that roosts were categorized as in the ‘central’ or ‘eastern’ flyways. We point out that these groupings are arbitrary, and based solely on our own interpretation of the roost map. Indeed, these groupings may change with additional data from this project, however their distinction here is convenient for statistical comparisons. We further grouped the data based on the time of the fall season the roost formed, by subdividing the data into 10 week intervals. We then used general linear model analyses to explore the factors influencing the timing of roost formation. For this, latitude was the dependent variable, while independents included week, year, and flyway (central versus eastern). All two-way interactions effects were also included. This analysis therefore asked do the average latitudes of roost locations vary with week (which we expect, given the southward progression of the migration); do they vary between years; and importantly, do they differ between flyways? Furthermore, the interaction effect of flyway and week was also important here, since it considered whether the timing of roost formation differs between flyways. We note here that data from the last two weeks were not included in this analysis because only one roost from the eastern flyway was recorded then. In addition, three roost sightings occurred in Florida (Fig. 2), but were not included in analyses. Our analysis in the study was performed using Statistica 6.1 (Statistica 2003).
Results
Data set
There were a total of 635 observations of roosts from the three years of data considered here (Table 1). During each week, the number of reports varied from a low of 4 (though this was at the end of the season) to a high of 36. The number of reports in the last year was considerably higher than the two prior years, though we do not know if this represents more roosts in 2007 or simply more participants in the program.
Flyway description
Plotting the locations of all roosts appeared to reveal two flyway patterns in North America (which we termed central and eastern, Fig. 2), and we grouped the data according to these patterns. We also noted the distinct absence of roosts in the southern states of Alabama, Louisiana and Mississippi, despite the fact that Journey North participants (at least those who participate in the spring project) live there (Fig. 2, inset map). Interestingly, there were three roosts sighted in northern Florida, although we did not consider these as part of the eastern flyway.
Comparison of flyway timing
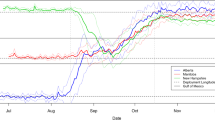
Our roost latitude analysis comparing the two flyways showed that the latitude of roosts varied significantly with week, as we expected (Table 2), but the interaction effect of week × year was significant, meaning that this temporal effect was not consistent among years. There was a significant effect of flyway, and importantly, a significant flyway × week interaction, which is shown in Fig. 3. This graph demonstrates that the roost latitudes of the central flyway progress steadily southward each week, but after the first three weeks the eastern roosts do not. By the middle of the migration season (the week of Sept. 20–26), the trajectory of the eastern roosts ‘diverged’ from the central and essentially leveled out at approximately 40° latitude. Thus, the roosts in the eastern flyway continued to be sighted by Journey North participants at relatively high latitudes long after the central roosts had reached lower latitudes. We point out that this temporal discrepancy can also be visualized on the Journey North website by viewing the animations of the fall roost sightings (http://www.learner.org/jnorth/images/graphics/monarch/maps/Animation_Fall2007_Roosts.html), and noting the colors of the eastern roosts. Finally, the non-significant effect of year × flyway indicates that there is little change in this pattern from year to year.

Average latitudes of monarch roosts each week during the fall (all years combined), grouped by flyways (central and eastern). Error bars represent standard deviations. See Table 1 for dates of each week
Discussion
This study shows, for the first time in clear detail and on a continent-wide scale, the fall migration flyway of monarch butterflies in eastern North America, based on citizen scientists’ observations of overnight roosts. This description is a considerable advancement over prior attempts to document the migration routes using recaptures of tagged monarchs (Urquhart and Urquhart 1977, 1978), and it provides much-needed information regarding priority areas for migration habitat conservation efforts. Specifically, these data show the existence of one main (‘central’) flyway that begins in the American Midwest states and southern Ontario, then progresses south-southwest through the states of Kansas, Missouri, Oklahoma and Arkansas, appearing to narrow along the way until it reaches Texas and finally Northern Mexico, where our roost observations end. These areas should be given highest priority for habitat preservation efforts to ensure adequate migration resources are conserved for the monarch butterfly. These data also appear to show the existence of a second flyway along the easternmost states and coastal areas, and this appears to be separate from the central flyway via a large gap where there are no roost sightings.
It should be made clear that in this study we are assuming the locations of roosts indicate the routes of monarch migration; however, this assumption is not unrealistic. At the very least, we argue these roosts show where the majority of monarchs migrate, and where their numbers are great enough so that large overnight clusters form that people can easily observe. The opposite may be true in areas where there are few roost sightings, such as the southeastern United States (Fig. 2). In this area there are clearly considerable numbers of migrating monarch butterflies each fall. In fact, one of the authors of this study lives in northern Georgia and routinely witnesses large waves (i.e. many hundreds) of migrating monarchs each fall in late October (Davis pers. obs.). Furthermore, a volunteer-run tagging program in the Florida panhandle (St. Marks, FL) claimed in 2002 to have captured and tagged over 12,500 migrating monarchs over 10 seasons (R. Rubino pers. comm.). In an earlier report from this same location, Urquhart and Urquhart (1976) witnessed ‘countless thousands of individuals’ annually from 1970 to 1975. So there is considerable evidence that monarch butterflies migrate to some extent throughout the southeastern states, but the few roost sightings in this area suggest that their numbers are small enough so that any overnight clusters may go unnoticed by the general public. Alternatively, it could be argued that monarchs migrate in equal numbers throughout their range in eastern North America, but that for some reason, they only form roosts in the interior states (i.e. the central flyway). We consider this idea unlikely, and support the former.
There is reason to believe that the eastern flyway described here is not an optimal route for migration to Mexico. First, tag recovery data from the overwintering colonies in Mexico support this idea: monarchs tagged in the fall at east coast sites have a very low chance of being recovered in Mexico, considerably lower than those tagged at inland locations (Garland and Davis 2002; Brindza et al. in review). Moreover, of the 12,500 monarchs tagged in the Florida panhandle, only 3 of these had been recovered in the Mexican overwintering sites (R. Rubino pers. comm.). Secondly, when we compared the latitudinal progression of roost formation between the central and eastern flyways we found that roost formation (which we assume is indicative of migration progress) in the eastern flyway lagged behind the central flyway roosts (Fig. 3). There were also very few roosts sighted in coastal states below the mid-Atlantic, despite the fact that there are many Journey North participants at these coastal areas (Fig. 2, inset), suggesting that this flyway does not progress further south than the mid-Atlantic. Taken together, this evidence suggests the eastern flyway is not optimal for monarch migration to Mexico. The reasons for this could be several. It may be that for many eastern flyway migrants, the Mexican overwintering site is not their final destination. Indeed, monarchs migrating southward along the Atlantic coast could continue following the coast of Florida until they reach the southern end, where they could join with the continuously breeding population that is known to occur there (Altizer et al. 2000). In addition, monarchs have also been shown to end up in Cuba, perhaps via this same scenario (Dockx et al. 2004), though it is not clear if this is by design or accident. Alternatively, it may be that the Mexico sites are indeed the intended destination, but the coastal journey is made more perilous by numerous water crossings and/or the threat of being blown offshore.
Our analysis of the eastern flyway is somewhat consistent with early speculations from tag-recapture trajectories. Urquhart and Urquhart (1979b) showed that while the majority of fall monarchs tagged in southern Ontario were recaptured in locations to the south or southwest, some monarchs were later found in coastal states southeast of their release point. Based on this observation, they speculated this was an ‘aberrant’ migration route (Urquhart and Urquhart 1979a), and provided some evidence these individuals follow the eastern coastline southward until they end up in southern Florida or Cuba. More recent studies using stable isotope analyses have confirmed that indeed some monarchs from North America end up in Cuba (Dockx et al. 2004). Our results are consistent with the Urquharts’ premise, although we note that their use of the term ‘aberrant’ to describe this flyway is not necessarily appropriate, given that this flyway is used year after year. A better term is perhaps ‘suboptimal’, especially given that at least some monarchs tagged at these locations do make it to the Mexican wintering colonies (Garland and Davis 2002; Brindza et al. in review). Also, even if this is a suboptimal flyway, since large numbers of monarchs regularly occur in these coastal areas and can become extremely concentrated, especially at geographic peninsulas (Garland and Davis 2002), their presence does allow annual estimates of abundance to be obtained (Walton et al. 2005; Gibbs et al. 2006), which makes these locations useful to monarch conservation efforts.
The migration through Texas deserves comment because of the consistency we found with prior work. In our map of the flyways we noticed that the migration through Texas appears to be divided into two regions; a main route that traverses the center of the state, and a second route that follows the eastern coastline (Fig. 2). These same spatial patterns have been documented before from a state-wide citizen-science program, Texas Monarch Watch (Calvert and Wagner 1997), whereby reports of monarch ‘accumulations’ (i.e. sightings of 100 monarchs or more, but not necessarily of roosts) in the state are compiled with a phone hotline. Calvert and Wagner (1997) used this data from 1993 to 1995 to describe these two Texas flyways in which they called the ‘central’ and ‘coastal’ flyways. The similarity between those results and ours a decade later speaks to the credibility of both data sources plus the year-to-year consistency of this state-wide pattern.
A final important observation that is not apparent from the flyway map in Fig. 2 is the ‘ephemeral’ nature of the monarch roosts, because the 3 years of data considered here (2005–2007) were combined to spatially demonstrate the flyways. Importantly, the majority of roost sightings occurred only once in that time. That is, when the sightings were categorized by town (i.e. roost location), 85% of towns only appeared in the data set in 1 year. In 10.7% of cases, a town was represented in two of the 3 years, and only 4.3% of towns appeared in all three years. In other words, monarchs do not appear to consistently roost in the same locations within the flyways each year. This is also emphasized by the text of the reports sent in by many observers, such as “in our 30 years living in this neighborhood we have never seen this before,” or “they were here one year but haven’t come in the last 5 years.” This ephemeral nature of roosts has large implications for migration habitat conservation. Importantly, it suggests that roost site selection is somewhat random, and if so, it would be difficult from a management standpoint to determine how best to protect roost sites. Therefore, it might make more sense to focus conservation efforts on preserving other migration resources such as nectar plants in the flyway, which monarchs use during migration to feed from and build up their lipid reserves (Brower et al. 2006). The ephemeral nature of monarch roosts also emphasizes the value of taking a large-scale perspective on conserving migration resources, as well as in the scientific study of monarch migration.
With the continued existence of this citizen-science program, we will eventually be able to fill in gaps where there are few Journey North participants, such as areas of southern Georgia and Alabama, or much of West Virginia, for example (Fig. 2 inset). In addition, these data will eventually allow for further scientific questions regarding the resources monarchs need for their migration. For example, future projects could compile information regarding the characteristics of roosts (i.e. what tree types are used, what is the duration of roosts, is there nectar available), as well as the behavior of monarchs at the roosts especially in the early morning (i.e. do they feed before flying off?). In any case, the continued existence of this program will serve as not only an excellent educational resource, but also as a scientific tool for studying monarch butterfly migratory biology and will provide conservationists with information they need to protect this aspect of its life cycle in North America.
References
Altizer SM, Oberhauser K, Brower LP (2000) Associations between host migration and the prevalence of a protozoan parasite in natural populations of adult monarch butterflies. Ecol Entomol 25:125–139. doi:10.1046/j.1365-2311.2000.00246.x
Brindza L, Brower LP, Davis AK, Van Hook T (in review) Comparative success of monarch butterfly migration to overwintering sites in Mexico from inland and coastal sites in Virginia. J Lepid Soc
Brower LP (1995) Understanding and misunderstanding the migration of the monarch butterfly (Nymphalidae) in North America: 1857–1995. J Lepid Soc 49:304–385
Brower LP (1996) Monarch butterfly orientation: missing pieces of a magnificent puzzle. J Exp Biol 199:93–103 Medline
Brower LP, Fink LS, Walford P (2006) Fueling the fall migration of the monarch butterfly. Integr Comp Biol 46:1123–1142. doi:10.1093/icb/icl029
Calvert WH, Wagner M (1997) Patterns in the monarch butterfly migration through Texas—1993 to 1995. In: Hoth J, Merino L, Oberhauser K, Pisanty I, Price S, Wilkinson T (eds) 1997 North American conference on the monarch butterfly Comission for Environmental Cooperation. Montreal, Quebec, pp 119–125
Davis AK, Garland MS (2004) Stopover ecology of monarchs in coastal Virginia: using ornithological methods to study monarch migration. In: Oberhauser K, Solensky M (eds) The monarch butterfly. Biology and conservation. Cornell University Press, Ithaca, pp 89–96
Davis AK, Howard E (2005) Spring recolonization rate of monarch butterflies in eastern North America: new estimates from citizen science data. J Lepid Soc 59:1–5
Dockx C, Brower LP, Wassenaar LI, Hobson KA (2004) Do North American monarch butterflies travel to Cuba? Stable isotope and chemical tracer techniques. Ecol Appl 14:1106–1114. doi:10.1890/03-5128
Garland MS, Davis AK (2002) An examination of monarch butterfly (Danaus plexippus) autumn migration in coastal Virginia. Am Midl Nat 147:170–174. doi:10.1674/0003-0031(2002)147[0170:AEOMBD]2.0.CO;2
Gibbs D, Walton R, Brower L, Davis AK (2006) Monarch butterfly (Lepidoptera, Nymphalidae) migration monitoring at Chincoteague, VA and Cape May, NJ: a comparison of long-term trends. J Kans Entomol Soc 79:156–164. doi:10.2317/0504.06.1
Howard E, Davis AK (2004) Documenting the spring movements of monarch butterflies with Journey North, a citizen science program. In: Oberhauser K, Solensky M (eds) The monarch butterfly. Biology and conservation. Cornell University Press, Ithaca, pp 105–114
Meitner CJ, Brower LP, Davis AK (2004) Migration patterns and environmental effects on stopover of monarch butterflies (Lepidoptera, Nymphalidae) at Peninsula Point, Michigan. Environ Entomol 33:249–256
Moskowitz D, Moskowitz J, Moskowitz S, Moskowitz H (2001) Notes on a large dragonfly and butterfly migration in New Jersey. North East Nat 8:483–490
Oberhauser K, Peterson AT (2003) Modeling current and future potential wintering distributions of eastern North American monarch butterflies. Proc Natl Acad Sci USA 100:14063–14068. doi:10.1073/pnas.2331584100
Oberhauser K, Rivers ERL (2003) Monarch butterfly (Danaus plexippus) larvae and Bt maize pollen: a review of ecological risk assessment for a non-target species. AgBiotechNet 5:1–7
Oberhauser K, Prysby MD, Mattila HR, Stanley-Horn DE, Sears MK, Dively G et al (2001) Temporal and spatial overlap between monarch larvae and corn pollen. Proc Natl Acad Sci USA 98:11913–11918. doi:10.1073/pnas.211234298
Oberhauser KS, Brinda SJ, Weaver S, Moon RD, Manweiler SA, Read N (2006) Growth and survival of monarch butterflies (Lepidoptera: Danaidae) after exposure to permethrin barrier treatments. Environ Entomol 35:1626–1634. doi:10.1603/0046-225X(2006)35[1626:GASOMB]2.0.CO;2
Stalleicken J, Mukhida M, Labhart T, Wehner R, Frost BJ, Mouritsen H (2005) Do monarch butterflies use polarized skylight for migratory orientation? J Exp Biol 208:2399–2408. doi:10.1242/jeb.01613
Statistica (2003) Statistica version 6.1, Statsoft Inc.
Urquhart FA (1976) Found at last: the monarch’s winter home. Natl Geogr Mag 150:161–173
Urquhart FA, Urquhart NR (1976) Migration of butterflies along the gulf coast of Northern Florida. J Lepid Soc 30:59–61
Urquhart FA, Urquhart NR (1977) Overwintering areas and migratory routes of the monarch butterfly (Danaus p. plexippus, Lepidoptera: Danaidae) in North America, with special reference to the western population. Can Entomol 109:1583–1589
Urquhart FA, Urquhart NR (1978) Autumnal migration routes of the eastern population of the monarch butterfly (Danaus plexippus) (L.) (Danaidae:Lepidoptera) in North America to the overwintering site in the neovolcanic plateau of Mexico. Can J Zool 56:1754–1764
Urquhart FA, Urquhart NR (1979a) Aberrant autumnal migration of the eastern population of the monarch butterfly, Danaus p. plexippus (Lepidoptera: Danaidae) as it relates to the occurrence of strong westerly winds. Can Entomol 111:1281–1286
Urquhart FA, Urquhart NR (1979b) Breeding areas and overnight roosting locations in the northern range of the monarch butterfly (Danaus plexippus plexippus) with a summary of associated migratory routes. Can Field Nat 93:41–47
Walton RK, Brower LP, Davis AK (2005) Long-term monitoring and fall migration patterns of the monarch butterfly (Nymphalidae: Danainae) in Cape May, NJ. Ann Entomol Soc Am 98:682–689. doi:10.1603/0013-8746(2005)098[0682:LMAFMP]2.0.CO;2
Acknowledgements
We thank the thousands of volunteers who participate in the Journey North program, without whose help, this study would have never been possible. We thank Mike Quinn, Chip Taylor and Don Davis for sharing data collected through the Texas Monarch Watch and Monarch Watch networks, and Cindy Schmid for tirelessly managing the Journey North databases. Lincoln Brower served as scientific advisor to the Journey North program. Support for Journey North is provided by the Annenberg Foundation. We also thank Sean Sterrett for helpful comments on an early stage of the manuscript. AKD was supported by funds from the D.B. Warnell School of Forestry and Natural Resources.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Howard, E., Davis, A.K. The fall migration flyways of monarch butterflies in eastern North America revealed by citizen scientists. J Insect Conserv 13, 279–286 (2009). https://doi.org/10.1007/s10841-008-9169-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-008-9169-y