
Abstract
Aquaculture provides half of the fish for human consumption, and the trend is on a significant rise over the coming decades. However, the soaring price of traditional ingredients used in aquafeeds is becoming prohibitive, especially in the case of capture fishery derivatives. Therefore, new alternative ingredients and additives are required in order to substitute fish meal and fish oil in aquaculture feeds. Microalgae are a reliable alternative to these as they are potential stabilizing agents against nutrient oxidation. In the present study, three experimental aquafeeds were elaborated with 15% microalgae biomass (Isochrysis galbana, Nannochloropsis gaditana, and Scenedesmus almeriensis); these were then stored under different temperature and light conditions for 15 months in order to analyze the stability of proteins, lipids, fatty acids, and carotenoids. The antioxidant activity of the natural pigments present in microalgae allowed frozen microalgae-based aquafeeds to maintain stable quality over 9 months of storage. Nannochloropsis- and Isochrysis-supplemented feeds had higher eicosapentaenoic and docosahexaenoic acid contents than the microalgae-free control feed. However, longer storage times led to a drop in protein and carotenoid levels.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Historically, fish meal has been used as the main ingredient in aquafeeds due to its balanced nutrient content, palatability, and high digestibility (Cowey 1994; New 2001). Nutritionally, fish meal is an appropriate protein source owing to its high protein content (usually over 60%) with digestibility coefficients above 80%. It also possesses a high biological value due to its well-balanced essential amino acid composition, which is similar to fish muscle. Additionally, elevated contents of highly unsaturated fatty acids (HUFAs), mainly docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA), along with important minerals (Ca, P, and Mg), vitamins (A, B12, C, D3, choline, and inositol), as well as trace minerals, are reported in fish meal (de la Higuera and Cardenete 1993). Nevertheless, its quality is quite variable and source dependent (fish species, season, and manufacturing process). Fish meal derives both from industrial fishery catches and from fish by-products obtained from seafood processing for human consumption.
The current supply of fish meal and fish oil could be outstripped by current demand as a result of unsupportable levels of industrial aquaculture and the diversification of farmed species. Likewise, the availability of protein ingredients is impacted by increasing demand and is leading to continually rising prices (FAO 2014).
Economic problems aside, a communication from the Commission to the European Parliament was adopted in 2009 regarding strategic guidelines for the sustainable development of EU aquaculture, the main objectives being to ensure public health and safe fishing products, as well as to set the aquaculture production system on a sustainable course. For this reason, an intensive search for alternative protein and oil sources, which totally or partially replace fish meal and oil in aquafeeds, is being carried out (Silva et al. 2010); this is in step with the FAO recommendations requiring a reduction in the use of fish meal and oil from extractive fishing.
Currently, interest is growing in microalgae as an alternative source of protein and lipids, vitamins, minerals, and nutraceutical substances in aquafeeds (Müller-Feuga 2000; Pereira et al. 2012; Güroy et al. 2011; Vizcaíno et al. 2014, 2016). In the last few decades, the popularity of microalgae has increased due to their appropriate size, high nutritional value, rapid growth, and antioxidant capacity (Roy and Pal 2015).
In addition, microalgae possess an interesting fatty acid profile due to their long-chain, polyunsaturated fatty acid (PUFA) content, which is so essential in fish diets; they are also a valuable source of essential vitamins (i.e., B6, B12, folic acid) and natural pigments (i.e., β-carotene, astaxanthin, diterpene) (Spolaore et al. 2006). Authors such as Tibaldi et al. (2015), Walker and Berlinsky (2011), and Vizcaíno et al. (2014) have performed different studies using Isochrysis sp., Nannochloropsis sp., and Scenedesmus almeriensis as dietary ingredients for feeding aquaculture fish species. These studies concluded that microalgal biomass enhances growth, nutrient utilization, and gastrointestinal morphology in different marine fish species (Vizcaíno et al. 2014, 2016). Sarker et al. (2016) found improved results in Nile tilapia growth with greater weight gain and a better protein efficiency ratio when fully replacing fish oil with Schizochytrium sp. microalgal biomass.
Previous studies have also shown that microalgae are a promising alternative source of fish meal and fish oil due to their biochemical composition, demonstrating that a small amount of microalgae in aquafeed (5% dry weight (d.w.)) leads to a slight improvement in survival rates compared to the commercial control diet; this is related to a better immune response as well as a significantly higher antioxidant content in prawn muscle (Li et al. 2015). Other studies have evaluated the stability of several fish and microalgal oils to oxidation, finding that oils obtained from microalgae can be as stable as krill oil and significantly more stable than other fish oils (Ryckebosch et al. 2013). Nonetheless, the oxidative stability of feed must be evaluated since there is evidence that using oxidized fish oil in diets has a negative effect on the immune system; there is also evidence that enzyme activity is increased because of antioxidant activity as is the stress response in fish (Mourente et al. 2002; Alves Martins et al. 2007). Lipidic oxidation is caused by chain reactions in which lipidic radicals react with oxygen-forming hydroperoxide and other lipidic radicals (Hernández et al. 2014); this process can be halted using antioxidants. Traditionally, synthetic antioxidants (butylhydroxytoluene (BHT), butylhydroxyanisole, and ethoxyquin) with high antioxidant activity have been used (Hamre et al. 2010), although other studies have demonstrated the presence of these compounds in fish muscle, which negatively affects health (Lundebye et al. 2010). Consequently, the use of natural antioxidants is being increasingly studied since food additive awareness is growing in the society. Nevertheless, no in-depth study on the effects of natural antioxidants on aquafeed storage has so far been carried out (Hamre et al. 2010).
Taking the above points into consideration, the aim of the present work was to study the stability of nutrients (proteins and lipids, with special attention to the fatty acid and carotenoid contents), as well as to evaluate the use of microalgae as potential stabilizing agents against nutrient oxidation in three experimental aquafeeds containing 15% Isochrysis galbana (ISO), Nannochloropsis gaditana (NAN), or Scenedesmus almeriensis (SCE) biomass, and then comparing them to a microalgae-free control (CT) feed under different storage conditions for a period of 15 months.
Materials and methods
Experimental diets
Microalgal biomasses were provided by the Estación Experimental “Las Palmerillas” (Fundación Cajamar, Almería, Spain). In brief, the cells were produced in an industrial-sized outdoor vertical tubular photobioreactor (3000 L) in continuous mode at a 0.3 day−1 dilution rate. This facility is designed to operate with seawater or fresh water in a closed circuit, with recirculation of the culture medium. Agricultural fertilizers were used instead of pure chemicals at a calcium nitrate concentration of 11 mM, according to the commercial Algal medium (Bionova, Santiago de Compostela, Spain). The inorganic compounds were added to artificial seawater (30 g L−1 NaCl; TorreSal, Unión Salinera de España). The cultures were grown at pH 8.0 by on-demand injection of CO2, and the temperature was maintained within the 23–25 °C range by passing thermostatically controlled water through a heat exchanger located inside the reactor. The biomass was harvested daily by centrifugation (RINA continuous centrifuge, Riera Nadeu SA, Spain), frozen at − 18 °C, freeze-dried, and finally milled to obtain a homogenized powder (< 100 μm) that was stored in the dark at − 20 °C until preparation of the experimental diets. Four isonitrogenous (40% dry weight) experimental feeds were elaborated at the CEIA3-University of Almería facilities (Experimental Diets Service, http://www.ual.es/stecnicos_spe). Three experimental feeds were formulated to contain 15% freeze-dried Isochrysis galbana (ISO), Nannochloropsis gaditana (NAN), or Scenedesmus almeriensis (SCE) biomass. A microalgae-free feed was used as the CT. The ingredient composition of the experimental feeds is shown in Table 1. Experimental aquafeeds were formulated to contain 35–40% crude protein and 10% crude lipids. Microalgal biomasses were used as alternative ingredients to reduce the use of fish meal and fish oil. However, microalgal biomasses contain different levels of proteins (from 35 to 44% d.w.), lipids (from 9 to 27% d.w.), and carbohydrates (from 22 to 36% d.w.). For this reason, the same inclusion level does not necessarily result in the same level of fish meal and fish oil in each experimental feed. Consequently, the amount of fish meal, fish oil, and maltodextrin was adjusted individually in each feed to achieve the required level of dietary proteins and lipids according to Vizcaíno et al. (2014). All the experimental aquafeeds were elaborated using a single screw laboratory extruder (Miltenz 51SP, JS Conwell Ltd., New Zealand) set to obtain 2-mm diameter pellets that were dried and kept frozen at − 20 °C until use.
Analytical procedures
The fatty acid profile of the experimental diets was determined by gas chromatography (Agilent Technologies 6890N Series Gas Chromatograph, USA) after direct transesterification, as described by Rodríguez-Ruiz et al. (1998). The chromatographic system was equipped with a capillary column of fused silica Omegawax (0.25 mm ×30 m, 0.25-μm standard film, Supelco, USA) and a flame ionization detector (FID). Nitrogen was used as the carrier gas at a flow rate of 58.1 mL min−1 and a split ratio of 1:40. The injector and detector temperatures were set at 250 and 260 °C, respectively. The oven temperature was initially set at 150 °C for 3 min, then programmed to increase to 240 °C at a rate of 7.5 °C min−1 and finally set at 240 °C for 12 min.
The total carotenoid content was determined by chromatography (HPLC) as described by Cerón et al. (2007). Neoxanthin, lutein, violaxanthin, fucoxanthin, and β-carotene standards were purchased from Sigma (USA). Vaucheriaxanthin, zeaxanthin, diatoxanthin, and dinoxanthin standards were purchased from DHI (Hørsholm, Denmark). Dinoxanthin is not present in I. galbana, but it is used to determine the diadinoxanthin content. Pigment quantification was based on calibration curves taken at different standard concentrations.
The total lipid content of the experimental diets was determined according to Kochert (1978). The protein content in the biomass was analyzed as described by González-Lopez et al. (2010).
All measurements were performed in triplicate.
Experimental diet storage: preservation conditions
The stability of the experimental aquafeeds was studied by analyzing the influence of the storage period, the temperature, and the presence or absence of light on their biochemical composition. The storage conditions in an inert atmosphere were as follows: 25 °C at ambient light, 25 °C under the absence of light, and refrigerated (6 °C) and frozen (− 20 °C) under the absence of light. The analysis of proteins and total lipids (total lipids, fatty acids, and total carotenoids) was carried out periodically over the 15-month period. Two replicates were taken for analyses from each condition assayed.
Statistical analyses
Statistical data analyses were performed using the Statgraphics Centurion XVI software package. Data, in percentage, were arcsin(x 1/2) transformed. The normality and homogeneity analyses were performed using the Kolmogorov–Smirnov and Levene tests, respectively. Multifactor ANOVA tests were used to study the effect of the factors (time, temperature, irradiance, and aquafeed) and their interactions (time-temperature, time-irradiance, time-aquafeed, temperature-aquafeed, irradiance-aquafeed) at a 95% confidence level, for the protein, total lipid, total fatty acid, carotenoid, DHA, EPA, saturated fatty acid (SFA), monounsaturated fatty acid (MUFA), and PUFA contents with the aim of deciding the most influential variable in the biochemical composition.
Results and discussion
The aim of the present work was to study the stability of aquafeeds supplemented with microalgae, for the partial replacement of fish meal and fish oil, evaluating different storage conditions such as time (t), temperature (T), and the presence or absence of light (I), as well as the type of aquafeed (A), on the stability of macronutrients (proteins and lipids, with special attention to the fatty acid profile, mainly EPA and DHA; as well as the effect of carotenoids in aquafeeds, which act as natural antioxidants leading to stabilized nutrients that are more susceptible to oxidation). The experimental aquafeeds were elaborated with 15% microalgal biomass (I. galbana, N. gaditana, or S. almeriensis) and stored under different conditions for a period of 15 months. Thus, a stability study of aquafeeds was designed.
Total protein
The evolution of the total protein content in aquafeeds was monitored over time under the different storage conditions. Figure 1 shows the effect of time (a), temperature (b), and light (c) on the loss of protein content, whereas Table 2 provides a statistical analysis of the abovementioned factors.

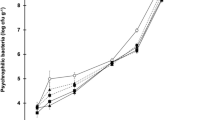
Influence of storage conditions on the protein and total lipid content loss in function of the time (a), temperature (b), and light (c) and on the total carotenoid content loss in function of time (d), temperature (e), and light (f). Values are the mean ± standard deviation in aquafeeds. “Average proteins” (dark blue circles) and “average lipids” (white circles) are average values of every aquafeed for each condition
The initial protein content in aquafeeds was 40% (d.w.); although in the case of S. almeriensis-supplemented feed, this value was significantly lower (35% d.w.) (Fig. 1a). It is known that culture conditions affect protein accumulation in microalgae (Camacho-Rodríguez et al. 2015), and in the case of the S. almeriensis biomass, this may explain why SCE feed had a lower protein content than other microalgae-supplemented feeds. With regard to Table 2, time is the most important factor affecting protein content, but no significant differences were observed whatever the microalgae species used in the aquafeeds, though the effect of interaction time-microalgae species (tA) was the most important. In the case of CT, the protein fraction was stable for 9 months; although after 12 months of storage, this value decreased to 30% (Fig. 1a). However, the NAN and ISO feeds showed a progressive decrease in the total protein content from the beginning (Fig. 1a). According to Gandía-Herrero et al. (2010), maltodextrin promotes the stability of biomolecules so the lower content of this carbohydrate in regard to CT might contribute to the protein reduction observed in NAN and ISO (Table 1); although in the case of ISO, the protein diminution was lower than that in CT (Fig. 1a). In spite of its lower initial protein content, SCE feed stayed preserved throughout the storage period. This could be explained by the ability of S. almeriensis to accumulate lutein under certain growth conditions (Sánchez et al. 2008), and certain pigments have demonstrated high antioxidant activity, thus being able to avoid biomolecular degradation (Cerón et al. 2007).
Regarding the effect of temperature, this factor makes a significant difference to protein content. At 25 °C, the protein level decreased slightly in all aquafeeds containing microalgae when compared to the values of samples stored at − 20 and 6 °C (Fig. 1b); no differences were observed between refrigerated and frozen samples. This observed temperature effect might be explained by the fact that natural pigments supplied by microalgal biomass, which have an antioxidant capability against biomolecular degradation (Cerón et al. 2007), are degraded when stored at room temperature and in the presence of light, with the consequent loss of pigments from antioxidant activity. Nevertheless, CT did not show protein reduction at 25 °C because the higher maltodextrin content contributes to increased protein stability at this temperature (Ferrari et al. 2013). Likewise, light did not affect the protein content in the aquafeeds (Table 2); although in the case of SCE, a significant drop was observed (Fig. 1c), probably due to the photodegradation of natural antioxidants, as demonstrated by Gouveia and Empis (2003), who found greater carotenoid degradation in microalgal biomass stored under light rather than under dark conditions.
Total lipid content
With reference to the total lipid content, the average values are shown in Fig. 1, observing no lipid degradation during the storage period whatever factor was evaluated (Fig. 1a–c); the value remained stable at 10.2% (d.w.). Similarly, Aryee et al. (2012) studied the effect of temperature (25, 4, − 18, and − 80 °C) over 45 days of storage on fatty acids in salmon oil and reported that the fatty acid composition was not affected by either the temperature or the storage period. Hernández et al. (2014) found no differences in the fat oxidative status of diets stored for 24 weeks; these included the synthetic antioxidant (BHT) and the natural antioxidant rosemary extract, allowing us to establish that both additives protect against lipid oxidation. However, studies carried out by Hamre et al. (2010) concluded that the synthetic antioxidant ethoxyquin can be replaced by natural antioxidants since the inclusion of rosemary extract at the highest level (6.0 g kg−1) achieved lower oxidation values than were reached in the control diet, and with no difference to those that used all the antioxidants present in their experiments.
Total carotenoids
Only experimental aquafeeds supplemented with microalgae included natural carotenoids. NAN was the aquafeed with the highest carotenoid content, 0.24% (d.w.); followed by ISO, 0.19%; and SCE 0.17% (Fig. 1d). The main factors affecting carotenoid stability in aquafeeds are time and temperature, although there is a significant effect from the interaction time and microalgae species (tA) (Table 2). This may be due to the fact that the carotenoid profile is greatly influenced by the microalgal strain as well as the culture conditions.
The carotenoids present in the aquafeeds did not degrade over the first 6 months of storage. After this, however, significant drops in the carotenoid content—35% in ISO, 32% in NAN, and 45% in SCE—were observed up to the end of the storage period (Fig. 1d). These results are in agreement with those of Gouveia and Empis (2003) for dry Chlorella vulgaris and Haematococcus pluvialis biomass, incorporated into formulated feeds for rainbow trout and gilthead seabream. In their study, the authors found that the carotenoid content remained quite stable for a storage period of 2.5 months; although after 6 months, carotenoid content losses of 24 and 18.8% were detected in diets enriched with C. vulgaris and H. pluvialis biomass, respectively.
Regarding temperature, no significant difference was observed between NAN stored at − 20 and 6 °C, with values of 0.23% (d.w.), yet they were significantly lower in feed samples kept at 25 °C (Fig. 1e). In the case of ISO and SCE feeds, the most suitable storage temperature is − 20 °C; indeed, significant drops of 40 and 80% were observed when stored at 6 and 25 °C, respectively (Fig. 1e). Other authors also established that refrigerating at 4 °C allows one to preserve feeds in good condition without adding antioxidants (Hernández et al. 2014). Hamre et al. (2010) achieved the lowest lipidic oxidation values when using natural antioxidants (rosemary extract) compared to those using a mix of synthetic antioxidants.
In the present work, no statistically significant differences were observed between samples stored under light and dark conditions for SCE, although NAN and ISO stored under light conditions suffered significantly higher losses than those under dark conditions (Fig. 1f). Gouveia and Empis (2003) also found no difference between samples stored under light and dark conditions over 2.5 months; although after 1 year, the samples under light conditions suffered a significantly higher carotenoid content loss than those kept under dark conditions.
Fatty acids
HUFAs (mainly EPA and DHA) are essential nutrients in fish nutrition; however, these compounds are quite susceptible to oxidation during aquafeed storage (Alves Martins et al. 2007; Fernandes et al. 2012). In the present study, the effect of the storage conditions on EPA and DHA content losses was quantified, as well as the ω-3/ω-6 ratio.
Fatty acid profile in aquafeeds
Figure 2 shows the effect of time, temperature, and the presence or absence of light, respectively, on the fatty acid profile of aquafeeds, categorized as SFAs, MUFAs, and PUFAs. Concerning the effect of the aquafeed type, Table 2 reveals that this factor had the biggest effect on the fatty acid content (p < 0.05). This is verified in Fig. 2d–g, where the fatty acid content is slightly higher in the CT feed compared to that in the other feeds that included microalgae. It is known that fish oil has a more complete fatty acid profile than microalgal biomass so it is expected that the fatty acid composition of microalgae-supplemented feed is not as ample as that of the CT feed, which includes fish oil as the main lipidic ingredient (data not shown). Previous studies reveal that fish oil substitution by plant oil is possible although an imbalance in the fatty acid profile of the feeds can appear (Montero et al. 2015).

Influence of storage conditions on the fatty acid content loss in function of time (a), temperature (b), and light (c). Effect of time on the fatty acid profile in the CT (d), NAN (e), ISO (f), and SCE (g) feeds
According to Fig. 2, PUFAs are the major fraction of fatty acids in aquafeeds with values up to 4% (d.w.). It reveals that there are no significant differences in the fatty acid profile in terms of the different factors considered during the storage period (time, temperature, and light), showing average values for SFAs and MUFAs ranging from 2.0 to 2.5% (d.w.) and 1.2 to 1.6% (d.w.), respectively.
The ω-3/ω-6 ratio is an important parameter since low values lead to a detrimental effect on fish growth and immunity (Montero et al. 2015). Data shown in Table 3 indicate that the ω-3/ω-6 ratio in ISO and SCE feeds resulted in values of 3.04 and 3.13, respectively, similar to that observed for the CT diet, of 3.04. However, this ratio was 23% lower in the case of the NAN feed due to the lower DHA content. The SCE feed also had a lower EPA content than the other aquafeeds, although in this case, it was compensated for by a higher 18:3ω-3 content. After 15 months of storage, the presence of light seems to be the most affective factor on ω3/ω6 in microalgae-supplemented diets—this ratio decreased by 10% in every case, while in samples stored under dark conditions, no differences were observed. On the other hand, refrigerated and frozen samples had a slightly increased ω-3/ω-6 content. In the CT feed, there was no appreciable drop in this ratio for samples stored under light conditions, probably due to the higher maltodextrin content in this diet which promotes biomolecular stability (Gandía-Herrero et al. 2010). However, previous studies have demonstrated the greater ability of plant natural antioxidants to efficiently avoid lipidic oxidation compared to synthetic antioxidants (Hernández et al. 2014), which have demonstrated a certain carcinogenic capacity (Ito et al. 1985).
EPA and DHA
The main factor affecting the EPA and DHA contents is the type of microalgae used for aquafeed preparation (Table 2). In the case of EPA, the highest initial content was observed in NAN and ISO, with 1 and 0.9% (d.w.), respectively (Fig. 3a). This is due to this HUFA’s high content in the biomass of these microalgae, making up 30 and 28% of total fatty acids, respectively (Camacho-Rodríguez et al. 2014). On this basis, the lowest EPA content was in SCE as this microalga lacks EPA in its fatty acid profile, meaning that fish oil is the sole supply of EPA in this feed.

Influence of storage conditions on EPA (time (a), temperature (b), and light (c)) and DHA (time (d), temperature (e), and light (f)) content loss in aquafeeds. Values are the mean ± standard deviation
With reference to DHA, ISO was expected to be the diet with the highest content since the I. galbana biomass can accumulate up to 1.2% (d.w.) of DHA (Batista et al. 2013). However, as one can observe in Fig. 3d, the CT and SCE diets actually had a higher DHA content than the ISO feed. This could be because less fish oil was added to the ISO feed (Table 1) given that the contribution of I. galbana biomass to the DHA content is significantly lower (1.8 gDHA kg−1 feed) than that of fish oil (4.8 gDHA kg−1). Miller, Nichols, and Carter (2007) remarked on the importance of this HUFA in the replacement of fish oil with DHA-enriched oil from the microalgae Schizochytrium sp.—they observed no decrease in the HUFA content of the feeds.
After 6 months of storage, a slight EPA content loss (lower than 10%) was observed in all of the aquafeeds (Fig. 3a), demonstrating significant differences over time (Table 2). However, this EPA loss did not increase along with increased storage time, probably due to the presence of natural antioxidants in the aquafeeds (natural-like pigments in the microalgae-supplemented feeds and vitamins in the control feed). It has been reported that biomolecules that have an antioxidant capacity avoid PUFA degradation (Cerón et al. 2007). Other authors reported similar results, such as Hamre et al. (2010), who studied lipidic oxidation in fish feeds by adding different antioxidant agents, concluding that natural antioxidants achieved the lowest oxidation values when compared to synthetic antioxidant-supplemented feeds, as well as the positive effects in terms of food safety in a society with a growing awareness of food additives. In the case of DHA, no significant effect was appreciated in the aquafeeds during the storage period tested (Fig. 3d) in spite of the fact that Table 2 reveals a significant effect from storage time on DHA content, probably due to measurement fluctuation caused by experimental deviations. As can be observed in Fig. 3b, e, the EPA and DHA contents in aquafeeds were not influenced by the storage temperature (Table 2).
The presence or absence of light is the second main factor affecting both EPA and DHA content loss (Table 2). In the case of CT, no difference was observed in samples stored under different light conditions (Fig. 3c, f). However, microalgae-supplemented aquafeeds showed a significant drop in both the EPA and DHA contents when stored under light (Fig. 3c, f). Antioxidant activity and the behavior of compounds could explain the effect of light on the EPA and DHA content of aquafeeds. With regard to the innocuous effect of light on the CT feed, this might be explained by the presence of maltodextrin, which, as explained before, contributes to biomolecular stability (Gandía-Herrero et al. 2010). Moreover, Cerón et al. (2007) demonstrated that the antioxidant activity of pigment esters is significantly higher than those offered by free pigments, and this could explain why these PUFAs are able to form esters with pigments. Under dark conditions, when higher pigment content remains in the aquafeeds, these esters provide a greater antioxidant effect with lower PUFA content loss (Table 4).
Conclusion
To conclude, microalgae are potential ingredients for aquafeeds due to their stabilizing activity against nutrient oxidation. Overall, the nutrient composition was not affected when 15% of I. galbana, N. gaditana, or S. almeriensis biomass was used as a fish meal and fish oil replacement. The nutrient stability in microalgae-supplemented aquafeeds after 9 months of storage was similar to that found in a control feed elaborated mainly with capture fishery derivatives. A significant protein loss was observed, but this was lower in aquafeeds supplemented with Isochrysis or Scenedesmus biomass than that in the microalgae-free control feed. The carotenoid content followed the same trend, with no difference observed in aquafeeds stored at − 20 and 6 °C, although samples kept at 25 °C showed a significantly higher loss. In addition, samples stored under dark conditions were better preserved than those kept under light conditions, probably due to the better preservation of feeds that contained natural antioxidants. The total lipid content and fatty acid profile in aquafeeds were not affected by the storage conditions, ensuring the stability of these nutrients for at least 15 months. Admittedly, the PUFA content was slightly higher in the control feed than in the microalgae-supplemented feeds although the EPA content was higher in NAN and ISO than in the control feed, which shows that the microalgal biomass composition also affected the quality of the aquafeeds.
References
Alves Martins D, Alfonso LO, Hosoya S, Lewis-McCrea LM, Valente LM, Lall SP (2007) Effects of moderately oxidized dietary lipid and the role of vitamin E on the stress response in Atlantic halibut (Hippoglossus hippoglossus L.) Aquaculture 272:573–580
Aryee ANA, Simpson BK, Phillip LE, Cue RI (2012) Effect of temperature and time on the stability of salmon skin oil during storage. J Am Oil Chem Soc 89:287–292
Batista AP, Gouveia L, Bandarra NM, Franco JM, Raymundo A (2013) Comparison of microalgal biomass profiles as novel functional ingredient for food products. Algal Res 2:164–173
Camacho-Rodríguez J, González-Céspedes AM, Cerón-García MC, Fernández-Sevilla JM, Acién-Fernández FG, Molina-Grima E (2014) A quantitative study of eicosapentaenoic acid (EPA) production by Nannochloropsis gaditana for aquaculture as a function of dilution rate, temperature and average irradiance. Appl Microbiol Biotechnol 98:2429–2440
Camacho-Rodríguez J, Cerón-García MC, Fernández-Sevilla JM, Molina-Grima E (2015) The influence of culture conditions on biomass and high value product generation by Nannochloropsis gaditana in aquaculture. Algal Res 11:63–73
Cerón MC, García-Malea MC, Rivas J, Acién FG, Fernández JM, Del Río E, Guerrero MG, Molina E (2007) Antioxidant activity of Haematococcus pluvialis cells grown in continuous culture as a function of their carotenoid and fatty acid content. Appl Microbiol Biotechnol 74:1112–1119
Cowey CB (1994) Review: Amino acid requirements of fish: a critical appraisal of present values. Aquaculture 124:1–11
De la Higuera M, Cardenete G (1993) La proteína en la nutrición de los peces. In: Castello Orvay F (ed) Acuicultura marina: fundamentos biológicos y tecnología de la producción. Universidad de Barcelona, Barcelona, pp 195–226
FAO (C) (2014) The state of world fisheries and aquaculture. http://fao.org/2/sofia14e
Fernandes TJR, Alves RC, Souza T, Silva JMG, Castro-Cunha M, Valente LMP (2012) Lipid content and fatty acid profile of Senegalese sole (Solea senegalensis Kaup, 1858) juveniles as affected by feed containing different amounts of plant protein sources. Food Chem 134:1337–1342
Ferrari CC, Marconi Germer SP, Alvim ID, de Aguirre J (2013) Storage stability of spray-dried blackberry powder produced with maltodextrin or gum arabic. Drying Technol 31:470–478
Gandía-Herrero F, Jiménez-Atiénzar M, Cabanes J, García-Carmona F, Escribano J (2010) Stabilization of the bioactive pigments of Opuntia fruits through maltodextrin encapsulation. J Agric Food Chem 58:10646–10652
González-López CV, Cerón MC, Acién FG, Segovia C, Chisti Y, Fernández JM (2010) Protein measurements of microalgal and cyanobacterial biomass. Bioresour Technol 101:7587–7591
Gouveia L, Empis J (2003) Relative stabilities of microalgal carotenoids in microalgal extracts, biomass and fish feed: effect of storage conditions. Innovative Food Sci Emerg Technol 4:227–233
Güroy D, Güroy B, Merrifield DL, Ergün S, Tekinay AA, Yiğit M (2011) Effect of dietary Ulva and Spirulina on weight loss and body composition of rainbow trout, Oncorhynchus mykiss (Walbaum), during a starvation period. J Anim Physiol Anim Nutr 95:320–327
Hamre K, Kolås K, Sandnes K (2010) Protection of fish feed, made directly from marine raw materials, with natural antioxidants. Food Chem 119:270–278
Hernández A, García García B, Jordán MJ, Hernández MD (2014) Natural antioxidants in extruded fish feed: protection at different storage temperatures. Anim Feed Sci Technol 195:112–119
Ito N, Fukushima S, Tsuda H (1985) Carcinogenicity and modification of the carcinogenic response by BHA, BHT, and other antioxidants. Crit Rev Toxicol 15:109–150
Kochert G (1978) Quantitation of the macromolecular components of microalgae. In: Hellebust J, Craigie JS (eds) Handbook of phycological methods, physiological and biochemical methods. Cambridge University Press, Cambridge, pp 189–195
Li Y, Xiao G, Mangott A, Kent M, Pirozzi I (2015) Nutrient efficacy of microalgae as aquafeed additives for the adult black tiger prawn, Penaeus monodon. Aquac Res 47:3625–3635
Lundebye AK, Hove H, Måge A, Bohne VJB, Hamre K (2010) Levels of synthetic antioxidants (ethoxyquin, butylated hydroxytolueneand butylatedhydroxianisole) in fish feed and commercially farmed fish. Food Addit Contam 27:1652–1657
Miller MR, Nichols PD, Carter CG (2007) Replacement of fish oil with thraustochytrid Schizochytrium sp. L oil in Atlantic salmon parr (Salmo salar L) diets. Comp Biochem Physiol A 148:382–392
Montero D, Benitez-Dorta V, Caballero MJ, Ponce M, Torrecillas S, Izquierdo M, Zamorano MJ, Manchado M (2015) Dietary vegetable oils: effects on the expression of immune-related genes in Senegalese sole (Solea senegalensis) intestine. Fish Shellfish Immunol 44:100–108
Mourente G, Díaz-Salvago E, Bell JG, Tocher DR (2002) Increased activities of hepatic antioxidant defence enzymes in juvenile gilthead sea bream (Sparus aurata L.) fed dietary oxidised oil: attenuation by dietary vitamin E. Aquaculture 214:343–361
Müller-Feuga A (2000) The role of microalgae in aquaculture: situation and trends. J Appl Phycol 12:527–534
New MB (2001) The European aquafeed industry, part II. Fish Farmer 24(5):46–48
Pereira TG, Oliva-Teles A (2003) Evaluation of corn gluten meal as a protein source in diets for gilthead sea bream (Sparus aurata L.) juveniles. Aquacult Res 34:1111-1117
Pereira R, Valente LMP, Sousa-Pinto I, Rema P (2012) Apparent nutrient digestibility of seaweeds by rainbow trout (Oncorhynchus mykiss) and Nile tilapia (Oreochromis niloticus). Algal Res 1:77–82
Rodríguez-Ruíz J, Belarbi E, Sánchez JLG, Alonso DL (1998) Rapid simultaneous lipid extraction and transesterification for fatty acid analyses. Biotechnol Tech 12:689–691
Roy SS, Pal S (2015) Microalgae in aquaculture: a review with special emphasis to nutritional value and fish dietetics. Proc Zool Soc 68:1–8
Ryckebosch E, Bruneel C, Lemahieu C, Muylaert K, Van Durme J, Goiris K, Foubert I (2013) Stability of omega-3 LC-PUFA-rich protoautotrophic microalgal oils compared to commercially available omega-3 LC-PUFA oils. J Agric Food Chem 61:10145–10155
Sánchez JF, Fernández JM, Acién FG, Rueda A, Pérez-Parra J, Molina E (2008) Influence of culture conditions on the productivity and lutein content of the new strain Scenedesmus almeriensis. Process Biochem 43:398–405
Sarker PK, Kapuscinski AR, Lanois AJ, Livesey ED, Bernhard KP, Coley ML (2016) Towards sustainable aquafeeds: complete substitution of fish oil with marine microalga Schizochytrium sp. improves growth and fatty acid deposition in juvenile Nile tilapia (Oreochromis niloticus). PLoS One 11(6):e0156684
Silva FCP, Nicoli JR, Zambonino-Infante JL, Le Gall M, Kaushik S, Gatesoupe FJ (2010) Influence of partial substitution of dietary fishmeal on the activity of digestive enzymes in the intestinal brush border membrane of gilthead sea bream, Sparus aurata and goldfish, Carassius auratus. Aquaculture 306:233–237
Spolaore P, Joannis-Cassan C, Duran E, Isambert A (2006) Commercial applications of microalgae. J Biosci Bioeng 101:87–96
Tibaldi E, Chini Zittelli G, Parisi G, Bruno M, Giorgi G, Tulli F, Venturini S, Tredici MR, Poli BM (2015) Growth performance and quality traits of European sea bass (D. labrax) fed diets including increasing levels of freeze-dried Isochrysis sp. (T-ISO) biomass as a source of protein and n-3 long chain PUFA in partial substitution of fish derivatives. Aquaculture 440:60–68
Vizcaíno AJ, López G, Sáez MI, Jiménez JA, Barros A, Hidalgo L, Camacho-Rodríguez J, Martínez TF, Cerón-García MC, Alarcón FJ (2014) Effects of the microalga Scenedesmus almeriensis as a fishmeal alternative in diets for gilthead sea bream, Sparus aurata, juveniles. Aquaculture 431:34–43
Vizcaíno AJ, Sáez MI, López G, Arizcun M, Abellán E, Martínez TF, Cerón-García MC, Alarcón FJ (2016) Tetraselmis suecica and Tisochrysis lutea meal as dietary ingredients for gilthead sea bream (Sparus aurata L.) fry. J Appl Phycol 28:2843–2855
Walker AB, Berlinsky DL (2011) Effects of partial replacement of fishmeal protein by microalgae on growth, feed intake, and body composition of Atlantic cod. N Am J Aquac 73:76–83
Acknowledgements
Experimental feeds were made at the Universidad de Almería facilities (Experimental Diets Service; http://www.ual.es/stecnicos_spe).
Funding
This work was supported by grant AGR-5334 and the European Regional Development Fund (ERDF and SABANA (grant no. 727874) from the European Union’s Horizon 2020 Research and Innovation program).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Camacho-Rodríguez, J., Macías-Sánchez, M.D., Cerón-García, M.C. et al. Microalgae as a potential ingredient for partial fish meal replacement in aquafeeds: nutrient stability under different storage conditions. J Appl Phycol 30, 1049–1059 (2018). https://doi.org/10.1007/s10811-017-1281-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-017-1281-5